

Defoliation in Perennial Plants: Predictable and Surprising Results in Senna spp.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- (1)
- Will plants experiencing greater levels of defoliation grow less than those with less damage?
- (2)
- Will plants experiencing greater levels of defoliation produce fewer new leaves?
- (3)
- Will plants experiencing greater levels of defoliation produce more foliar nectaries?
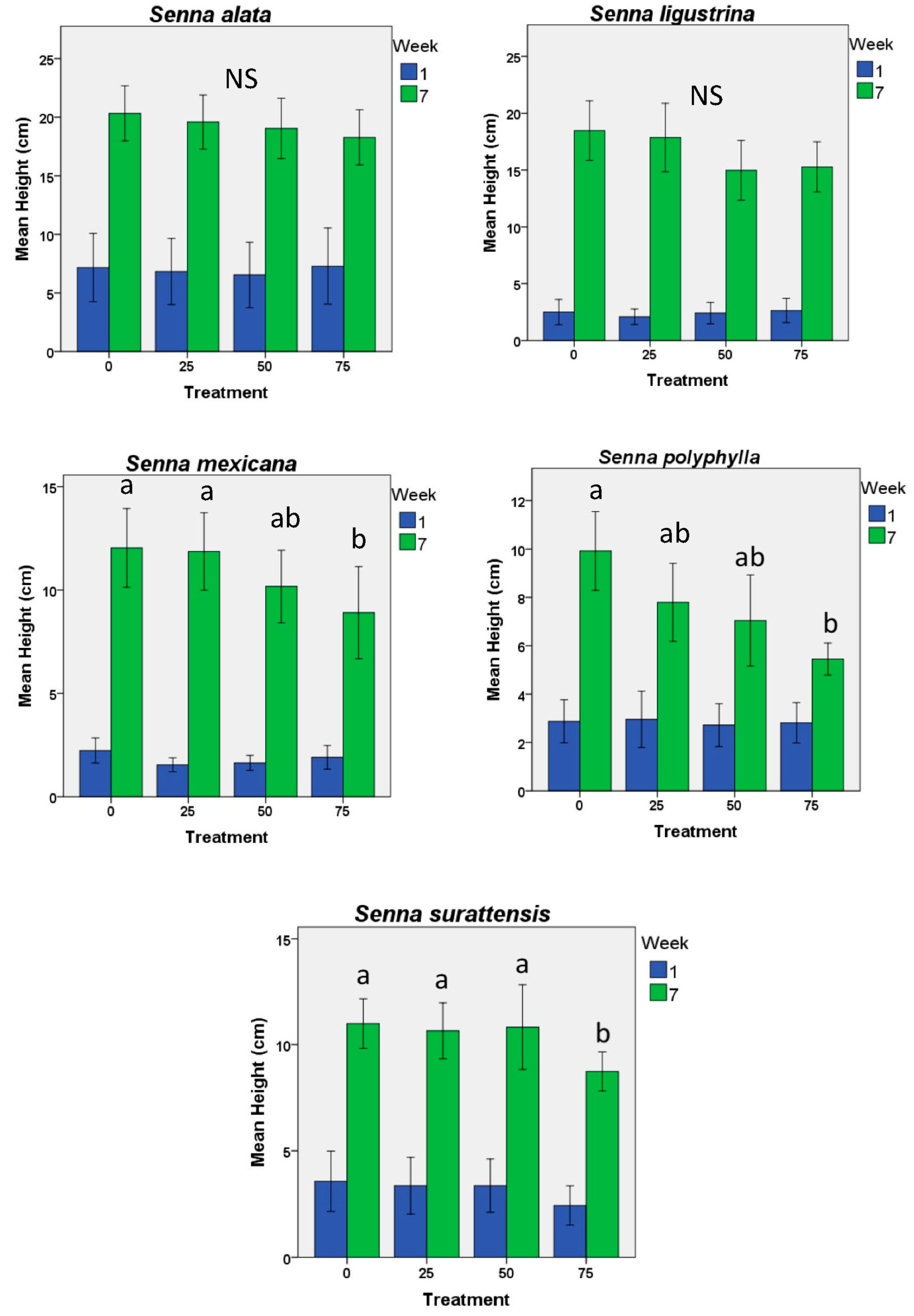
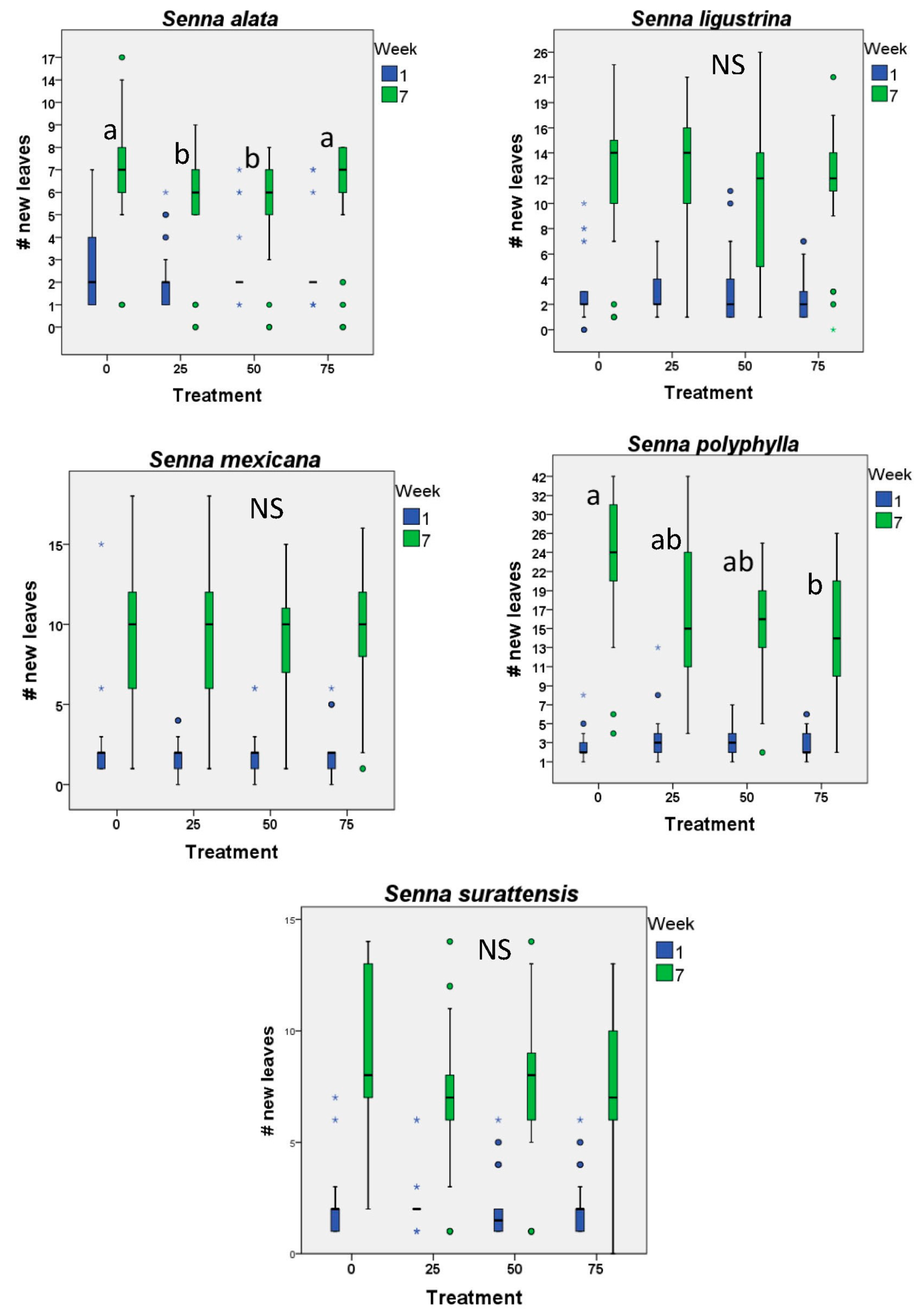
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Species
4.2. Experimental Design
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crawley, M.J. Herbivory. The Dynamics of Animal-Plant Interactions; University of California Press: Berkeley, CA, USA, 1983; 437p. [Google Scholar]
- Schupp, E.W. Azteca protection of Cecropia: Ant occupation benefits juvenile trees. Oecol 1986, 70, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Rockwood, L.L. The effect of defoliation on seed production in six Costa Rican tree species. Ecology 1973, 54, 1363–1369. [Google Scholar] [CrossRef]
- Marshall, D.L.; Abrahamson, N.J.; Avritt, J.J.; Hall, P.M.; Medeiros, J.S.; Reynolds, J.; Shaner, M.G.M.; Simpson, H.L.; Trafton, A.N.; Tyler, A.P.; et al. Differences in Plastic Responses to Defoliation due to Variation in the Timing of Treatments for Two Species of Sesbania (Fabaceae). Ann. Bot. 2005, 95, 1049–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, S.; Tieszen, L.L. Effects of simulated grazing on foliage and root production and biomass allocation in an arctic tundra sedge (Eriophorum vaginatum). Oecologia 1983, 58, 92–102. [Google Scholar] [CrossRef]
- Rockwood, L.L.; Lobstein, M.B. The effects of experimental defoliation on reproduction in four species of herbaceous perennials from northern Virginia. Castanea 1994, 59, 41–50. [Google Scholar]
- Reece, P.W.; Morris, J.W.; Schacht, W.H.; Koehler, A.E.; Volesky, J.D.; Moser, L.E. Prairie sandreed response to preceding-year defoliation and precipitation regime. Great Plains Res. 2007, 17, 215–224. [Google Scholar]
- McNaughton, S.J. Compensatory plant growth as a response to herbivory. Oikos 1983, 40, 329–336. [Google Scholar] [CrossRef]
- Klubertanz, T.H.; Pedigo, L.P.; Carlson, R.E. Soybean Physiology, Regrowth, and Senescence in Response to Defoliation. Agron. J. 1996, 88, 577–582. [Google Scholar] [CrossRef]
- Barton, K.E.; Shiels, A.B. Additive and non-additive responses of seedlings to simulated herbivory and drought. Biotropica 2020, 52, 1217. [Google Scholar] [CrossRef]
