
Hormetic Effect Caused by Sublethal Doses of Glyphosate on Toona ciliata M. Roem

 , , , , , , , , , and
, , , , , , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
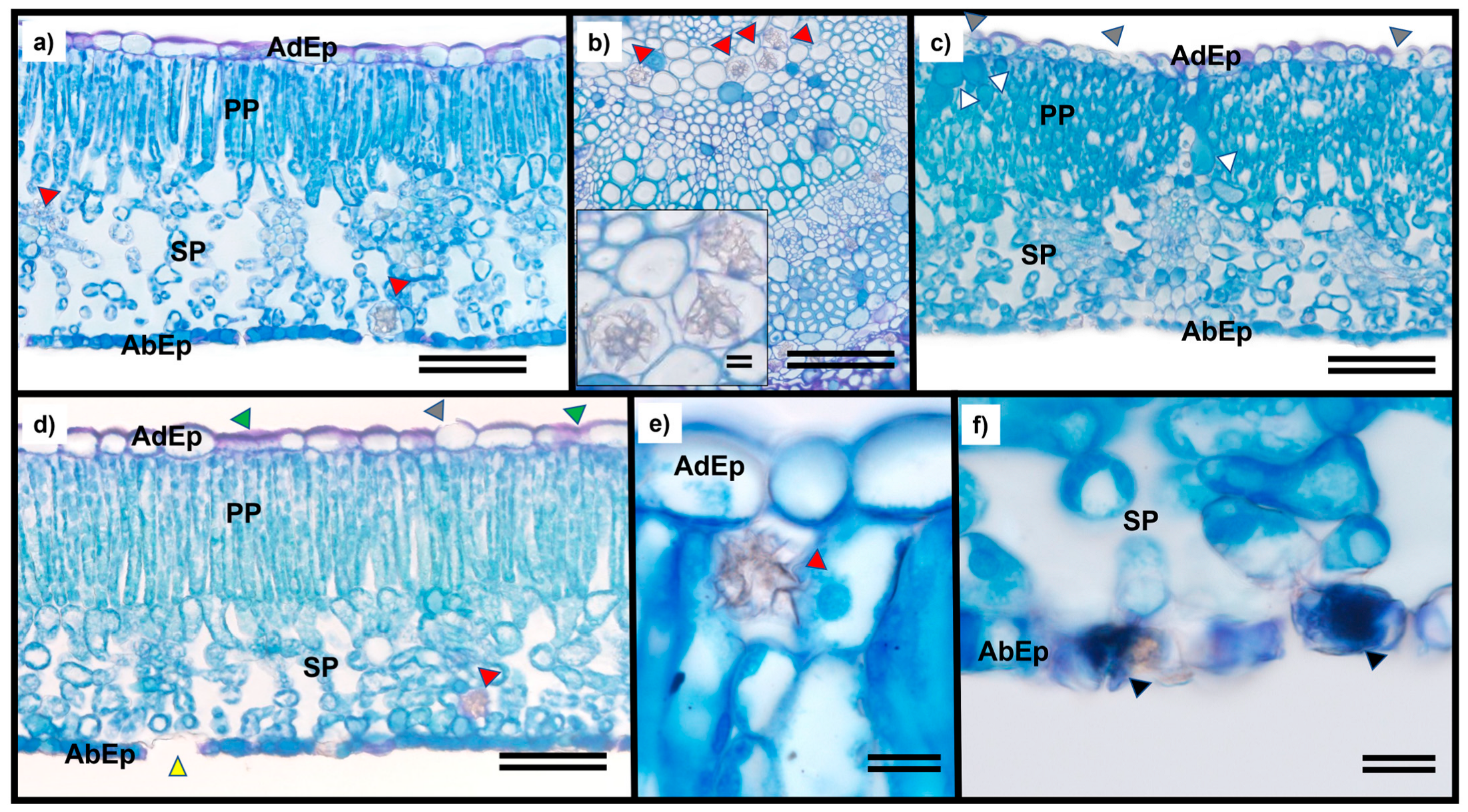
2.1. Visual Symptoms and Anatomical Assessments
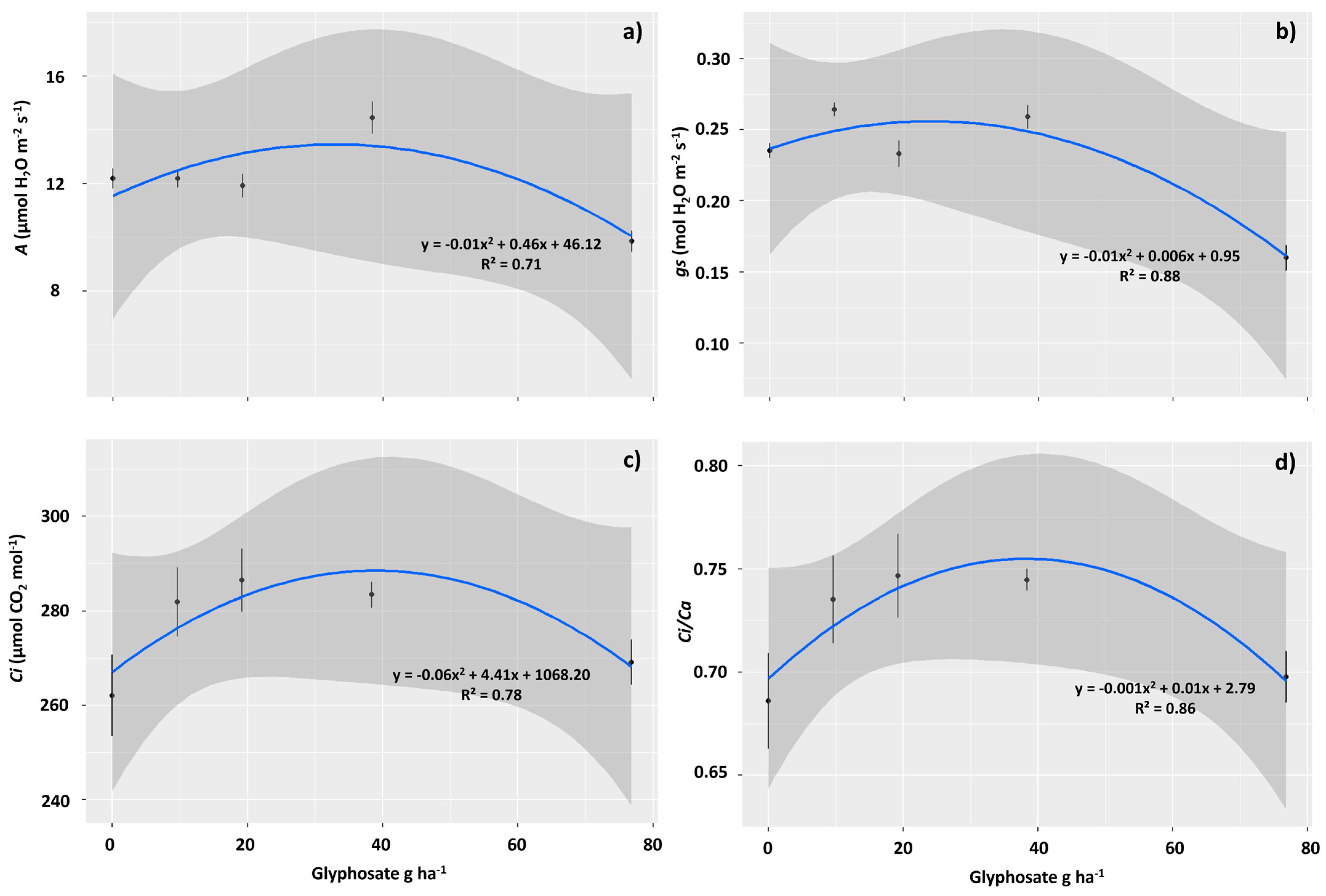
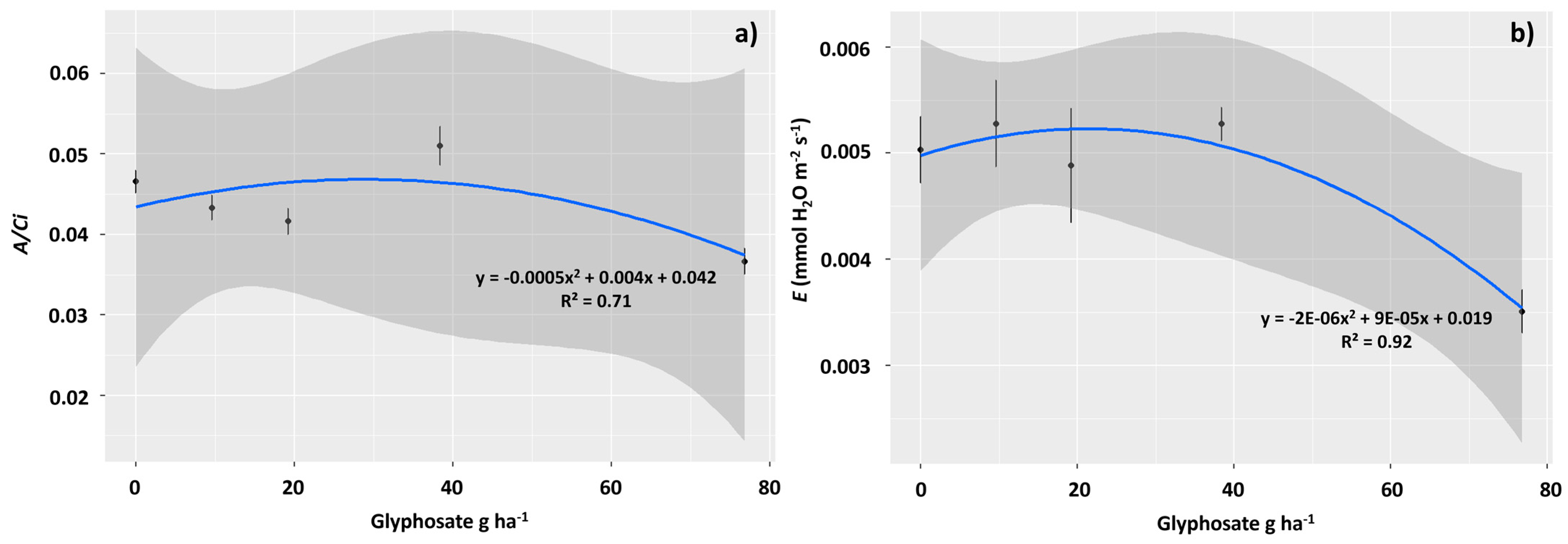
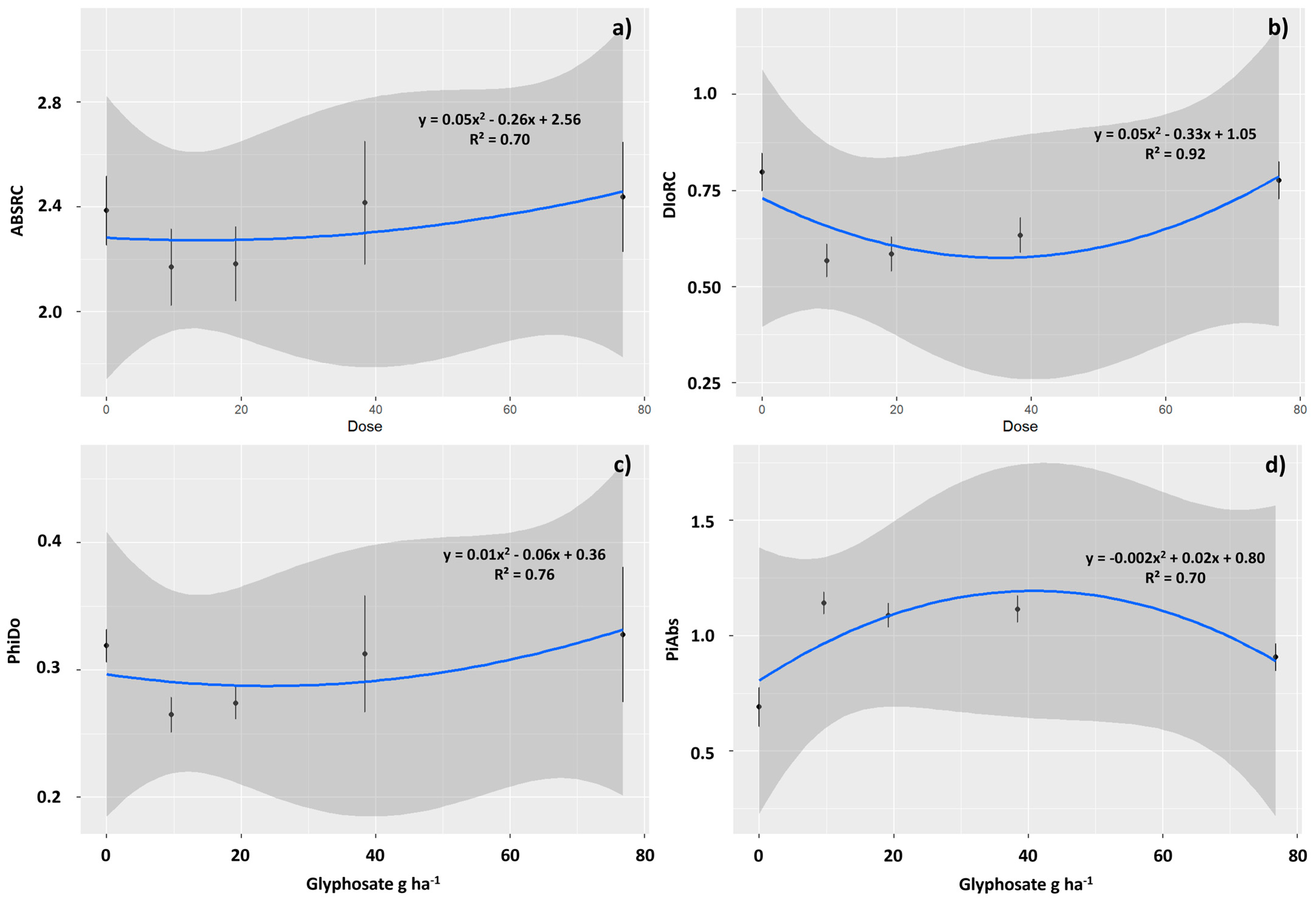
2.2. Physiological Assessments
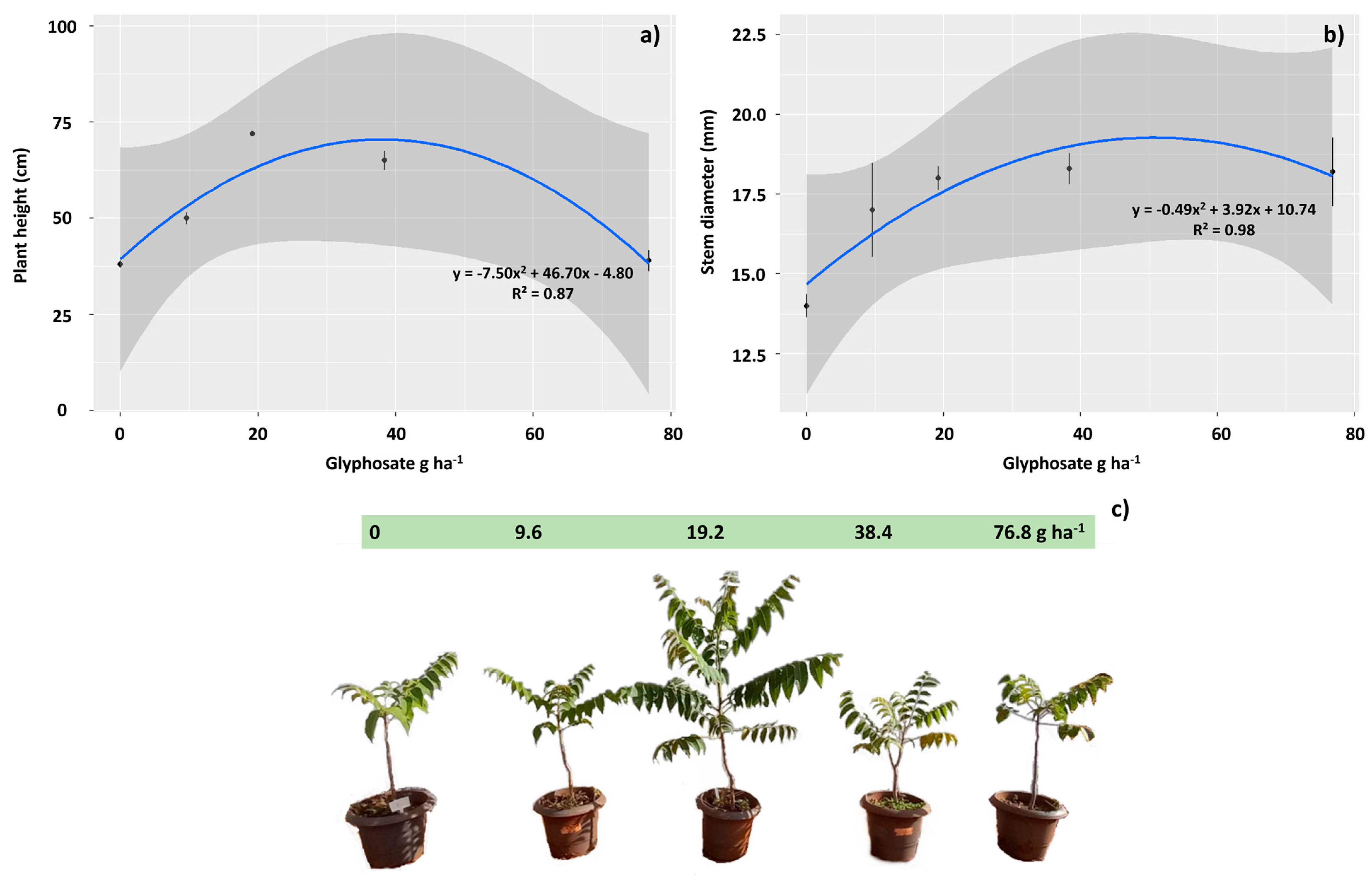
2.3. Biometric Assessments
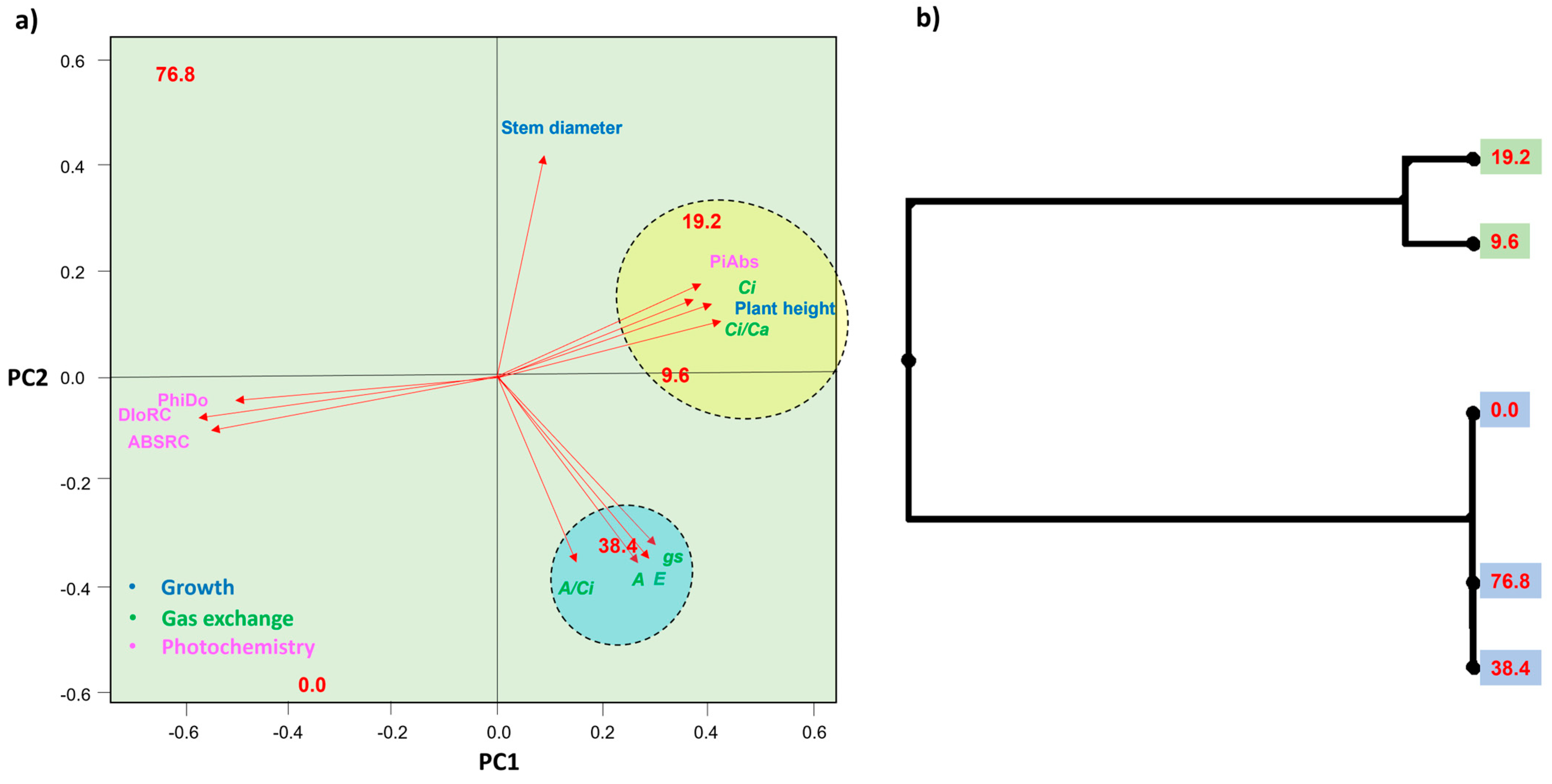
2.4. Principal Component Analysis and UPGMA
3. Discussion
3.1. Toxicity Symptoms Were Observed in T. ciliata Leaves Applying Sublethal Doses of Glyphosate above 19.2 g ha−1
3.2. The Sublethal Dose of 76.8 g ha−1 of Glyphosate Negatively Affected the Photosynthetic Performance of T. ciliata
3.3. Sublethal Doses of 9.6 and 19.2 g ha−1 of Glyphosate Improved Photochemical Performance in T. ciliata Leaves
3.4. Sublethal Doses of 9.6, 19.2 and 38.4 g ha−1 of Glyphosate Stimulate the Growth of T. ciliata Plants
4. Materials and Methods
4.1. Experimental Design and Conduct
4.2. Visible Leaf Symptoms
4.3. Leaf Morphoanatomical Characterization
4.4. Gas Exchange and Chlorophyll a Fluorescence
4.5. Growth Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liang, R.L.; Lião, R.Y.; Dai, J. Causas ameaçadas e estratégia de proteção de Toona ciliata. Guangxi Para Sci. 2011, 40, 201–203. [Google Scholar]
- Zhan, X.; Li, P.; Hui, W.; Deng, Y.; Gan, S.; Sun, Y.; Deng, X. Genetic diversity and population structure of Toona ciliata revealed by simple sequence repeat markers. Biotechnol. E Biotechnol. Equip. 2019, 33, 214–222. [Google Scholar] [CrossRef]
