

Dynamic Changes in the Thylakoid Proteome of Cyanobacteria during Light-Regulated Thylakoid Membrane Development
 , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
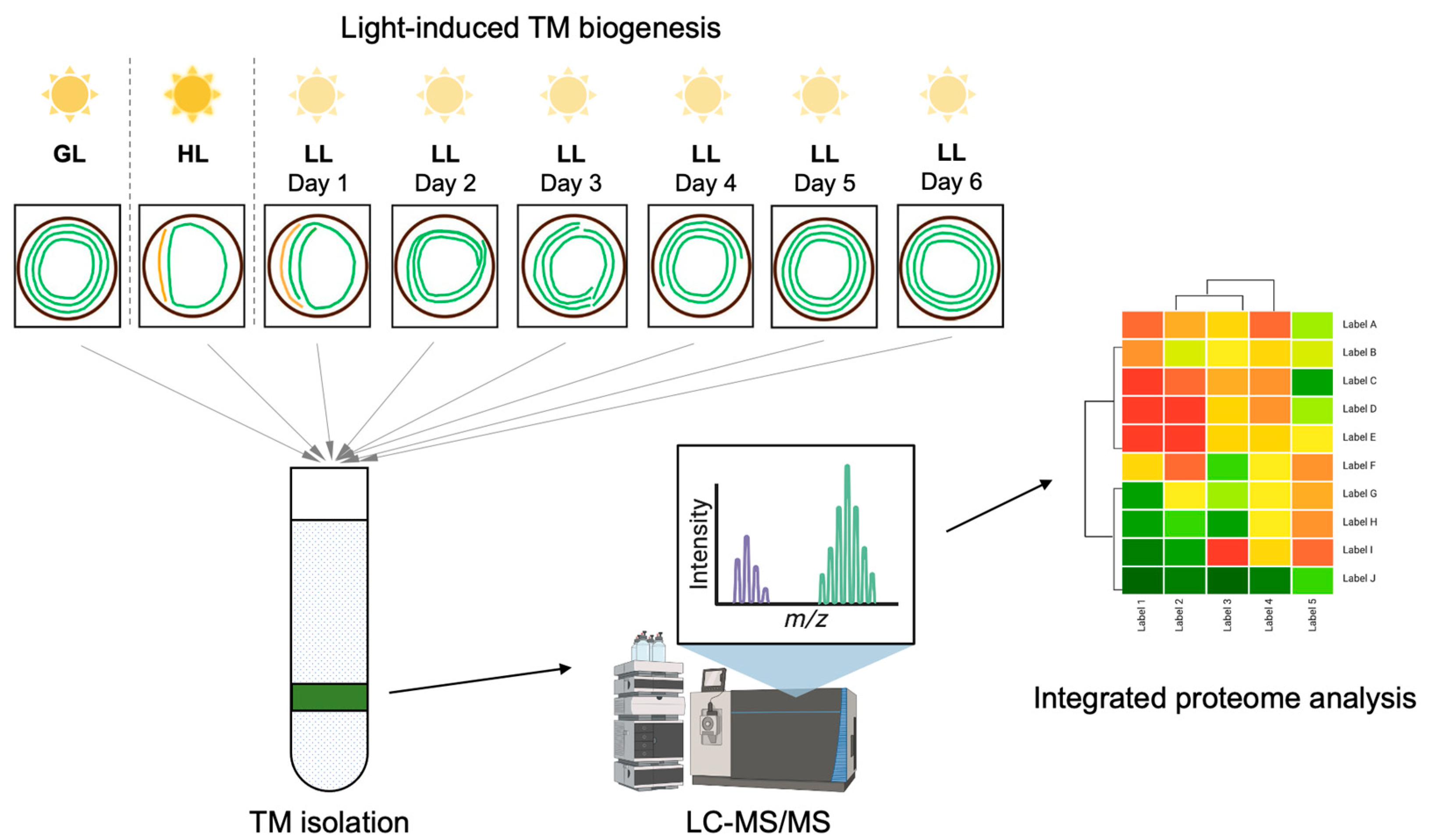
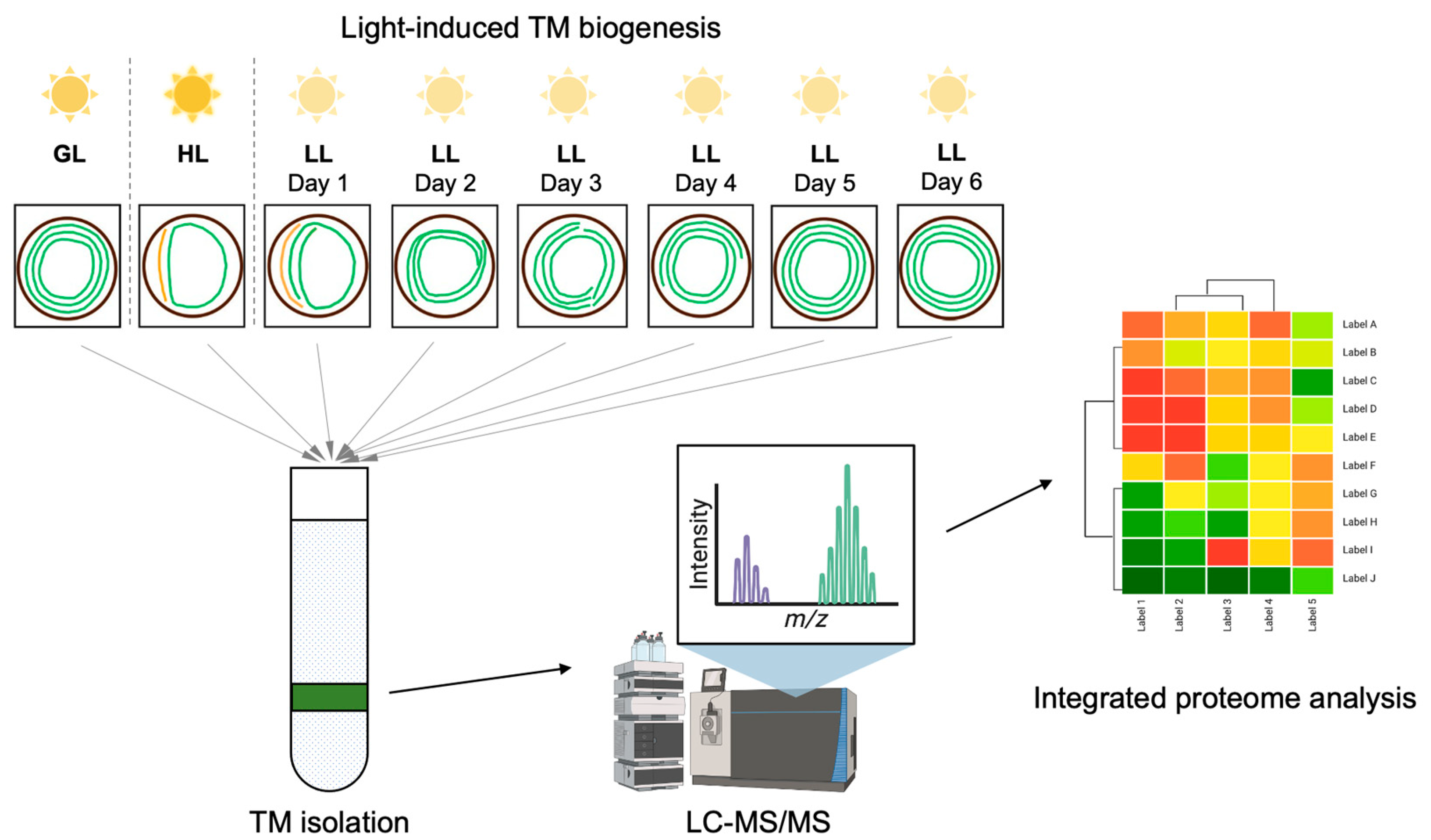
2.1. Proteome Data Generation and Processing
2.2. Statistical Analysis, Pathway and GO Term Enrichment Analysis
2.3. Characterization of Identified Proteins
2.4. The Proteome of HL-Adapted Cells
2.4.1. Photosystems and Assembly Factors
2.4.2. Light-Harvesting Antenna
2.4.3. NDH Complexes
2.4.4. Two-Component Systems
2.4.5. Membrane Transporters
2.4.6. Stress-Related Proteins
2.4.7. Genetic Information Processing
2.5. Proteome Dynamics during LL Treatment and TM Biogenesis
2.5.1. Cluster 1
2.5.2. Cluster 2
2.5.3. Cluster 3
2.5.4. Cluster 4
2.5.5. Cluster 5
2.5.6. Cluster 6
3. Conclusions
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Membrane Preparation
4.3. Mass Spectrometry Measurement
4.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sui, N.; Huang, F.; Liu, L.N. Photosynthesis in Phytoplankton: Insights from the Newly Discovered Biological Inorganic Carbon Pumps. Mol. Plant 2020, 13, 949–951. [Google Scholar] [CrossRef]
- Liu, L.N. Advances in the bacterial organelles for CO2 fixation. Trends Microbiol. 2022, 30, 567–580. [Google Scholar] [CrossRef]
- Pennington, J.T.; Mahoney, K.L.; Kuwahara, V.S.; Kolber, D.D.; Calienes, R.; Chavez, F.P. Primary production in the eastern tropical Pacific: A review. Prog. Oceanogr. 2006, 69, 285–317. [Google Scholar] [CrossRef]
- Liu, L.N. Distribution and dynamics of electron transport complexes in cyanobacterial thylakoid membranes. Biochim. Biophys. Acta 2016, 1857, 256–265. [Google Scholar] [CrossRef]
- Liberton, M.; Saha, R.; Jacobs, J.M.; Nguyen, A.Y.; Gritsenko, M.A.; Smith, R.D.; Koppenaal, D.W.; Pakrasi, H.B. Global Proteomic Analysis Reveals an Exclusive Role of Thylakoid Membranes in Bioenergetics of a Model Cyanobacterium. Mol. Cell Proteom. 2016, 15, 2021–2032. [Google Scholar] [CrossRef] [PubMed]
- Mullineaux, C.W.; Liu, L.N. Membrane dynamics in phototrophic bacteria. Annu. Rev. Microbiol. 2020, 74, 633–654. [Google Scholar] [CrossRef] [PubMed]
- DeRuyter, Y.S.; Fromme, P. Molecular structure of the photosynthetic apparatus. In The Cyanobacteria: Molecular Biology, Genomics, and Evolution; Herrero, A., Flores, E., Eds.; Caister Academic Press: Norfolk, UK, 2008; pp. 217–270. [Google Scholar]
- Vermaas, W.F. Photosynthesis and respiration in cyanobacteria. In Encyclopedia of Life Sciences; Nature Publishing Group: London, UK, 2001; pp. 245–251. [Google Scholar]
- Flannery, S.E.; Hepworth, C.; Wood, W.H.J.; Pastorelli, F.; Hunter, C.N.; Dickman, M.J.; Jackson, P.J.; Johnson, M.P. Developmental acclimation of the thylakoid proteome to light intensity in Arabidopsis. Plant J. 2021, 105, 223–244. [Google Scholar] [CrossRef]
- Huokko, T.; Ni, T.; Dykes, G.F.; Simpson, D.M.; Brownridge, P.; Conradi, F.D.; Beynon, R.J.; Nixon, P.J.; Mullineaux, C.W.; Zhang, P.; et al. Probing the biogenesis pathway and dynamics of thylakoid membranes. Nat. Commun. 2021, 12, 3475. [Google Scholar] [CrossRef]
- Stingaciu, L.R.; O’Neill, H.; Liberton, M.; Urban, V.S.; Pakrasi, H.B.; Ohl, M. Revealing the dynamics of thylakoid membranes in living cyanobacterial cells. Sci. Rep. 2016, 6, 19627. [Google Scholar] [CrossRef]
