

Phytotoxicity of Quillaja lancifolia Leaf Saponins and Their Bioherbicide Potential

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Phytotoxicity Assay
2.1.1. Osmotic Potential, pH of Extracts, and Presence of Saponins
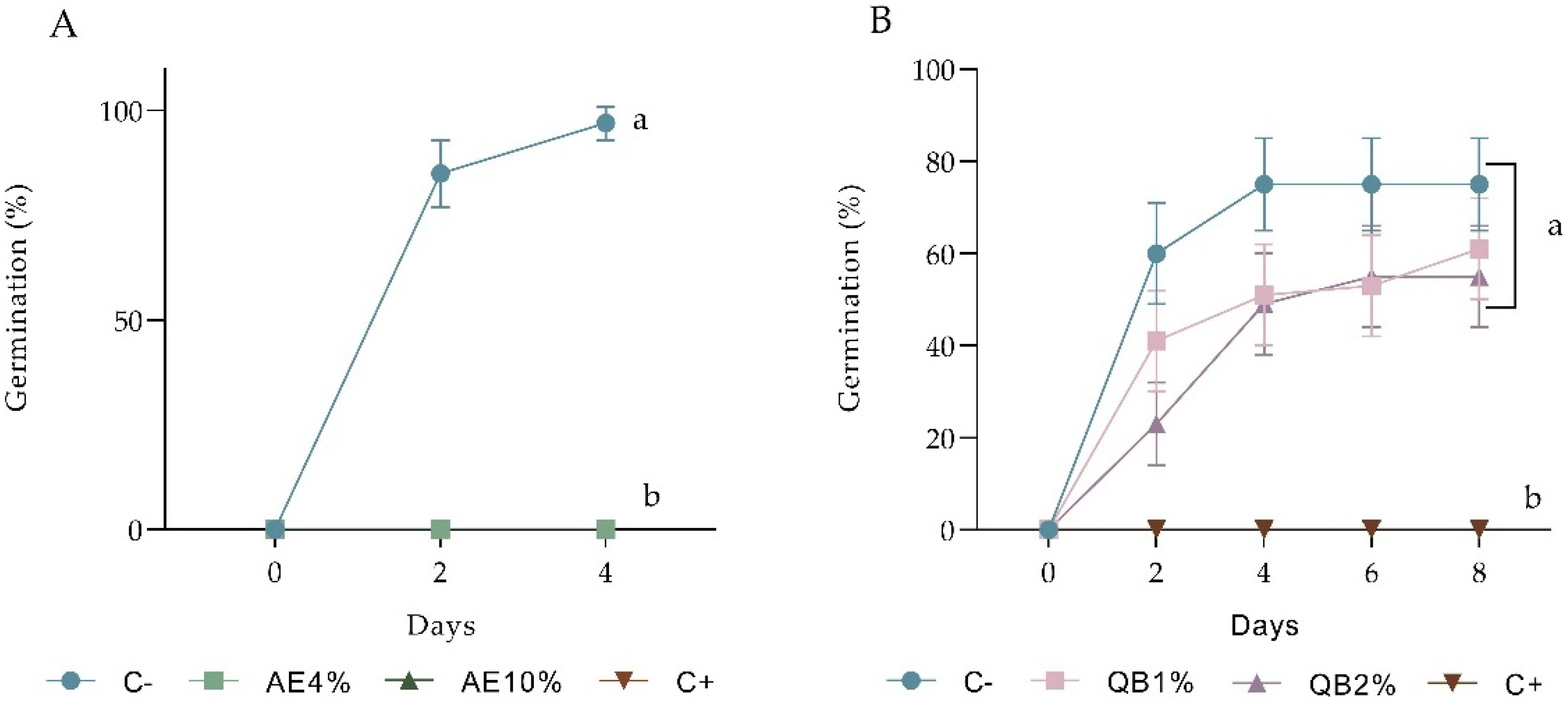
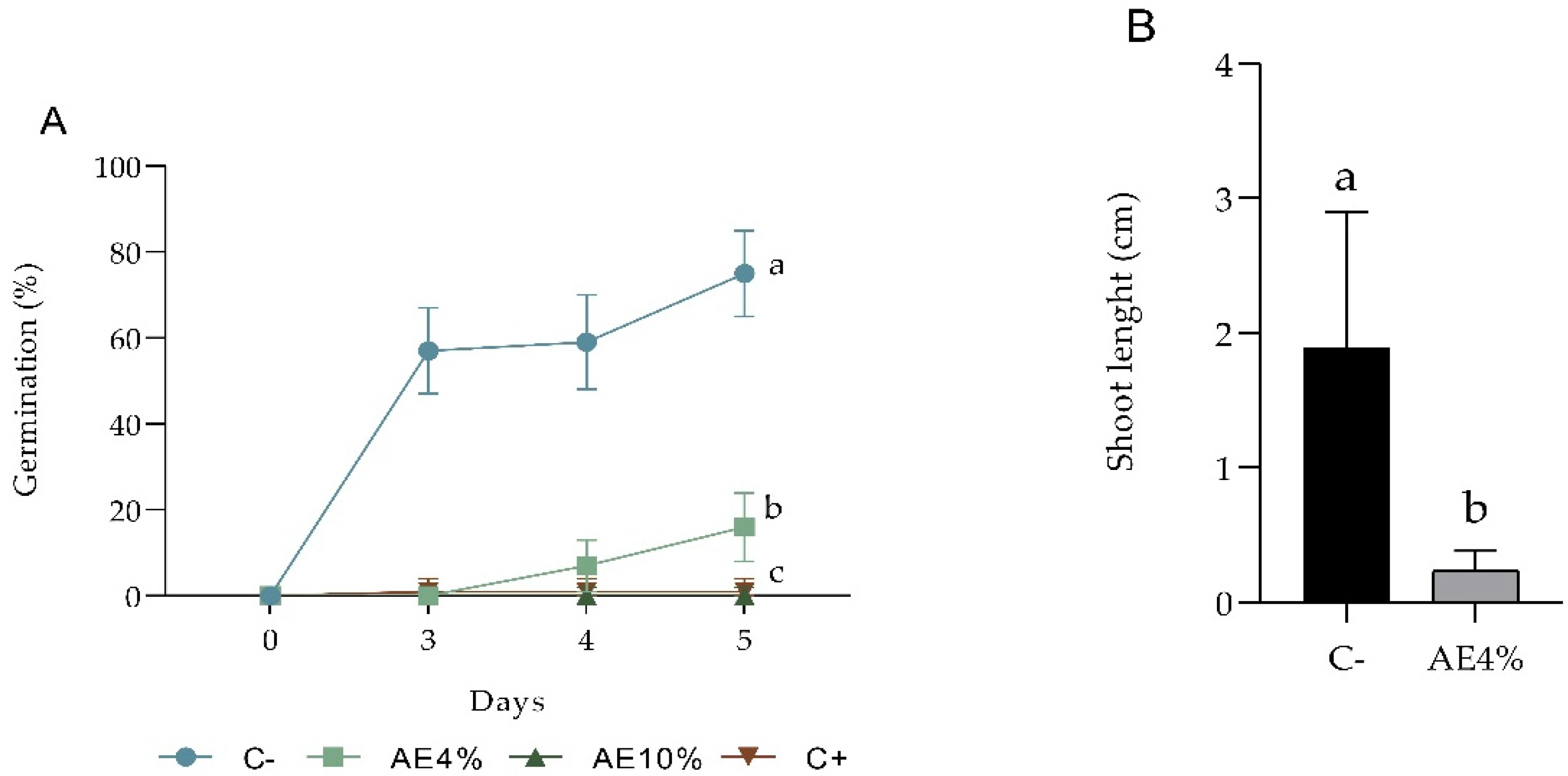