- Wang, X.; Xiao, Y.; He, Z.H.; Li, L.L.; Song, H.Y.; Zhang, J.J.; Hu, X.S. A Chromosome-Level Genome Assembly of Toona ciliata (Meliaceae). Genome Biol. Evol. 2022, 14, evac121. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, E.P.; Ferreira, P.A.A.; Furtini Neto, A.E.; Soares, C.R.F.S. Arbuscular mycorrhiza and phosphate on growth of australian red cedar seedlings. Ciênc. Florest. 2017, 27, 1269–1281. [Google Scholar] [CrossRef]
- Dordel, J.; Seely, B.; Simard, S.W. Relationships between simulated water stress and mortality and growth rates in underplanted Toona ciliata Roem. in subtropical Argentinean plantations. Ecol. Model. 2011, 222, 3226–3235. [Google Scholar] [CrossRef]
- Mirza, A.U.; Khan, M.S.; Abdul Kareem, A.A.; Nami, S.A.; Bhat, S.A.; Singh, M.p.; Nahid Nishat, N. Biomediated synthesis, characterization, and biological applications of nickel oxide nanoparticles derived from Toona ciliata, Ficus carica and Pinus roxburghii. Bioprocess Biosyst. Eng. 2021, 44, 1461–1476. [Google Scholar] [CrossRef]
- Souza, C.O.M.; Cruz, J.C.; Cesila, C.A.; Gonzalez, N.; Rocha, B.A.; Adeyemi, J.Á.; Barbosa, F. Recent trends in pesticides in crops: A critical review of the duality of risks-benefits and the Brazilian legislation issue. Pesq. Ambient. 2023, 228, 115811. [Google Scholar] [CrossRef]
- Zavariz, A.; Berryhill, Q.T.A.; Guimarães, E.T.; Pereira, F.A.C. The use of glyphosate in coffee growing, an epistemological study. Braz. J. Dev. 2020, 6, 36046–36058. [Google Scholar] [CrossRef]
- Brovini, E.M.; De Deus, B.C.T.; Vilas Boas, J.Á.; Quadra, G.R.; Carvalho, L.; Mendonca, R.F.; Cardoso, S.J. Three-bestseller pesticides in Brazil: Freshwater concentrations and potential environmental risks. Sci. Total Environ. 2021, 771, 144754. [Google Scholar] [CrossRef]
- Bondarenko, A.; Tyutyuma, N. Biological effectiveness of onion plant protection scheme. Rev. Agric. Neotrop. 2022, 9, e6930. [Google Scholar] [CrossRef]
