

Impact of Chill and Heat Exposures under Diverse Climatic Conditions on Peach and Nectarine Flowering Phenology

 ,
,  , ,
, , 
 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
2.2. Bloom Characterization
2.3. Chill and Heat Models
2.4. Estimated Dates of Dormancy Release and Bloom
2.5. Statistical Analyses and Model Validation
3. Results and Discussion
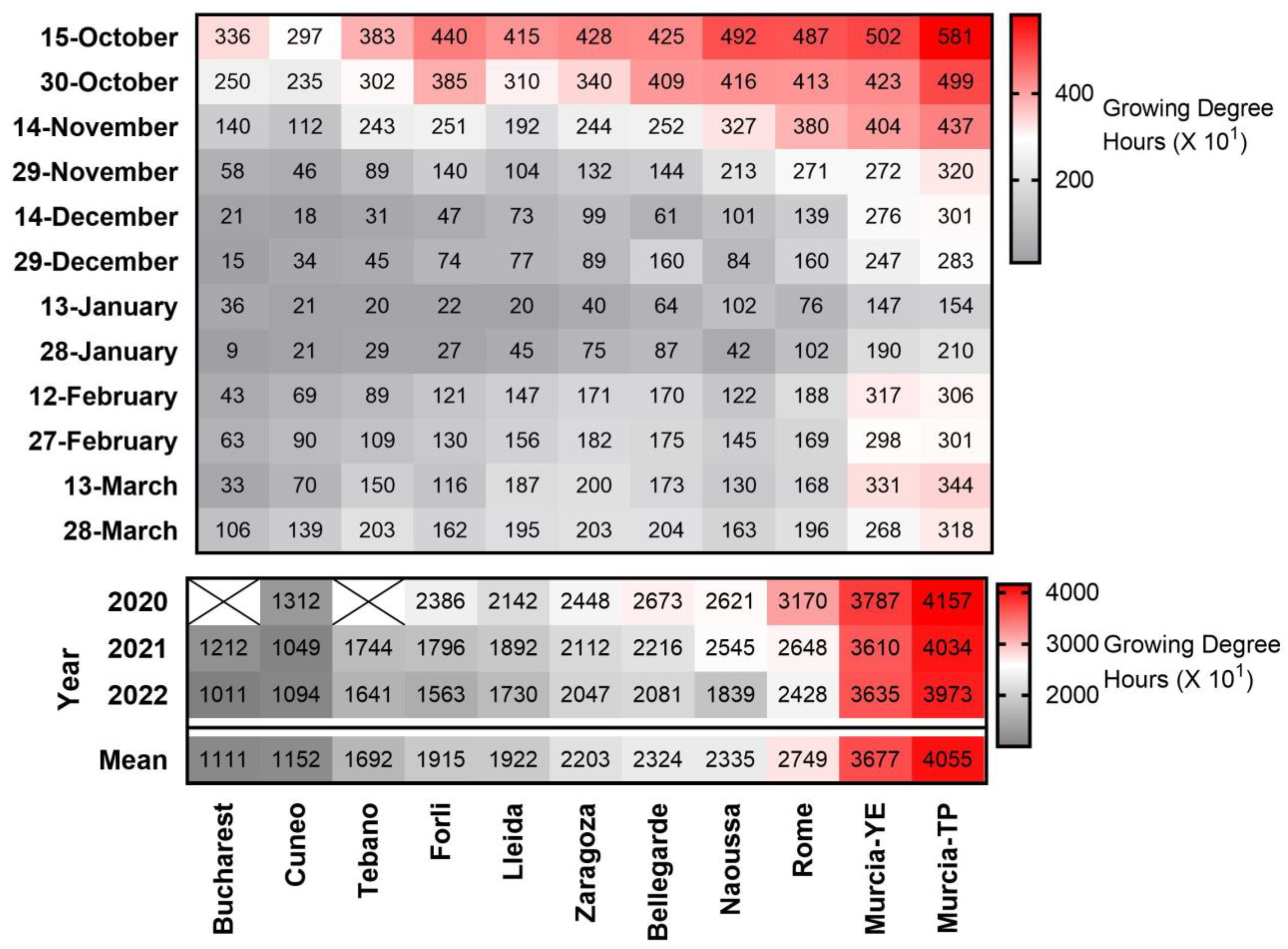
3.1. Chill and Heat Accumulation
3.2. Rest Completion
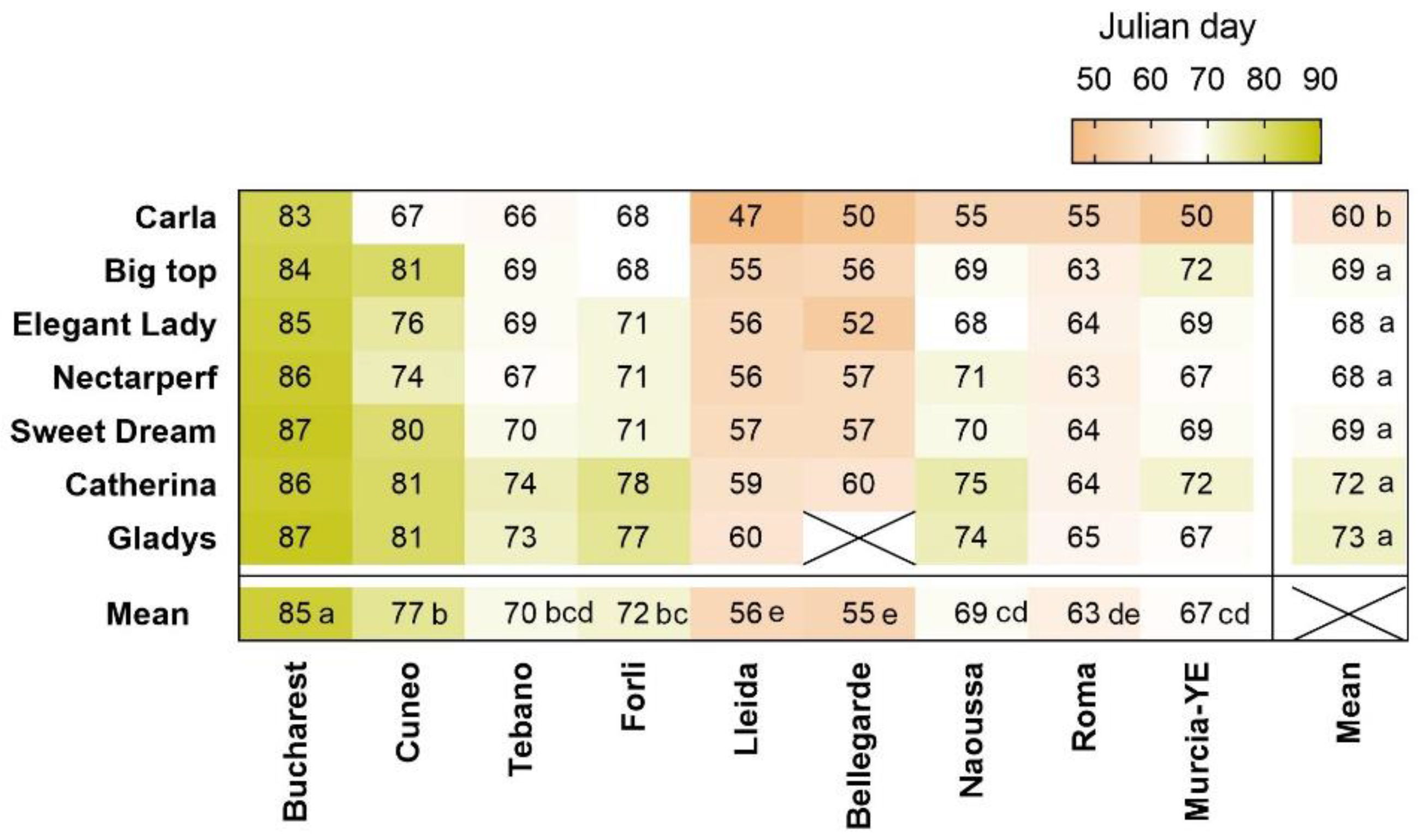
3.3. Bloom Dates
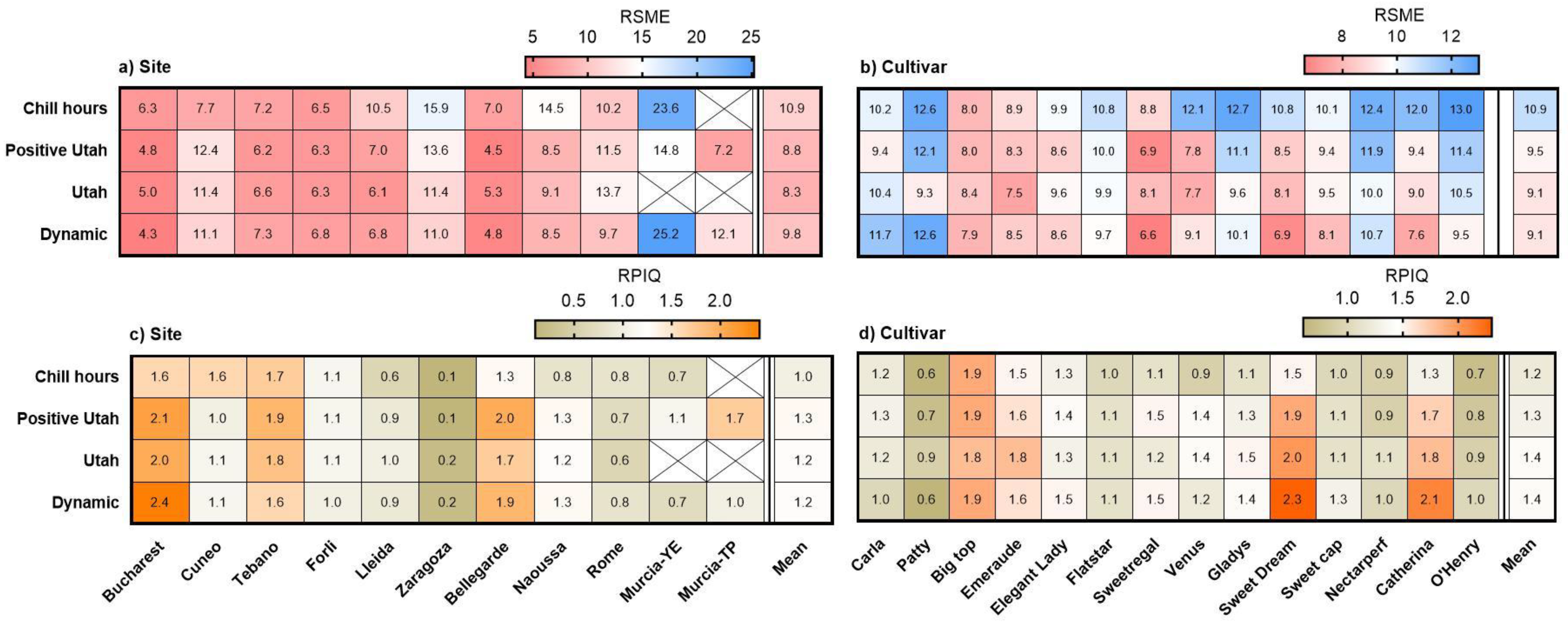
3.4. Observed vs. Estimated Bloom Dates
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Cantin, C.M.; Wang, X.W.; Almira, M.; Arús, P.; Eduardo, I. Inheritance and QTL analysis of chilling and heat requirements for flowering in an interspecific almond x peach (Texas x Earlygold) F2 population. Euphytica 2020, 216, 51. [Google Scholar] [CrossRef]
- Fadón, E.; Herrera, S.; Guerrero, B.I.; Guerra, M.E.; Rodrigo, J. Chilling and Heat Requirements of Temperate Stone Fruit Trees (Prunus sp.). Agronomy 2020, 10, 409. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Fang, W.C.; Zhu, G.R.; Cao, K.; Chen, C.W.; Wang, X.W.; Wang, L.R. Accumulated chilling hours during endodormancy impact blooming and fruit shape development in peach (Prunus persica L.). J. Integr. Agric. 2016, 15, 1267–1274. [Google Scholar] [CrossRef] [Green Version]
- Pawasut, A.; Fujishige, N.; Yamane, K.; Yamaki, Y.; Honjo, H. Relationships between chilling and heat requirement for flowering in ornamental peaches. J. Jpn. Soc. Hort. Sci. 2004, 73, 519–523. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, D.; Campoy, J.A.; Egea, J. Chilling and heat requirements of apricot cultivars for flowering. Environ. Exp. Bot. 2007, 61, 254–263. [Google Scholar] [CrossRef]
- Li, Y.; Wang, L.; Zhu, G.; Fang, W.; Cao, K.; Chen, C.; Wang, X.; Wang, X. Phenological response of peach to climate change exhibits a relatively dramatic trend in China; 1983–2012. Sci. Hortic. 2016, 209, 192–200. [Google Scholar] [CrossRef]
- Benmoussa, H.; Ghrab, M.; Ben Mimoun, M.; Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agric. For. Meteorol. 2017, 239, 34–46. [Google Scholar] [CrossRef]