- Liu, L.N.; Bryan, S.J.; Huang, F.; Yu, J.F.; Nixon, P.J.; Rich, P.R.; Mullineaux, C.W. Control of electron transport routes through redox-regulated redistribution of respiratory complexes. Proc. Natl. Acad. Sci. USA 2012, 109, 11431–11436. [Google Scholar] [CrossRef]
- Zhao, L.S.; Li, C.Y.; Chen, X.L.; Wang, Q.; Zhang, Y.Z.; Liu, L.N. Native architecture and acclimation of photosynthetic membranes in a fast-growing cyanobacterium. Plant Physiol. 2022, 190, 1883–1895. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.S.; Huokko, T.; Wilson, S.; Simpson, D.M.; Wang, Q.; Ruban, A.V.; Mullineaux, C.W.; Zhang, Y.Z.; Liu, L.N. Structural variability, coordination, and adaptation of a native photosynthetic machinery. Nat. Plants 2020, 6, 869–882. [Google Scholar] [CrossRef]
- Casella, S.; Huang, F.; Mason, D.; ZHAO, G.Y.; Johnson, G.N.; Mullineaux, C.W.; Liu, L.N. Dissecting the native architecture and dynamics of cyanobacterial photosynthetic machinery. Mol. Plant 2017, 10, 1434–1448. [Google Scholar] [CrossRef] [PubMed]
- MacGregor-Chatwin, C.; Nurnberg, D.J.; Jackson, P.J.; Vasilev, C.; Hitchcock, A.; Ho, M.Y.; Shen, G.; Gisriel, C.J.; Wood, W.H.J.; Mahbub, M.; et al. Changes in supramolecular organization of cyanobacterial thylakoid membrane complexes in response to far-red light photoacclimation. Sci. Adv. 2022, 8, eabj4437. [Google Scholar] [CrossRef]
- Mullineaux, C.W.; Emlyn-Jones, D. State transitions: An example of acclimation to low-light stress. J. Exp. Bot. 2005, 56, 389–393. [Google Scholar] [CrossRef]
- Bailey, S.; Grossman, A. Photoprotection in cyanobacteria: Regulation of light harvesting. Photochem. Photobiol. 2008, 84, 1410–1420. [Google Scholar] [CrossRef] [PubMed]
- Schuergers, N.; Lenn, T.; Kampmann, R.; Meissner, M.V.; Esteves, T.; Temerinac-Ott, M.; Korvink, J.G.; Lowe, A.R.; Mullineaux, C.W.; Wilde, A. Cyanobacteria use micro-optics to sense light direction. Elife 2016, 5, e12620. [Google Scholar] [CrossRef]
- Chau, R.M.; Bhaya, D.; Huang, K.C. Emergent Phototactic Responses of Cyanobacteria under Complex Light Regimes. mBio 2017, 8, e02330-16. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, L.S.; Liu, L.N. Characterizing the supercomplex association of photosynthetic complexes in cyanobacteria. R. Soc. Open Sci. 2021, 8, 202142. [Google Scholar] [CrossRef]
- Plohnke, N.; Seidel, T.; Kahmann, U.; Rögner, M.; Schneider, D.; Rexroth, S. The proteome and lipidome of Synechocystis sp. PCC 6803 cells grown under light-activated heterotrophic conditions. Mol. Cell. Proteom. 2015, 14, 572–584. [Google Scholar] [CrossRef]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in 1H NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef] [PubMed]