- Erb, M.; Lenk, C.; Degenhardt, J.; Turlings, T.C.J. The underestimated role of roots in defense against leaf attackers. Trends Plant Sci. 2009, 14, 653–659. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.D. Buried but unsafe—Defoliation depletes the underground storage organ (USO) of the mesic grassland geophyte, Hypoxis hemerocallidea. S. Afr. J. Bot. 2021, 141, 265–272. [Google Scholar] [CrossRef]
- Hinman, E.D.; Fridley, J.D. Impacts of experimental defoliation on native and invasive saplings: Are native species more resilient to canopy disturbance? Tree Physiol. 2020, 40, 969–979. [Google Scholar] [CrossRef]
- Meuriot, F.; Prud’homme, M.; Noiraud-Romy, N. Defoliation, wounding, and methyl jasmonate induce expression of the sucrose lateral transporter LpSUT1 in ryegrass (Lolium perenne L.). Physiol. Plantarum. 2022, 174, e13744. [Google Scholar] [CrossRef]
- Agrawal, A.A. Overcompensation of plants in response to herbivory and the by-product benefits of mutualism. Trends Plant Sci. 2000, 5, 309–313. [Google Scholar] [CrossRef]
- Fleet, R.R.; Young, B.L. Facultative Mutualism between Imported Fire Ants (Solenopsis invicta) and a Legume (Senna occidentalis). Southwest. Nat. 2000, 45, 289–298. [Google Scholar] [CrossRef]
- Koptur, S.; Jones, I.M.; Pena, J.E. The Influence of Host Plant Extrafloral Nectaries on Multitrophic Interactions: An Experimental Investigation. PLoS ONE 2015, 10, e0138157. [Google Scholar] [CrossRef]
- Koptur, S.; Jones, I.M.; Liu, H.; Diaz-Castelazo, C. Playing the System: The Impacts of Invasive Ants and Plants on Facultative Ant-Plant Interactions. In Ant-Plant Interactions—Impacts of Humans on Terrestrial Ecosystems; Oliveira, P.S., Koptur, S., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 249–266. [Google Scholar]
- Koptur, S. Extrafloral nectary-mediated interactions between insects and plants. In Insect-Plant Interactions; Bernays, E., Ed.; CRC Press: Boca Raton, FL, USA, 1992; Volume 4, pp. 81–129. [Google Scholar]
- Marazzi, B.; Bronstein, J.L.; Koptur, S. The diversity, ecology and evolution of extrafloral nectaries: Current perspectives and future challenges. Ann. Bot. 2013, 111, 1243–1250. [Google Scholar] [CrossRef] [Green Version]
- Bentley, B. Extrafloral nectaries and protection by pugnacious bodyguards. Annu. Rev. Ecol. Syst. 1977, 88, 407–427. [Google Scholar] [CrossRef]
- Barton, A.M. Spatial variation in the effect of ants on an extrafloral nectary plant. Ecology 1986, 67, 495–504. [Google Scholar] [CrossRef]