- Benmoussa, H.; Luedeling, E.; Ghrab, M.; Yahmed, J.B.; Ben Mimoun, M. Performance of pistachio (Pistacia vera L.) in warming Mediterranean orchards. Environ. Exp. Bot. 2017, 140, 76–85. [Google Scholar] [CrossRef]
- Tabuenca, M.C. Necesidades de frío invernal de variedades de albaricoquero, melocotonero y peral. An. Estac. Exp. Aula Dei 1964, 7, 113–132. [Google Scholar]
- Campoy, J.A.; Ruiz, D.; Allderman, L.; Cook, N.; Egea, J. The fulfilment of chilling requirements and the adaptation of apricot (Prunus armeniaca L.) in warm winter climates: An approach in Murcia (Spain) and the Western cape (South Africa). Eur. J. Agron. 2012, 37, 43–55. [Google Scholar] [CrossRef]
- Alburquerque, N.; García-Montiel, F.; Carrillo, A.; Burgos, L. Chilling and heat requirements of sweet cherry cultivars and the relationship between altitude and the probability of satisfying the chill requirements. Environ. Exp. Bot. 2008, 64, 162–170. [Google Scholar] [CrossRef]
- Azizi Gannouni, T.; Campoy, J.A.; Quero-García, J.; Barreneche, T.; Arif, A.; Albouchi, A.; Ammari, Y. Dormancy related traits and adaptation of sweet cherry in northern Africa: A case of study in two Tunisian areas. Sci. Hortic. 2017, 219, 272–279. [Google Scholar] [CrossRef]
- Pantelidis, G.; Reig, G.; Cantín, C.M.; Giovannini, D.; Drogoudi, P. Estimation of chilling and heat requirements of peach and nectarine cultivars grown in the EUFRIN trials located in different pedoclimatic conditions. Acta Hortic. 2022, 1352, 463–470. [Google Scholar] [CrossRef]
- Giovannini, D.; Bassi, D.; Cutuli, M.; Drogoudi, P.; Foschi, S.; Hilaire, C.; Liverani, A.; Ruesch, J.; Iglesias, I. Evaluation of novel peach cultivars in the European Union: The EUFRIN Peach and Apricot Working Group initiative. Acta Hortic. 2021, 1304, 13–19. [Google Scholar] [CrossRef]