- Hallgren, J.; Tsirigos, K.D.; Pedersen, M.D.; Armenteros, J.J.A.; Marcatili, P.; Nielsen, H.; Krogh, A.; Winther, O. DeepTMHMM predicts alpha and beta transmembrane proteins using deep neural networks. bioRxiv 2022. [Google Scholar] [CrossRef]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Komenda, J. Role of two forms of the D1 protein in the recovery from photoinhibition of photosystem II in the cyanobacterium Synechococcus PCC 7942. Biochim. Biophys. Acta 2000, 1457, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Shukla, P. Tightening the Screws on PsbA in Cyanobacteria. Trends Genet. 2021, 37, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Komenda, J.; Tichý, M.; Eichacker, L.A. The PsbH protein is associated with the inner antenna CP47 and facilitates D1 processing and incorporation into PSII in the cyanobacterium Synechocystis PCC 6803. Plant Cell Physiol. 2005, 46, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Yu, L.; Chitnis, V.P.; Chitnis, P.R. Function and organization of photosystem I in a cyanobacterial mutant strain that lacks PsaF and PsaJ subunits. J. Biol. Chem. 1994, 269, 3205–3211. [Google Scholar] [CrossRef]
- Chitnis, V.P.; Chitnis, P.R. PsaL subunit is required for the formation of photosystem I trimers in the cyanobacterium Synechocystis sp. PCC 6803. FEBS Lett. 1993, 336, 330–334. [Google Scholar] [CrossRef]
- Nickelsen, J.; Rengstl, B. Photosystem II assembly: From cyanobacteria to plants. Annu. Rev. Plant Biol. 2013, 64, 609–635. [Google Scholar] [CrossRef]
- Heinz, S.; Liauw, P.; Nickelsen, J.; Nowaczyk, M. Analysis of photosystem II biogenesis in cyanobacteria. Biochim. Biophys. Acta 2016, 1857, 274–287. [Google Scholar] [CrossRef]
- Hihara, Y.; Ikeuchi, M. Toward the Elucidation of Physiological Significance of pmgA-mediated High-Light Acclimation to Adjust Photosystem Stoichiometry: Effects of the Prolonged High-Light Treatment on pmgA Mutants. In Photosynthesis: Mechanisms and Effects: Volume I–V: Proceedings of the XIth International Congress on Photosynthesis, Budapest, Hungary, 17–22 August 1998; Garab, G., Ed.; Springer: Dordrecht, The Netherlands, 1998; pp. 2929–2932. [Google Scholar]
- Sonoike, K.; Hihara, Y.; Ikeuchi, M. Physiological significance of the regulation of photosystem stoichiometry upon high light acclimation of Synechocystis sp. PCC 6803. Plant Cell Physiol. 2001, 42, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Dann, M.; Ortiz, E.M.; Thomas, M.; Guljamow, A.; Lehmann, M.; Schaefer, H.; Leister, D. Enhancing photosynthesis at high light levels by adaptive laboratory evolution. Nat. Plants 2021, 7, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Rochaix, J.D. Regulation of photosynthetic electron transport. Biochim. Biophys. Acta 2011, 1807, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Edelman, M.; Mattoo, A.K. D1-protein dynamics in photosystem II: The lingering enigma. Photosynth. Res. 2008, 98, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Mulo, P.; Sakurai, I.; Aro, E.M. Strategies for psbA gene expression in cyanobacteria, green algae and higher plants: From transcription to PSII repair. Biochim. Biophys. Acta 2012, 1817, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Komenda, J.; Sobotka, R.; Nixon, P.J. Assembling and maintaining the Photosystem II complex in chloroplasts and cyanobacteria. Curr. Opin. Plant Biol. 2012, 15, 245–251. [Google Scholar] [CrossRef]
- Schelin, J.; Lindmark, F.; Clarke, A.K. The clpP multigene family for the ATP-dependent Clp protease in the cyanobacterium Synechococcus. Microbiology 2002, 148, 2255–2265. [Google Scholar] [CrossRef]