- Duke, S.O. Herbicides: Chemistry, Degradation and Mode of Action; Kearney, P.C., Kaufman, D.D., Eds.; Marcel Dekker: New York, NY, USA, 1988; pp. 1–70. [Google Scholar]
- Duke, S.O. Secondary effects of glyphosate on plants. In A Era Glyphosate: Agricultura, Meio Ambiente e Homem; Meschede, D.K., Gazziero, D., Eds.; Midiograph: Londrina, Brazil, 2016; pp. 87–96. [Google Scholar]
- De Lima Prieto, F.; Augustinho, B.; De Toledo, E.A. Produtividade da aveia branca submetida a dessecação pré colheita. Rev. Cult. Saber. 2023, Mercado e Pesquisa, 1–9. Available online: https://cultivandosaber.fag.edu.br/index.php/cultivando/article/view/1189 (accessed on 11 August 2022).
- Parreira, M.L.; Côrrea, F.R.; Da Silva, N.F.; Da Silva Cavalcante, W.S.; Ribeiro, D.F.; Rodrigues, E. Herbicides with potential for desiccation of the area in pre-sowing of the soybean crop. Braz. J. Sci. 2023, 2, 46–59. [Google Scholar] [CrossRef]
- Mulla, S.I.; Ameen, F.; Talwar, M.P.; Eqani, S.A.M.A.S.; Bharagava, R.N.; Saxena, G.; Tallur, P.N.; Ninnekar, H.Z. Organophosphate pesticides: Impact on environment, toxicity, and their degradation. In Bioremediation of Industrial Waste for Environmental Safety: Volume I: Industrial Waste and Its Management; Saxena, G., Bharagava, R., Eds.; Springer: Berlin, Germany, 2020; pp. 265–290. [Google Scholar]
- de Jesus, R.A.; Barros, G.P.; Bharagava, R.N.; Liu, J.; Mulla, S.I.; Azevedo, L.C.B.; Ferreira, L.F.R. Occurrence of pesticides in wastewater: Bioremediation approach for environmental safety and its toxicity. In Advances in Chemical Pollution, Environmental Management and Protection; Elsevier: Amsterdam, The Netherlands, 2023; pp. 17–33. [Google Scholar] [CrossRef]