- Meier, U.; Graf, H.; Hess, M.; Kennel, K.; Klose, R.; Mappes, D.; Seipp, D.; Stauss, R.; Streif, J.; van den Boom, T. Phänologische Entwick-lungsstadien des Kernobstes (Malus domestica Borkh. und Pyrus communis L.); des Steinobstes (Prunus-Arten); der Johannisbeere (Ribes-Arten) und der Erdbeere (Fragaria x ananassa Duch.). Nachrichtenbl. Deut. Pflanzenschutzd 1994, 46, 141–153. [Google Scholar]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature dependence of dormancy breaking in plants—Computer simulation of processes studied under controlled temperatures. J. Theor. Biol. 1987, 126, 309–321. [Google Scholar] [CrossRef]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature dependence of dormancy breaking in plants—Mathematical analysis of a two-step model involving a cooperative transition. J. Theor. Biol. 1987, 124, 473–483. [Google Scholar] [CrossRef]
- Richardson, E.A.; Seeley, S.D.; Walker, D.R. A model for estimating the completion of rest for “Redhaven” and “Elberta” peach trees. Hortic. Sci. 1974, 9, 331–332. [Google Scholar]
- Linsley-Noakes, G.C.; Allan, P. Comparison of two models for the prediction of rest completion in peaches. Sci. Hortic. 1994, 59, 107–113. [Google Scholar] [CrossRef]
- Weinberger, J.H. Chilling requirements of peach varieties. Proc. Am. Soc. Hortic. Sci. 1950, 56, 122–128. [Google Scholar]
- Luedeling, E. Climate change impacts on winter chill for temperate fruit and nut production: A review. Sci. Hortic. 2012, 144, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Okie, W.R. Handbook of Peach and Nectarine Varieties; US Department of Agriculture, Agricultural Research Service: Washington, DC, USA, 1998; No. 714; 808p. [Google Scholar]
- Luedeling, E. Interpolating hourly temperatures for computing agroclimatic metrics. Int. J. Biometeorol. 2018, 62, 1799–1807. [Google Scholar] [CrossRef] [PubMed]
- Bellon-Maurel, V.; Fernández-Ahumada, E.; Palagos, B.; Roger, J.M.; McBratney, A. Critical review of chemometric indicators commonly used for assessing the quality of the prediction of soil attributes by NIR spectroscopy. Trends Analyt. Chem. 2010, 29, 1073–1081. [Google Scholar] [CrossRef]
- Luedeling, E.; Zhang, M.; Luedeling, V.; Girvetz, E.H. Sensitivity of winter chill models for fruit and nut trees to climatic changes expected in California’s Central Valley. Agric. Ecosyst. Envir. 2009, 133, 23–31. [Google Scholar] [CrossRef]