- Clarke, A.K.; Schelin, J.; Porankiewicz, J. Inactivation of the clpP1 gene for the proteolytic subunit of the ATP-dependent Clp protease in the cyanobacterium Synechococcus limits growth and light acclimation. Plant Mol. Biol. 1998, 37, 791–801. [Google Scholar] [CrossRef]
- Xiong, Q.; Feng, J.; Li, S.T.; Zhang, G.Y.; Qiao, Z.X.; Chen, Z.; Wu, Y.; Lin, Y.; Li, T.; Ge, F.; et al. Integrated transcriptomic and proteomic analysis of the global response of Synechococcus to high light stress. Mol. Cell Proteom. 2015, 14, 1038–1053. [Google Scholar] [CrossRef]
- Sui, S.F. Structure of Phycobilisomes. Annu. Rev. Biophys. 2021, 50, 53–72. [Google Scholar] [CrossRef]
- Scheer, H.; Zhao, K.H. Biliprotein maturation: The chromophore attachment. Mol. Microbiol. 2008, 68, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Schluchter, W.M.; Shen, G.; Alvey, R.M.; Biswas, A.; Saunée, N.A.; Williams, S.R.; Mille, C.A.; Bryant, D.A. Phycobiliprotein biosynthesis in cyanobacteria: Structure and function of enzymes involved in post-translational modification. Adv. Exp. Med. Biol. 2010, 675, 211–228. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Ding, W.L.; Zeng, X.L.; Dong, L.L.; Zhao, B.; Zhou, M.; Scheer, H.; Zhao, K.H.; Yang, X. Structure and mechanism of the phycobiliprotein lyase CpcT. J. Biol. Chem. 2014, 289, 26677–26689. [Google Scholar] [CrossRef] [PubMed]
- Peltier, G.; Aro, E.-M.; Shikanai, T. NDH-1 and NDH-2 Plastoquinone Reductases in Oxygenic Photosynthesis. Annu. Rev. Plant Biol. 2016, 67, 55–80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Shuai, J.; Ran, Z.; Zhao, J.; Wu, Z.; Liao, R.; Wu, J.; Ma, W.; Lei, M. Structural insights into NDH-1 mediated cyclic electron transfer. Nat. Commun. 2020, 11, 888. [Google Scholar] [CrossRef]
- Pan, X.; Cao, D.; Xie, F.; Xu, F.; Su, X.; Mi, H.; Zhang, X.; Li, M. Structural basis for electron transport mechanism of complex I-like photosynthetic NAD(P)H dehydrogenase. Nat. Commun. 2020, 11, 610. [Google Scholar] [CrossRef]
- Schuller, J.M.; Birrell, J.A.; Tanaka, H.; Konuma, T.; Wulfhorst, H.; Cox, N.; Schuller, S.K.; Thiemann, J.; Lubitz, W.; Setif, P.; et al. Structural adaptations of photosynthetic complex I enable ferredoxin-dependent electron transfer. Science 2019, 363, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; He, Z.; Xu, M.; Peng, L.; Mi, H. NdhV subunit regulates the activity of type-1 NAD(P)H dehydrogenase under high light conditions in cyanobacterium Synechocystis sp. PCC 6803. Sci. Rep. 2016, 6, 28361. [Google Scholar] [CrossRef]
- Bernát, G.; Appel, J.; Ogawa, T.; Rögner, M. Distinct Roles of Multiple NDH-1 Complexes in the Cyanobacterial Electron Transport Network as Revealed by Kinetic Analysis of P700+ Reduction in Various ndh-Deficient Mutants of Synechocystis sp. Strain PCC6803. J. Bacteriol. 2011, 193, 292–295. [Google Scholar] [CrossRef]
- Howitt, C.A.; Udall, P.K.; Vermaas, W.F. Type 2 NADH dehydrogenases in the cyanobacterium Synechocystis sp. strain PCC 6803 are involved in regulation rather than respiration. J. Bacteriol. 1999, 181, 3994–4003. [Google Scholar] [CrossRef]
- Ashby, M.K.; Houmard, J. Cyanobacterial two-component proteins: Structure, diversity, distribution, and evolution. Microbiol. Mol. Biol. Rev. 2006, 70, 472–509. [Google Scholar] [CrossRef] [PubMed]