- De Araújo, E.P.; Silva, K.L.F. Morphoanatomical responses of Bowdichia virgilioides (Fabaceae) to glyphosate drift in an agroecosystem of the Brazilian cerrado Ibero-Am. J. Environ. Sci. 2021, 12, 112–124. [Google Scholar]
- Nassur, O.A.C.; Rosado, L.R.; Rosado, S.C.S.; De Carvalho, P.M. Variations in log quality of eighteen years old Toona ciliata M. Roem. Cerne 2013, 19, 43–49. [Google Scholar] [CrossRef]
- Alves, J.Á.; Calegario, N.; Rosado, S.C.S.; Silva, G.A.; Possato, E.L.; Melo, E.A. Hypsometric equations for Toona ciliata with inclusion of covariates. Ciênc. Florest. 2017, 27, 581–595. [Google Scholar] [CrossRef]
- Nunes, M.T.A. Produção e Sortimento de Madeira de Toona ciliata var. Australis Cultivada no Entorno de Cafezais. Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, Brasil, 2020. [Google Scholar]
- De Sá, L.C.; Lazarotto, M.; Avrella, E.D.; Hilgert, M.A.; Fior, C.S. Estresse hídrico e salino no crescimento inicial de mudas de Toona ciliata var. australis. Pesq. Florest. Bras. 2023, 43, e202002108. [Google Scholar] [CrossRef]
- Andrade, R.S.D.; Navroski, M.C.; Pereira, M.D.O.; Sá, A.C.S. Morphological and physiological variation in Toona ciliata under water and salinity stress. Cienc. Rural 2020, 50. [Google Scholar] [CrossRef]
- Ethridge, S.R.; Chandra, S.; Locke, A.M.; Everman, W.J.; Jordan, D.L.; Owen, M.D.; Leon, R.G. Changes in herbicide sensitivity and competitive ability of Abutilon theophrasti over 28 years: Implications for hormesis and weed evolution. Pest Manag. Sci. 2023, 79, 4048–4056. [Google Scholar] [CrossRef]
- Xu, N.; Sun, Y.; Wang, Y.; Cui, Y.; Jiang, Y.; Zhang, C. Hormesis effects in tomato plant growth and photosynthesis due to acephate exposure based on physiology and transcriptomic analysis. Pest Manag. Sci. 2023, 79, 2029–2039. [Google Scholar] [CrossRef]