- Guo, L.; Dai, J.; Ranjitkar, S.; Xu, J.; Luedeling, E. Response of chestnut phenology in China to climate variation and change. Agric. For. Meteorol. 2013, 180, 164–172. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Hadley, P.; Ordidge, M.; Xu, X.; Luedeling, E. Delayed chilling appears to counteract flowering advances of apricot in southern UK. Agric. For. Meteorol. 2017, 237–238, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Luedeling, E.; Kunz, A.; Blanke, M. Identification of chilling and heat requirements of cherry trees—A statistical approach. Intern. J. Biometeorol. 2013, 57, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Maulión, E.; Valentini, G.H.; Kovalevski, L.; Prunello, M.; Monti, L.L.; Daorden, M.E.; Quaglino, M.; Cervigni, G.D.L. Comparison of methods for estimation of chilling and heat requirements of nectarine and peach genotypes for flowering. Sci. Hortic. 2014, 177, 112–117. [Google Scholar] [CrossRef]
- Luedeling, E.; Guo, L.; Dai, J.; Leslie, C.; Blanke, M.M. Differential responses of trees to temperature variation during the chilling and forcing phases. Agric. For. Meteorol. 2013, 181, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Sawamura, Y.; Suesada, Y.; Sugiura, T.; Yaegaki, H. Chilling requirements and blooming dates of leading peach cultivars and a promising early maturing peach selection; Momo Tsukuba 127. Hortic. J. 2014, 86, 426–436. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Dai, J.H.; Ranjitkar, S.; Yu, H.Y.; Xu, J.C.; Luedeling, E. Chilling and heat requirements for flowering in temperate fruit trees. Int. J. Biometeorol. 2014, 58, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Pope, K.S.; Da Silva, D.; Brown, P.H.; DeJong, T.M. A biologically based approach to modeling spring phenology in temperate deciduous trees. Agric. For. Meteorol. 2014, 198–199, 15–23. [Google Scholar] [CrossRef]
- Luedeling, E.; Schiffers, K.; Fohrmann, T.; Urbach, C. PhenoFlex—An integrated model to predict spring phenology in temperate fruit trees. Agric. For. Meteorol. 2021, 307, 108491. [Google Scholar] [CrossRef]
- Atagul, O.; Calle, A.; Demirel, G.; Lawton, J.M.; Bridges, W.C.; Gasic, K. Estimating heat requirement for flowering in peach germplasm. Agronomy 2022, 12, 1002. [Google Scholar] [CrossRef]