- Takai, N.; Nakajima, M.; Oyama, T.; Kito, R.; Sugita, C.; Sugita, M.; Kondo, T.; Iwasaki, H. A KaiC-associating SasA-RpaA two-component regulatory system as a major circadian timing mediator in cyanobacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 12109–12114. [Google Scholar] [CrossRef] [PubMed]
- MacCready, J.S.; Hakim, P.; Young, E.J.; Hu, L.; Liu, J.; Osteryoung, K.W.; Vecchiarelli, A.G.; Ducat, D.C. Protein gradients on the nucleoid position the carbon-fixing organelles of cyanobacteria. Elife 2018, 7, e39723. [Google Scholar] [CrossRef] [PubMed]
- Kanamaru, K.; Kashiwagi, S.; Mizuno, T. A copper-transporting P-type ATPase found in the thylakoid membrane of the cyanobacterium Synechococcus species PCC7942. Mol. Microbiol. 1994, 13, 369–377. [Google Scholar] [CrossRef] [PubMed]
- López-Maury, L.; Giner-Lamia, J.; Florencio, F.J. Redox control of copper homeostasis in cyanobacteria. Plant Signal Behav. 2012, 7, 1712–1714. [Google Scholar] [CrossRef]
- Katoh, H.; Hagino, N.; Grossman, A.R.; Ogawa, T. Genes essential to iron transport in the cyanobacterium Synechocystis sp. strain PCC 6803. J. Bacteriol. 2001, 183, 2779–2784. [Google Scholar] [CrossRef]
- Muramatsu, M.; Hihara, Y. Acclimation to high-light conditions in cyanobacteria: From gene expression to physiological responses. J. Plant Res. 2012, 125, 11–39. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, J.J.; Park, S.I.; Diamond, S.; Boyd, J.S.; Taton, A.; Kim, I.S.; Golden, J.W.; Yoon, H.S. Expression of OsTPX gene improves cellular redox homeostasis and photosynthesis efficiency in Synechococcus elongatus PCC 7942. Front. Plant Sci. 2018, 9, 1848. [Google Scholar] [CrossRef]
- Stork, T.; Laxa, M.; Dietz, M.S.; Dietz, K.J. Functional characterisation of the peroxiredoxin gene family members of Synechococcus elongatus PCC 7942. Arch. Microbiol. 2009, 191, 141–151. [Google Scholar] [CrossRef]
- Stork, T.; Michel, K.P.; Pistorius, E.K.; Dietz, K.J. Bioinformatic analysis of the genomes of the cyanobacteria Synechocystis sp. PCC 6803 and Synechococcus elongatus PCC 7942 for the presence of peroxiredoxins and their transcript regulation under stress. J. Exp. Bot. 2005, 56, 3193–3206. [Google Scholar] [CrossRef]
- Komenda, J.; Sobotka, R. Cyanobacterial high-light-inducible proteins--Protectors of chlorophyll-protein synthesis and assembly. Biochim. Biophys. Acta 2016, 1857, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Rast, A.; Schaffer, M.; Albert, S.; Wan, W.; Pfeffer, S.; Beck, F.; Plitzko, J.M.; Nickelsen, J.; Engel, B.D. Biogenic regions of cyanobacterial thylakoids form contact sites with the plasma membrane. Nat. Plants 2019, 5, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Selão, T.T.; Pisareva, T.; Qian, J.; Sze, S.K.; Carlberg, I.; Norling, B. Deletion of Synechocystis sp. PCC 6803 Leader Peptidase LepB1 Affects Photosynthetic Complexes and Respiration. Mol. Cell. Proteom. 2013, 12, 1192–1203. [Google Scholar] [CrossRef] [PubMed]
- Klinkert, B.; Ossenbühl, F.; Sikorski, M.; Berry, S.; Eichacker, L.; Nickelsen, J. PratA, a Periplasmic Tetratricopeptide Repeat Protein Involved in Biogenesis of Photosystem II in Synechocystis sp. PCC 6803. J. Biol. Chem. 2004, 279, 44639–44644. [Google Scholar] [CrossRef] [PubMed]