- Milosevic, L.; Osipitan, O.A.; Scott, J.E.; Knezevic, S.Z. Soybean Tolerance to ultra-low doses of dicamba: Hormesis or not. Crop Prot. 2023, 173, e106356. [Google Scholar] [CrossRef]
- Vilela, E.S.; Stehling, E.D.C. Recomendações de Plantio de Para o Cedro Australiano; Cedro Australiano; Bela Vista Florestal: Campo Belo, Brazil, 2015. [Google Scholar]
- Cedergreen, N. Is the growth stimulation by low doses of glyphosate sustained over time? Environ. Pollut. 2008, 156, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.E.S.; Freitas Silva, L.; Ribeiro, C.; Silva, L.C. Physiological and morphoanatomical effects of glyphosate in Eugenia uniflora, a Brazilian plant species native to the Atlantic Forest biome. Environ. Sci. Pollut. Res. 2021, 28, 21334–21346. [Google Scholar] [CrossRef] [PubMed]
- Santos, V.R.S.; Crispim Filho, A.J.; Santana, M.M.; Costa, A.C.; Silva, K.L.F. Análises fisiológicas e morfoanatômicas de Cenostigma macrophyllum tul. (fabaceae) submetida a diferentes concentrações de glifosato. Rev. IberoAm. Cienc. Ambient. 2020, 11, 159–173. [Google Scholar] [CrossRef]
- Vicentini, T.A. Respostas Morfoanatômicas e Fisiológicas de Duas Espécies Arbóreas sob Deriva Simulada do Herbicida Glifosato. Ph.D. Thesis, Instituto Federal Goiano Rio Verde Goiás, Rio Verde, Brazil, 2023. [Google Scholar]
- Jacinto Júnior, S.G.; Moraes, J.G.L.; Silva, F.D.B.D.; Silva, B.D.N.; Sousa, G.G.D.; Oliveira, L.L.B.D.; Mesquita, R.O. Respostas fisiológicas de genótipos de fava (Phaseolus lunatus L.) submetidas ao estresse hídrico cultivadas no Estado do Ceará. Rev. Bras. Meteorol. 2019, 34, 413–422. [Google Scholar] [CrossRef]
- Carloto, B.W. Alterações Fenotípicas de Duas Espécies de Eragrostis Wolf. (Poaceae) sob Diferentes Condições de Umidade do Solo. Ph.D. Thesis, Universidade Federal de Santa Maria, Santa Maria, Brazil, 2016. [Google Scholar]