- Bielenberg, D.G.; Gasic, K. Peach [Prunus persica (L.) Batsch] Cultivars differ in apparent base temperature and growing degree hour requirement for floral bud break. Front. Plant Sci. 2022, 13, 801606. [Google Scholar] [CrossRef]
- Gilreath, P.R.; Buchanan, D.W. Floral and vegetative bud development of ‘Sungold’ and ‘Sunlite’ nectarine as influenced by evaporative cooling by overhead sprinkling during rest. J. Am. Soc. Hort. Sci. 1981, 106, 321–332. [Google Scholar] [CrossRef]
- Fernández, E.; Luedeling, E.; Behrend, D.; Van de Vliet, S.; Kunz, A.; Fadón, E. Mild Water Stress Makes Apple Buds More Likely to Flower and More Responsive to Artificial Forcing—Impacts of an unusually warm and dry summer in Germany. Agronomy 2020, 10, 274. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fruit Type | Flesh Colour | Flesh Texture | Fruit Taste | Ripening Period | Breeding Programme | |
|---|---|---|---|---|---|---|
| Carlacov | Peach | Yellow | Melting | Sweet | Early | PSB Producción Vegetal, Spain |
| Catherina® | Peach | Yellow | Non-melting | Equilibrate | Medium | L. Hough, USA |
| O’Henry | Peach | Yellow | Melting | Equilibrate | Late | G. Merrill, USA |
| Sweet Dreamcov | Peach | Yellow | Melting | Very sweet | Medium | Zaiger Genetics, USA |
| Elegant Lady | Peach | Yellow | Melting | Sour | Medium | G. Merrill, USA |
| Patty® | Peach | White | Melting | Sour | Early | Zaiger Genetics, USA |
| Sweetregalcov | Peach | White | Melting | Very sweet | Medium | A. & L. Maillard, France |
| Gladys® | Peach | White | Melting | Sour | Late | Zaiger Genetics, USA |
| Sweet Cap® | Flat Peach | White | Melting | Very sweet | Medium | A. & L. Maillard, France |
| Flatstarcov | Flat Peach | White | Melting | Sweet | Medium | ASF, France |
| BigTop® | Nectarine | Yellow | Melting | Sweet | Medium | Zaiger Genetics, USA |
| Venus | Nectarine | Yellow | Melting | Sour | Medium | CREA, Italy |
| Nectarperfcov | Nectarine | White | Melting | Sweet | Late | A. & L. Maillard, France |
| Emeraude® | Nectarine | White | Melting | Sweet | Medium | R. Monteux-Caillet, France |