- Schottkowski, M.; Gkalympoudis, S.; Tzekova, N.; Stelljes, C.; Schünemann, D.; Ankele, E.; Nickelsen, J. Interaction of the Periplasmic PratA Factor and the PsbA (D1) Protein during Biogenesis of Photosystem II in Synechocystis sp. PCC 6803. J. Biol. Chem. 2009, 284, 1813–1819. [Google Scholar] [CrossRef] [PubMed]
- Zak, E.; Norling, B.; Maitra, R.; Huang, F.; Andersson, B.; Pakrasi, H.B. The initial steps of biogenesis of cyanobacterial photosystems occur in plasma membranes. Proc. Natl. Acad. Sci. USA 2001, 98, 13443–13448. [Google Scholar] [CrossRef] [PubMed]
- Komenda, J.; Knoppova, J.; Kopecna, J.; Sobotka, R.; Halada, P.; Yu, J.; Nickelsen, J.; Boehm, M.; Nixon, P.J. The Psb27 assembly factor binds to the CP43 complex of photosystem II in the cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol. 2012, 158, 476–486. [Google Scholar] [CrossRef]
- Nowaczyk, M.M.; Krause, K.; Mieseler, M.; Sczibilanski, A.; Ikeuchi, M.; Rögner, M. Deletion of psbJ leads to accumulation of Psb27-Psb28 photosystem II complexes in Thermosynechococcus elongatus. Biochim. Biophys. Acta 2012, 1817, 1339–1345. [Google Scholar] [CrossRef]
- Mabbitt, P.D.; Wilbanks, S.M.; Eaton-Rye, J.J. Structure and function of the hydrophilic Photosystem II assembly proteins: Psb27, Psb28 and Ycf48. Plant Physiol. Biochem. 2014, 81, 96–107. [Google Scholar] [CrossRef]
- Zabret, J.; Bohn, S.; Schuller, S.K.; Arnolds, O.; Möller, M.; Meier-Credo, J.; Liauw, P.; Chan, A.; Tajkhorshid, E.; Langer, J.D.; et al. Structural insights into photosystem II assembly. Nat. Plants 2021, 7, 524–538. [Google Scholar] [CrossRef]
- Dühring, U.; Irrgang, K.-D.; Lünser, K.; Kehr, J.; Wilde, A. Analysis of photosynthetic complexes from a cyanobacterial ycf37 mutant. Biochim. Biophys. Acta (BBA)—Bioenerg. 2006, 1757, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Xu, X. Depletion of Vipp1 in Synechocystis sp. PCC 6803 affects photosynthetic activity before the loss of thylakoid membranes. FEMS Microbiol. Lett. 2009, 292, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Bryan, S.J.; Burroughs, N.J.; Shevela, D.; Yu, J.; Rupprecht, E.; Liu, L.N.; Mastroianni, G.; Xue, Q.; Llorente-Garcia, I.; Leake, M.C.; et al. Localisation and interactions of the Vipp1 protein in cyanobacteria. Mol. Microbiol. 2014, 94, 1179–1195. [Google Scholar] [CrossRef] [PubMed]
- Gupta, T.K.; Klumpe, S.; Gries, K.; Heinz, S.; Wietrzynski, W.; Ohnishi, N.; Niemeyer, J.; Spaniol, B.; Schaffer, M.; Rast, A.; et al. Structural basis for VIPP1 oligomerization and maintenance of thylakoid membrane integrity. Cell 2021, 184, 3643–3659 e3623. [Google Scholar] [CrossRef]
- Zhang, S.; Shen, G.; Li, Z.; Golbeck, J.H.; Bryant, D.A. Vipp1 Is Essential for the Biogenesis of Photosystem I but Not Thylakoid Membranes in Synechococcus sp. PCC 7002. J. Biol. Chem. 2014, 289, 15904–15914. [Google Scholar] [CrossRef]
- Göhre, V.; Ossenbühl, F.; Crèvecoeur, M.l.; Eichacker, L.A.; Rochaix, J.-D. One of Two Alb3 Proteins Is Essential for the Assembly of the Photosystems and for Cell Survival in Chlamydomonas. Plant Cell 2006, 18, 1454–1466. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gao, F.; Zhao, J.; Ogawa, T.; Wang, Q.; Ma, W. NdhP is an exclusive subunit of large complex of NADPH dehydrogenase essential to stabilize the complex in Synechocystis sp. strain PCC 6803. J. Biol. Chem. 2014, 289, 18770–18781. [Google Scholar] [CrossRef]