- Trivisiol, V.S. Espécies de Capim Arroz Submetidas às Diferentes Doses de Herbicidas, Condições de Umidade do Solo e Adubação Nitrogenada. Ph.D. Thesis, Universidade Federal de Santa Maria, Santa Maria, Brazil, 2020. [Google Scholar]
- Belz, R.G.; Duke, S.O. Modelling biphasic hormetic dose responses to predict sub-NOAEL effects using plant biology as an example. Curr. Opin. Toxicol. 2022, 29, 36–42. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Agathokleous, E. Nitric oxide, hormesis and plant biology. Sci. Total Environ. 2023, 29, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Zobiole, L.H.S.; Kremer, R.J.; Oliveira, R.S.; Junior Constantin, J. Glyphosate affects chlorophyll, nodulation and nutrient accumulation of “second generation” glyphosate-resistant soybean (Glycine max L.). Pest Biochem. Physiol. 2011, 99, 53–60. [Google Scholar] [CrossRef]
- Oliveira, A.P.A.D.; Crispim Filho, A.J.; Santos, V.R.S.; Costa, A.C.; Silva, K.L.F. Physiological and morphoanatomic responses of Bowdichia virgilioides kunth. (Fabaceae) to glyphosate. Rev. Árvore. 2021, 45. [Google Scholar] [CrossRef]
- Bianchi, A.; Bianchi, G. Surface lipid composition of C3 and C4 plants. Biochem. Syst. Ecol. 1990, 18, 533–537. [Google Scholar] [CrossRef]
- Dos Reis, F.C.; Mendes, K.F.; Baccin, L.; Takeshita, V.; Tornisielo, V.L.; Filho, R.V. Selectivity, hormesis and physiology of herbicides in plants. In Matologia: Estudos Sobre Plantas Daninhas; Fábrica da Palavra: Jaboticabal, Brazil, 2021; pp. 295–323. Available online: https://www.researchgate.net/profile/Kassio-Mendes-/publication/349173646_SELETIVIDADE_HORMESIS_E_FISIOLOGIA_DOS_HERBICIDAS_NAS_PLANTAS/links/6023cc16a6fdcc37a818675a/SELETIVIDADE-HORMESIS-E-FISIOLOGIA-DOS-HERBICIDAS-NAS-PLANTAS.pdf (accessed on 11 August 2022).