| Country | Location | Latitude | Longitude | Tree Density | Training System |
|---|---|---|---|---|---|
| Romania | Bucharest | 44.4 | 26.1 | 1667 | Vertical axe |
| Italy | Cuneo | 44.4 | 7.5 | 966 | Bi-axis * |
| Italy | Forlì | 44.2 | 12.0 | 741 | Open vase |
| Italy | Tebano | 44.1 | 12.2 | 635 | Open vase |
| France | Bellegarde | 43.8 | 4.5 | 556 | Double Y |
| Italy | Rome | 41.9 | 12.5 | 741 | Open vase |
| Spain | Zaragoza | 41.7 | −0.9 | 571 | Open vase |
| Spain | Lleida | 41.6 | 0.6 | 667 | Catalan vase |
| Greece | Naoussa | 40.6 | 22.1 | 571 | Open vase |
| Spain | Murcia−Yéchar | 38.1 | −1.4 | 800 | Open vase |
| Spain | Murcia−Torre Pacheco | 37.8 | −1.0 | 850 | Open vase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drogoudi, P.; Cantín, C.M.; Brandi, F.; Butcaru, A.; Cos-Terrer, J.; Cutuli, M.; Foschi, S.; Galindo, A.; García-Brunton, J.; Luedeling, E.; et al. Impact of Chill and Heat Exposures under Diverse Climatic Conditions on Peach and Nectarine Flowering Phenology. Plants 2023, 12, 584. https://doi.org/10.3390/plants12030584
Drogoudi P, Cantín CM, Brandi F, Butcaru A, Cos-Terrer J, Cutuli M, Foschi S, Galindo A, García-Brunton J, Luedeling E, et al. Impact of Chill and Heat Exposures under Diverse Climatic Conditions on Peach and Nectarine Flowering Phenology. Plants. 2023; 12(3):584. https://doi.org/10.3390/plants12030584
Chicago/Turabian StyleDrogoudi, Pavlina, Celia M. Cantín, Federica Brandi, Ana Butcaru, José Cos-Terrer, Marcello Cutuli, Stefano Foschi, Alejandro Galindo, Jesus García-Brunton, Eike Luedeling, and et al. 2023. "Impact of Chill and Heat Exposures under Diverse Climatic Conditions on Peach and Nectarine Flowering Phenology" Plants 12, no. 3: 584. https://doi.org/10.3390/plants12030584
APA StyleDrogoudi, P., Cantín, C. M., Brandi, F., Butcaru, A., Cos-Terrer, J., Cutuli, M., Foschi, S., Galindo, A., García-Brunton, J., Luedeling, E., Moreno, M. A., Nari, D., Pantelidis, G., Reig, G., Roera, V., Ruesch, J., Stanica, F., & Giovannini, D. (2023). Impact of Chill and Heat Exposures under Diverse Climatic Conditions on Peach and Nectarine Flowering Phenology. Plants, 12(3), 584. https://doi.org/10.3390/plants12030584