- Wulfhorst, H.; Franken, L.E.; Wessinghage, T.; Boekema, E.J.; Nowaczyk, M.M. The 5 kDa protein NdhP is essential for stable NDH-1L assembly in Thermosynechococcus elongatus. PLoS ONE 2014, 9, e103584. [Google Scholar] [CrossRef]
- Liu, L.N.; Chen, X.L.; Zhang, Y.Z.; Zhou, B.C. Characterization, structure and function of linker polypeptides in phycobilisomes of cyanobacteria and red algae: An overview. Biochim. Biophys. Acta (BBA)—Bioenerg. 2005, 1708, 133–142. [Google Scholar] [CrossRef]
- Li, W.; Su, H.N.; Pu, Y.; Chen, J.; Liu, L.N.; Liu, Q.; Qin, S. Phycobiliproteins: Molecular structure, production, applications, and prospects. Biotechnol. Adv. 2019, 37, 340–353. [Google Scholar] [CrossRef]
- You, X.; Zhang, X.; Cheng, J.; Xiao, Y.; Ma, J.; Sun, S.; Zhang, X.; Wang, H.W.; Sui, S.F. In situ structure of the red algal phycobilisome-PSII-PSI-LHC megacomplex. Nature 2023, 616, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, S.; Geng, X.; Okamoto, S.; Yura, K.; Murata, T.; Go, M.; Ohmori, M.; Ikeuchi, M. Mutational analysis of genes involved in pilus structure, motility and transformation competency in the unicellular motile cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 2001, 42, 63–73. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Vermaas, W. Genetic deletion of proteins resembling Type IV pilins in Synechocystis sp. PCC 6803: Their role in binding or transfer of newly synthesized chlorophyll. Plant Mol. Biol. 1999, 39, 1175–1188. [Google Scholar] [CrossRef] [PubMed]
- Yegorov, Y.; Sendersky, E.; Zilberman, S.; Nagar, E.; Waldman Ben-Asher, H.; Shimoni, E.; Simkovsky, R.; Golden, S.S.; Wang, A.L.; Schwarz, R. A cyanobacterial component required for pilus biogenesis affects the exoproteome. mBio 2021, 12, e03674-20. [Google Scholar] [CrossRef] [PubMed]
- Green, E.R.; Mecsas, J. Bacterial Secretion Systems: An Overview. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Nakai, M.; Sugita, D.; Omata, T.; Endo, T. Sec-Y Protein Is Localized in Both the Cytoplasmic and Thylakoid Membranes in the Cyanobacterium Synechococcus PCC7942. Biochem. Biophys. Res. Commun. 1993, 193, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Knoppová, J.; Sobotka, R.; Tichý, M.; Yu, J.; Konik, P.; Halada, P.; Nixon, P.J.; Komenda, J. Discovery of a Chlorophyll Binding Protein Complex Involved in the Early Steps of Photosystem II Assembly in Synechocystis. Plant Cell 2014, 26, 1200–1212. [Google Scholar] [CrossRef]
- Nixon, P.J.; Barker, M.; Boehm, M.; de Vries, R.; Komenda, J. FtsH-mediated repair of the photosystem II complex in response to light stress. J. Exp. Bot. 2005, 56, 357–363. [Google Scholar] [CrossRef]
- Sacharz, J.; Bryan, S.J.; Yu, J.; Burroughs, N.J.; Spence, E.M.; Nixon, P.J.; Mullineaux, C.W. Sub-cellular location of FtsH proteases in the cyanobacterium Synechocystis sp. PCC 6803 suggests localised PSII repair zones in the thylakoid membranes. Mol. Microbiol. 2015, 96, 448–462. [Google Scholar] [CrossRef]
- Lea-Smith, D.J.; Ross, N.; Zori, M.; Bendall, D.S.; Dennis, J.S.; Scott, S.A.; Smith, A.G.; Howe, C.J. Thylakoid terminal oxidases are essential for the cyanobacterium Synechocystis sp. PCC 6803 to survive rapidly changing light intensities. Plant Physiol. 2013, 162, 484–495. [Google Scholar] [CrossRef]