- Kelly, C.A. Extrafloral nectaries: Ants, herbivores and fecundity in Cassia fasciculata. Oecol 1986, 69, 600–605. [Google Scholar] [CrossRef]
- Koptur, S. Nectar as fuel for plant protectors. In Plant-Provided Food for Carnivorous Insects: Protective Mutualism and Its Applications; Wackers, F.L., VanRijn, P.C.J., Bruin, J., Eds.; Cambridge University Press: New York, NY, USA, 2005; pp. 75–108. [Google Scholar]
- Boecklen, W.J. The role of extrafloral nectaries in the herbivore defence of Cassia fasiculata. Ecol. Entomol. 1984, 9, 243–249. [Google Scholar] [CrossRef]
- Lange, D.; Calixto, E.S.; Del-Claro, K. Variation in Extrafloral Nectary Productivity Influences the Ant Foraging. PLoS ONE 2017, 12, e0169492. [Google Scholar] [CrossRef] [PubMed]
- Jones, I.M.; Koptur, S. Dynamic Extrafloral Nectar Production: The Timing of Leaf Damage Affects the Defensive Response in Senna mexicana var. chapmanii (Fabaceae). Am. J. Bot. 2015, 102, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koptur, S. Is extrafloral nectar production an inducible defense? In Evolutionary Ecology of Plants; Bock, J., Linhart, Y., Eds.; Westview Press: Boulder, CO, USA, 1989; pp. 323–339. [Google Scholar]
- Rogers, W.E.; Sieman, E.; Lankau, R.A. Damage Induced Production of Extrafloral Nectaries in Native and Invasive Seedlings of Chinese Tallow Tree (Sapium sebiferum). Am. Midl. Nat. 2003, 149, 413–417. [Google Scholar] [CrossRef]
- Heil, M. Induction of two indirect defences benefits Lima bean (Phaseolus lunatus, Fabaceae) in nature. J. Ecol. 2004, 92, 527–536. [Google Scholar] [CrossRef]
- Heil, M. Indirect defence via tritrophic interactions. New Phytol. 2008, 178, 41–61. [Google Scholar] [CrossRef]
- Hernandez-Cumplido, J.; Forter, B.; Moreira, X.; Heil, M.; Benrey, B. Induced Floral and Extrafloral Nectar Production Affect Ant-pollinator Interactions and Plant Fitness. Biotropica 2016, 48, 342–348. [Google Scholar] [CrossRef]
- Yamawo, A.; Suzuki, N. Induction and relaxation of extrafloral nectaries in response to simulated herbivory in young Mallotus japonicus plants. J. Plant Res. 2018, 131, 255–260. [Google Scholar] [CrossRef]
- Chinarelli, H.D.; Nogueira, A.; Leal, L.C. Extrafloral nectar production induced by simulated herbivory does not improve ant bodyguard attendance and ultimately plant defence. Biol. J. Linn. Soc. 2021, 135, 429–446. [Google Scholar] [CrossRef]
- Jones, I.M.; Koptur, S.; Gallegos, H.R.; Tardanico, J.P.; Trainer, P.A.; Peña, J. Changing light conditions in pine rockland habitats affect the intensity and outcome of ant-plant interactions. Biotropica 2017, 49, 83–91. [Google Scholar] [CrossRef]