- Yilmaz, G.; Dane, F. Phytotoxicity Induced by Herbicide and Surfactant on stomata and epicuticular wax of Wheat. Rom. Biotech. Lett. 2012, 6, 7757–7765. [Google Scholar]
- Mota, L.M.; Mendes, K.F.; Júnior, L.H.B.; Silva, E.M.G.; Furtado, I.F.; Tornisielo, V.L. Light availability interferes with absorption and translocation of 14C-glyphosate in Urochloa brizantha cv. Marandu plants. Radioanal. Nucl. Chem. 2020, 326, 683–693. [Google Scholar] [CrossRef]
- De Paiva Ferreira, G.A. Influence of Light Intensity on Chemical Control of Weeds. Ph.D. Thesis, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil, 2021. [Google Scholar]
- Silva, L.Q.; Jakelaitis, A.; Vasconcelos Filho, S.C.; Costa, A.C.; Araújo, A.C.F. Morpho-anatomical changes of pequi leaves (Caryocar brasiliense Cambess.) exposed to simulated drift of glyphosate. Rev. Árvore. 2016, 40, 669–677. [Google Scholar] [CrossRef]
- Freitas-Silva, L.; de Araújo, T.O.; Nunes-Nesi, A.; Ribeiro, C.; Costa, A.C.; da Silva, L.C. Evaluation of morphological and metabolic responses to glyphosate exposure in two neotropical plant species. Ecol. Indic. 2020, 113, 106246. [Google Scholar] [CrossRef]
- Zini, A.D.S.; Martins, S.; Toderke, M.L.; Temponi, L.G. Foliar anatomy of Rubiaceae occurring in an urban forest fragment of Atlantic Forest, Paraná State, Brazil. Hoehnea 2015, 43, 173–182. [Google Scholar] [CrossRef]
- Nakata, P.A. advances in our understanding of calcium oxalate crystal formation and function in plants. Plant Sci. 2003, 164, 901–909. [Google Scholar] [CrossRef]
- Messerli, M.A.; Creton, R.; Jaffe, L.F.; Robinson, K.R. Periodic Increases in elongation rate precede increase in cytosolic Ca2+ during pollen tube growth. Dev. Biol. 2000, 222, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Holdaway Clarke, T.L.; Weddle, N.M.; Kim, S.; Robia, A.; Parris, C.; Kunkel, J.G.; Hepler, P.K. Effect of extracellular calcium, pH and borate on growth oscillations in Lilium formosanum pollen tubes. J. Exp. Bot. 2003, 54, 65–72. [Google Scholar] [CrossRef]
- Appezzato Da Glória, B.; Carmello Guerreiro, S.M. Anatomia Vegetal; UFV: Viçosa, Brazil, 2006. [Google Scholar]
- Carneiro, E.B.; Smiljanic, K.B. Caracterização Morfoanatômica de Portulaca oleracea L.; Universitário de Mineiros (UNIFIMES): Mineiros, Brazil, 2016. [Google Scholar]
- Lima, É.R.; Silva, R.A.D.; Pereira, G.M.; Moreira, F.G.L.; Silva, A.N.E.; Lichston, J.E. Anthurium affine schott (araceae): Análise anatômica e histoquímica para identificação de estruturas secretoras. Rev. Ibero-Am. Ciênc. Ambient. 2020, 11, 92–98. [Google Scholar] [CrossRef]
- Cuéllar Cruz, M.; Pérez, K.S.; Mendoza, M.E.; Moreno, A. Biocrystals in plants: A short review on biomineralization processes and the role of phototropins into the uptake of calcium. Crystals 2020, 10, e591. [Google Scholar] [CrossRef]
- Cavalcante, H.S.; Dos Santos, M.C.P.; Da Silva Brito, G.S.M.; Teixeira, V.I.; Pereira, C.F.; Andrade, A.P.; Da Silva, D.S. Anatomia foliar de genótipos do gênero Desmanthus. Res. Soc. Dev. 2021, 16, e418101623776. [Google Scholar] [CrossRef]
- Pereira, E.R.D.L.; Medeiros, M.B.D.; Suassuna, J.F.; Morais, V.M.M.; Fernandes, H.F.; Lima, A.S.D.; Fernandes, P.D. Resposta fisiológica do algodão colorido em estresse salino. Rev. Bras. Gest. Ambient. Sustent. 2020, 7, 653–664. [Google Scholar] [CrossRef]
- Palharini, K.M.Z.; Vitorino, L.C.; Bessa, L.A.; De Carvalho Vasconcelos Filho, S.; Silva, F.G. Parmotrema tinctorum as an indicator of edge effect and air quality in forested areas bordered by intensive agriculture. Environ. Sci. Pollut. Res. 2021, 28, 68997–69011. [Google Scholar] [CrossRef] [PubMed]
- Maynard, K.I. Hormesis pervasiveness and its potential implications for pharmaceutical research and development. Dose Response 2011, 9, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Abbas, T.; Tanveer, A.; Maqbool, R.; Zohaib, A.; Shehzad, M.A. Glyphosate hormesis in broad-leaved weeds: A challenge for weed management. Arch. Agron. Soil Sci. 2017, 63, 344–351. [Google Scholar] [CrossRef]
- Abbas, T.; Nadeem, M.A.; Tanveer, A.; Zohaib, A.; Rasool, T. Glyphosate hormesis increases growth and yield of chickpea (Cicer arietinum L.). Pak. J. Weed Sci. Res. 2015, 21, 533–542. [Google Scholar]
- Brito, I.P.; Tropaldi, L.; Carbonari, C.A.; Velini, E.D. Hormetic effects of glyphosate on plants. Pest Manag. Sci. 2018, 74, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Jalal, A.; de Oliveira Junior, J.C.; Ribeiro, J.S.; Fernandes, G.C.; Mariano, G.G.; Trindade, V.D.R.; Dos Reis, A.R. Hormesis in plants: Physiological and biochemical responses. Ecotoxicol. Environ. Saf. 2021, 207, 111225. [Google Scholar] [CrossRef]