- Leek, J.T.; Johnson, W.E.; Parker, H.S.; Fertig, E.J.; Jaffe, A.E.; Zhang, Y.; Storey, J.D.; Torres, L.C. sva: Surrogate Variable Analysis; Bioconductor: Boston, MA, USA, 2023. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Sarda-Espinosa, A. dtwclust: Time Series Clustering Along with Optimizations for the Dynamic Time Warping Distance, R Package Version 5.5.12; R Core Team: Vienna, Austria, 2023.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProt | Description | Fold Change | UniProt | Name | Fold Change |
|---|---|---|---|---|---|
| PSII core | PSI core | ||||
| Q31RR2 | PsbH | 0.092 | Q31NT9 | PsaF | 0.130 |
| P11004 | PsbC (CP47) | 0.210 | Q31QV2 | PsaC | 0.135 |
| P11472 | PsbO | 0.238 | P95822 | PsaL | 0.155 |
| P31094 | PsbB (CP43) | 0.241 | Q31NU0 | PsaJ | 0.161 |
| Q8KPP1 | PsbL | 0.250 | Q5MZZ8 | PsaM | 0.164 |
| P04996 | PsbA1 (D1) | 0.258 | Q31PI7 | PsaD | 0.176 |
| P11005 | PsbD (D2) | 0.267 | Q31LJ1 | PsaB | 0.195 |
| Q8KPP3 | PsbE (Cyt b559) | 0.284 | Q31LJ0 | PsaA | 0.208 |
| Q31LS7 | PsbY | 0.342 | |||
| P04997 | PsbA2 | 2.255 | |||
| PSII assembly factors | PSI assembly factors | ||||
| Q31QR0 | Psb34 | 0.300 | Q31Q43 | VIPP1 | 0.455 |
| Q31RE4 | Psb27 | 0.439 | Q31NR5 | Ycf37 | 0.631 |
| Q31ML0 | Psb28 | 0.693 | |||
| Q31KQ9 | CtpA | 0.530 | |||
| Cyt b6f | NDH-1 | ||||
| Q31NV8 | PetA | 0.236 | Q31QG9 | NdhP | 0.365 |
| Q54710 | PetD | 0.393 | Q8VPV4 | NdhF4 | 0.384 |
| Q54711 | PetB | 0.400 | Q8VPU9 | NdhD4 | 0.466 |
| Q31NV7 | PetC | 0.434 | Q31NJ3 | NdhE | 1.974 |
| Q31LY1 | NdhV | 1.978 | |||
| Q8VPV6 | CupS | 2.663 | |||
| Q8VPV9 | NdhF3 | 3.055 | |||
| Q8VPV7 | CupA | 5.060 | |||
| Q31LE7 | NdhD3 | 12.812 | |||
| PBS | |||||
| Q31PD9 | CpcL | 0.166 | |||
| Q31PE0 | CpcH | 0.259 | |||
| P06539 | CpcB1 | 0.268 | |||
| Q31PD8 | CpcD | 0.269 | |||
| Q31RF9 | ApcE | 0.295 | |||
| Q31RG0 | ApcA | 0.305 | |||
| O50209 | ApcF | 0.328 | |||
| Q31RG1 | ApcB | 0.333 | |||
| Q31LK9 | CpcG | 0.342 | |||
| Q31RP7 | ApcD | 0.416 | |||
| Q31Q37 | CpcT | 0.621 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, F.; Grauslys, A.; Huokko, T.; Caamaño Gutiérrez, E.; Jones, A.R.; Liu, L.-N. Dynamic Changes in the Thylakoid Proteome of Cyanobacteria during Light-Regulated Thylakoid Membrane Development. Plants 2023, 12, 3967. https://doi.org/10.3390/plants12233967
Huang F, Grauslys A, Huokko T, Caamaño Gutiérrez E, Jones AR, Liu L-N. Dynamic Changes in the Thylakoid Proteome of Cyanobacteria during Light-Regulated Thylakoid Membrane Development. Plants. 2023; 12(23):3967. https://doi.org/10.3390/plants12233967
Chicago/Turabian StyleHuang, Fang, Arturas Grauslys, Tuomas Huokko, Eva Caamaño Gutiérrez, Andrew R. Jones, and Lu-Ning Liu. 2023. "Dynamic Changes in the Thylakoid Proteome of Cyanobacteria during Light-Regulated Thylakoid Membrane Development" Plants 12, no. 23: 3967. https://doi.org/10.3390/plants12233967
APA StyleHuang, F., Grauslys, A., Huokko, T., Caamaño Gutiérrez, E., Jones, A. R., & Liu, L.-N. (2023). Dynamic Changes in the Thylakoid Proteome of Cyanobacteria during Light-Regulated Thylakoid Membrane Development. Plants, 12(23), 3967. https://doi.org/10.3390/plants12233967