- Kersch, M.F.; Fonseca, C.R. Abiotic factors and the conditional outcome of an ant-plant mutualism. Ecology 2005, 86, 2117–2126. [Google Scholar] [CrossRef] [Green Version]
- Jones, I.M.; Koptur, S.; Pena, J.E. Exploring whether and how ants (Hymenoptera: Formicidae) affect reproductive fitness in Senna mexicana var. chapmanii (Fabaceae). Fla. Entomol. 2017, 100, 539–545. [Google Scholar] [CrossRef]
- Mondor, E.B.; Addicott, J.F. Conspicuous extra-floral nectaries are inducible in Vicia faba. Ecol. Lett. 2003, 6, 495–497. [Google Scholar] [CrossRef]
- Mondor, E.B.; Tremblay, M.N.; Messing, R.H. Extrafloral nectary phenotypic plasticity is damage- and resource-dependent in Vicia faba. Biol. Lett. 2006, 2, 583–585. [Google Scholar] [CrossRef] [Green Version]
- Pulice, C.E.; Packer, A.A. Simulated herbivory induces extrafloral nectary production in Prunus avium. Funct. Ecol. 2008, 22, 801–807. [Google Scholar] [CrossRef]
- Yamawo, A.; Suzuki, N.; Tagawa, J. Extrafloral nectary-bearing plant Mallotus japonicus uses different types of extrafloral nectaries to establish effective defense by ants. J. Plant Res. 2019, 132, 499–507. [Google Scholar] [CrossRef]
- Wirf, L.A. The effect of manual defoliation and Macaria pallidata (Geometridae) herbivory on Mimosa pigra: Implications for biological control. Biol. Control 2006, 37, 346–353. [Google Scholar] [CrossRef]
- Mondor, E.B.; Keiser, C.N.; Pendarvis, D.E.; Vaughn, M.N. Broad bean cultivars increase extrafloral nectary numbers, but not extrafloral nectar, in response to leaf damage. Open J. Ecol. 2013, 3, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Calixto, E.S.; Lange, D.; Bronstein, J.; Torezan-Silingardi, H.M.; Del-Claro, K. Optimal Defense Theory in an ant–plant mutualism: Extrafloral nectar as an induced defence is maximized in the most valuable plant structures. J. Ecol. 2021, 109, 167–178. [Google Scholar] [CrossRef]
- Calixto, E.S.; Lange, D.; Moreira, X.; Del-Claro, K. Plant species specificity of ant–plant mutualistic interactions: Differential predation of termites by Camponotus crassus on five species of extrafloral nectaries plants. Biotropica 2021, 53, 1406–1414. [Google Scholar] [CrossRef]
- Calixto, E.S.; Lange, D.; Del-Claro, K. Net benefits of a mutualism: Influence of the quality of extrafloral nectar on the colony fitness of a mutualistic ant. Biotropica 2021, 53, 846–856. [Google Scholar] [CrossRef]
- Weltzin, J.F.; Archer, S.R.; Heitschmidt, R.K. Defoliation and woody plant (Prosopis glandulosa) seedling regeneration: Potential vs realized herbivory tolerance. Plant Ecol. 1998, 138, 127–135. [Google Scholar] [CrossRef]