- Da Silva, T.F.; Do Carmo, K.B.; Armiliato, N. Toxicidade celular do herbicida glifosato sobre os ovários do peixe Danio rerio. Saúde Meio Ambiente Ver. Interdiscip. 2019, 8, 1–12. [Google Scholar] [CrossRef]
- Azul, A.S.S. Biomonitorização de Glifosato em Urina Numa População do Baixo Mondego. Ph.D. Thesis, Universidade de Coimbra, Coimbra, Brazil, 2021. [Google Scholar]
- Moustaka, G.; Ouzounidou, I.; Sperdouli, M.; Moustakas, J. Photosystem II is more sensitive than photosystem I to Al3+ induced phytotoxicity. Materials 2018, 11, 1772. [Google Scholar] [CrossRef]
- Li, F.; Xing, D.; Xing, Z. Characterization of target site of aluminum phytotoxicity in photosynthetic electron transport by fluorescence techniques in tobacco leaves. Plant Cell Physiol. 2012, 53, 1295–1309. [Google Scholar] [CrossRef]
- Moustakas, M.; Moustaka, J.; Sperdouli, I. Hormesis in photosystem II: A mechanistic understanding. Curr. Opin. Toxicol. 2022, 29, 57–64. [Google Scholar] [CrossRef]
- Costa, R.N.; Bevilaqua, N.D.C.; Krenchinski, F.H.; Giovanelli, B.F.; Pereira, V.G.C.; Velini, E.D.; Carbonari, C.A. Hormetic Effect of Glyphosate on the Morphology, Physiology and Metabolism of Coffee Plants. Plants 2023, 12, 2249. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, O.M.; Silva, G.B.; Neto, R.M.R.; Campos, O.R.; Peres, W.M. Interferência de subdoses de glyphosate no desenvolvimento de plantas jovens de nim. Nativa 2017, 5, 163–268. [Google Scholar] [CrossRef]
- Marques, K.D.M.; De Lima Moreira, W.C.; De França Silva, J.; Do Vale Moreira, J.G.; Melhorança Filho, A.L. Hormetic effect of glyphosate on the initial growth of paricá seedlings (Schizolobium amazonicum). Agrarian 2020, 13, 9–16. [Google Scholar] [CrossRef]
- Da Silva, A.C.S.; Caioni, C.; da Veiga, J.B.; Neto, R.M.R.; De Oliveira, A.S. Aerial biomass stock, carbon and carbon dioxide sequestration in forest systems in the amazon mato-grossense. Acta Iguazu 2014, 3, 22–35. [Google Scholar]
- Carlos, L.; Venturin, N.; Macedo, R.L.G.; Higashikawa, E.M.; Garcia, M.B.; Farias, E.D.S. Growth and mineral nutrition in seedlings of pequi subjected to nutrient deprivation. Ciênc. Florest. 2014, 24, 13–21. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Fluorescência da clorofila—Um guia prático. J. Bot. Exp. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanisms, Regulation and Adaptation; CRC Press: Boca Raton, FL, USA, 2000; pp. 445–483. [Google Scholar]
- Delarmelina, W.M.; Caldeira, M.V.W.; Faria, J.C.T.; Gonçalves, E.D.O.; Rocha, R.L.F. Different substrates for the production of Sesbania virgata seedlings. Floresta Environ. 2014, 21, 224–233. [Google Scholar] [CrossRef]
- Portes, T.A.; Castro Junior, L.D. Análise de crescimento de plantas: Um programa computacional auxiliar1. Rev. Bras. Fisiol. Veg. 1991, 3, 53–56. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 27 March 2022).
- Sokal, R.R.; Rohlf, F.J. The comparison of dendrograms by objective methods. Taxon 1962, 11, 33–40. [Google Scholar] [CrossRef]
- Garcia-Vallve, S.; Palau, J.; Romeu, A. Horizontal gene transfer in glycosyl hydrolases inferred from codon usage in Escherichia coli and Bacillus subtilis. Mol. Biol. Evol. 1999, 16, 1125–1134. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Faria, G.S.; Carlos, L.; Jakelaitis, A.; de Freitas, S.T.F.; Vicentini, T.A.; Silva, I.O.F.; Vasconcelos Filho, S.C.; Lourenço, L.L.; Farnese, F.S.; Batista, M.A.; et al. Hormetic Effect Caused by Sublethal Doses of Glyphosate on Toona ciliata M. Roem. Plants 2023, 12, 4163. https://doi.org/10.3390/plants12244163
de Faria GS, Carlos L, Jakelaitis A, de Freitas STF, Vicentini TA, Silva IOF, Vasconcelos Filho SC, Lourenço LL, Farnese FS, Batista MA, et al. Hormetic Effect Caused by Sublethal Doses of Glyphosate on Toona ciliata M. Roem. Plants. 2023; 12(24):4163. https://doi.org/10.3390/plants12244163
Chicago/Turabian Stylede Faria, Giselle Santos, Leandro Carlos, Adriano Jakelaitis, Samylla Tassia Ferreira de Freitas, Taíza Andressa Vicentini, Igor Olacir Fernandes Silva, Sebastião Carvalho Vasconcelos Filho, Lucas Loram Lourenço, Fernanda Santos Farnese, Marco Aurélio Batista, and et al. 2023. "Hormetic Effect Caused by Sublethal Doses of Glyphosate on Toona ciliata M. Roem" Plants 12, no. 24: 4163. https://doi.org/10.3390/plants12244163
APA Stylede Faria, G. S., Carlos, L., Jakelaitis, A., de Freitas, S. T. F., Vicentini, T. A., Silva, I. O. F., Vasconcelos Filho, S. C., Lourenço, L. L., Farnese, F. S., Batista, M. A., & Vitorino, L. C. (2023). Hormetic Effect Caused by Sublethal Doses of Glyphosate on Toona ciliata M. Roem. Plants, 12(24), 4163. https://doi.org/10.3390/plants12244163