- Agrawal, A.A. A scale-dependent framework for trade-offs, syndromes, and specialization in organismal biology. Ecology 2020, 101, e02924. [Google Scholar] [CrossRef] [PubMed]
- Archer, S.; Detling, J.K. The Effects of Defoliation and Competition on Regrowth of Tillers of Two North American Mixed-Grass Prairie Graminoids. Oikos 1984, 43, 351–357. [Google Scholar] [CrossRef]
- Quentin, A.G.; Pinkard, E.A.; Beadle, C.L.; Wardlaw, T.J.; O’Grady, A.P.; Paterson, S.; Mohammed, C.L. Do artificial and natural defoliation have similar effects on physiology of Eucalyptus globulus Labill. seedlings? Ann. For. Sci. 2010, 67, 203. [Google Scholar] [CrossRef] [Green Version]
- Rakotonoely, H.; Ramamonjisoa, N. Seedlings of the Invasive Strawberry Guava Psidium cattleianum Were More Sensitive to Defoliation Than the Closely Related Malagasy Native Eugenia goviala in a Simulated Herbivory Experiment. Trop. Conserv. Sci. 2020, 13, 1940082920961230. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Kotanen, P.M. Herbivores and the success of exotic plants: A phylogenetically controlled experiment. Ecol. Lett. 2003, 6, 712–715. [Google Scholar] [CrossRef] [Green Version]
- Veeraragavan, S.; Ramamoorthy, D. Studies On Micro-Morphological Characteristics Of Extra Floral Nectar -Secreting Glandular Structures In Cassia alata Linn. Int. J. Recent Sci. Res. 2015, 6, 5759–5763. [Google Scholar]
- Veeraragavan, S.; Ramamoorthy, D. SEM based anatomical studies in stipular extra floral nectary tissues in Cassia alata Linn. Int. J. Curr. Res. 2016, 8, 25955–25960. [Google Scholar]
- Marazzi, B.; Conti, E.; Sanderson, M.J.; McMahon, M.M.; Bronstein, J.L. Diversity and evolution of a trait mediating ant-plant interactions: Insights from extrafloral nectaries in Senna (Leguminosae). Ann. Bot. 2013, 111, 1263–1275. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koptur, S.; Primoli, A.S.; Pimienta, M.C. Defoliation in Perennial Plants: Predictable and Surprising Results in Senna spp. Plants 2023, 12, 587. https://doi.org/10.3390/plants12030587
Koptur S, Primoli AS, Pimienta MC. Defoliation in Perennial Plants: Predictable and Surprising Results in Senna spp. Plants. 2023; 12(3):587. https://doi.org/10.3390/plants12030587
Chicago/Turabian StyleKoptur, Suzanne, Andrea Salas Primoli, and María Cleopatra Pimienta. 2023. "Defoliation in Perennial Plants: Predictable and Surprising Results in Senna spp." Plants 12, no. 3: 587. https://doi.org/10.3390/plants12030587