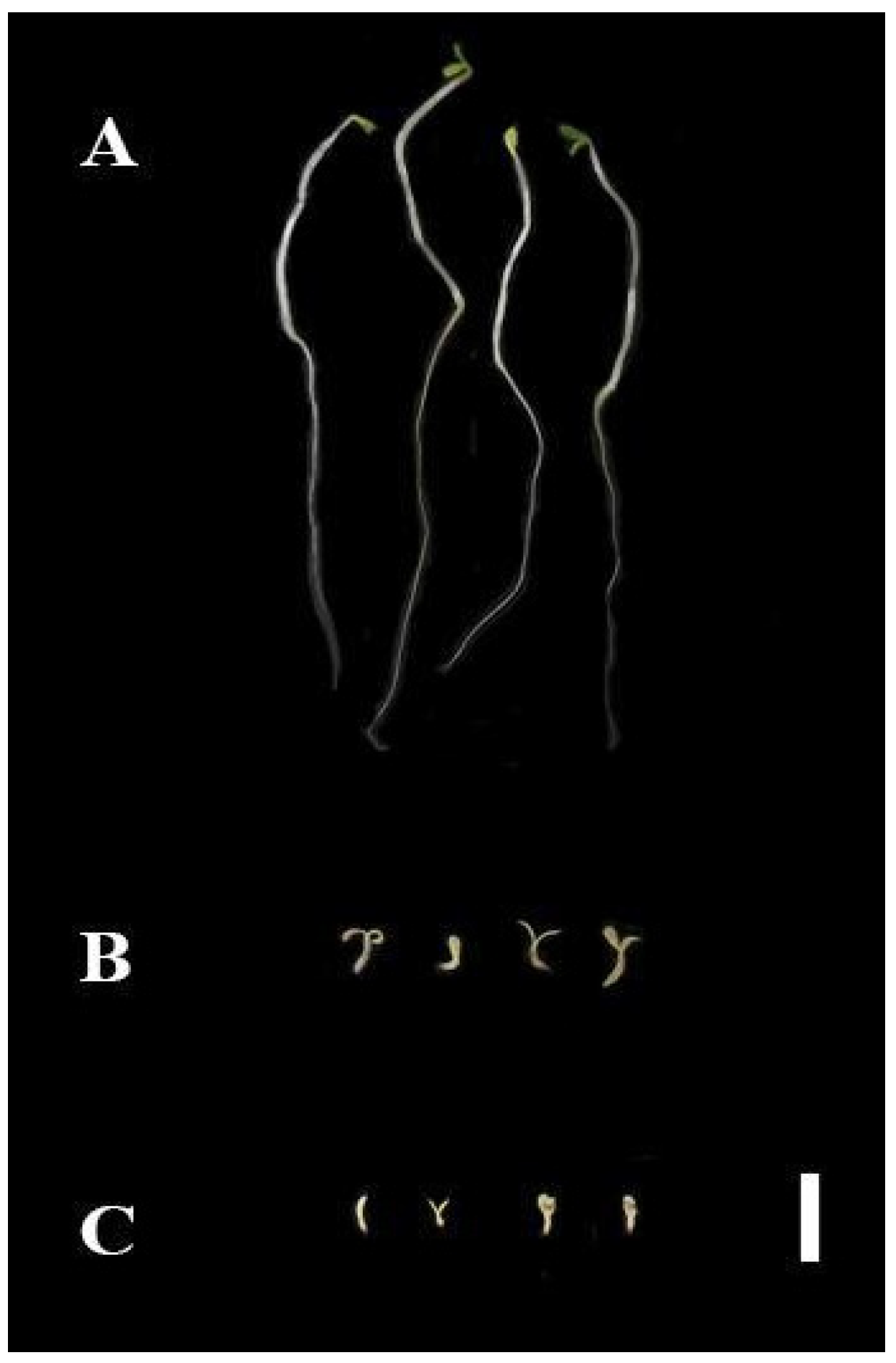
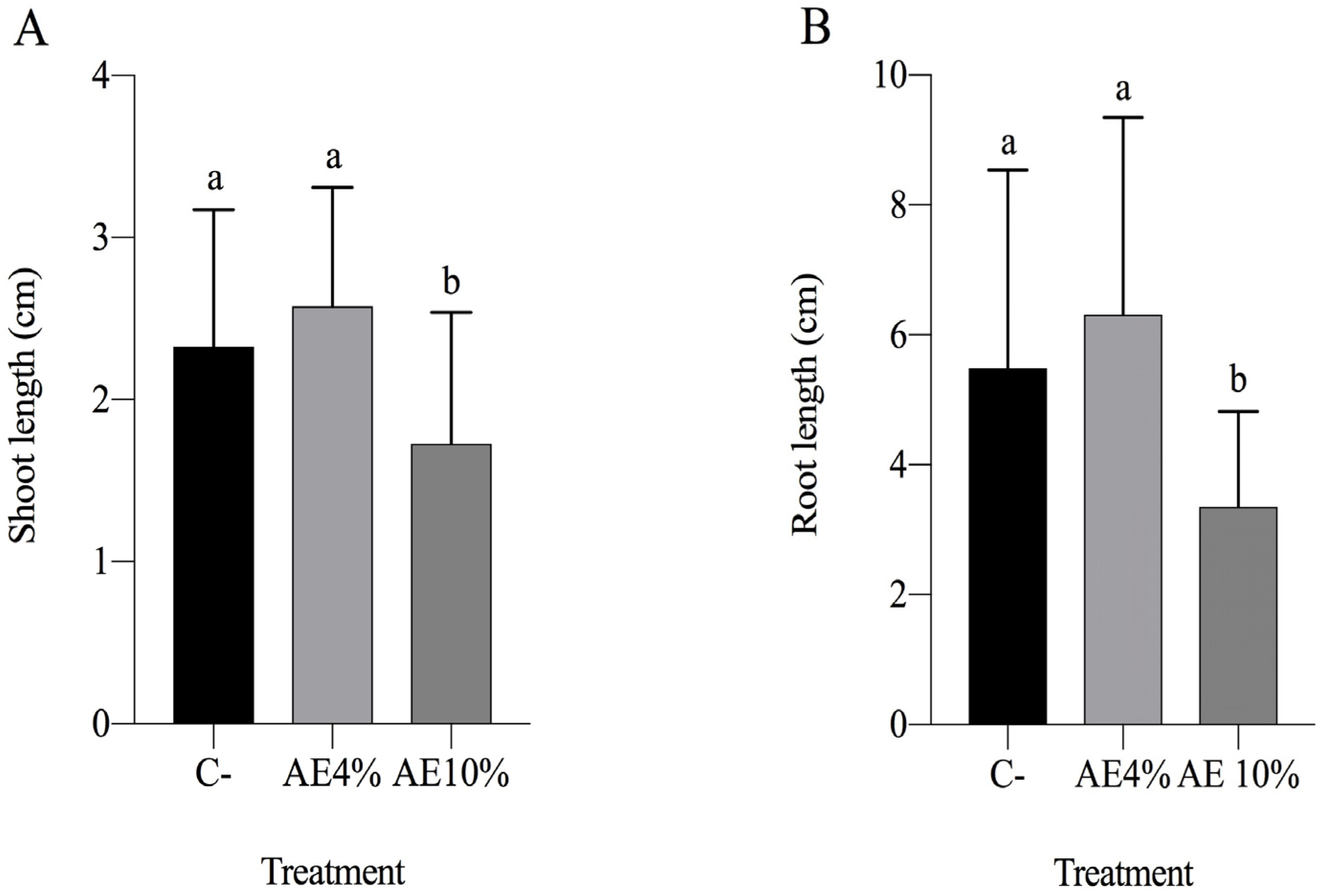
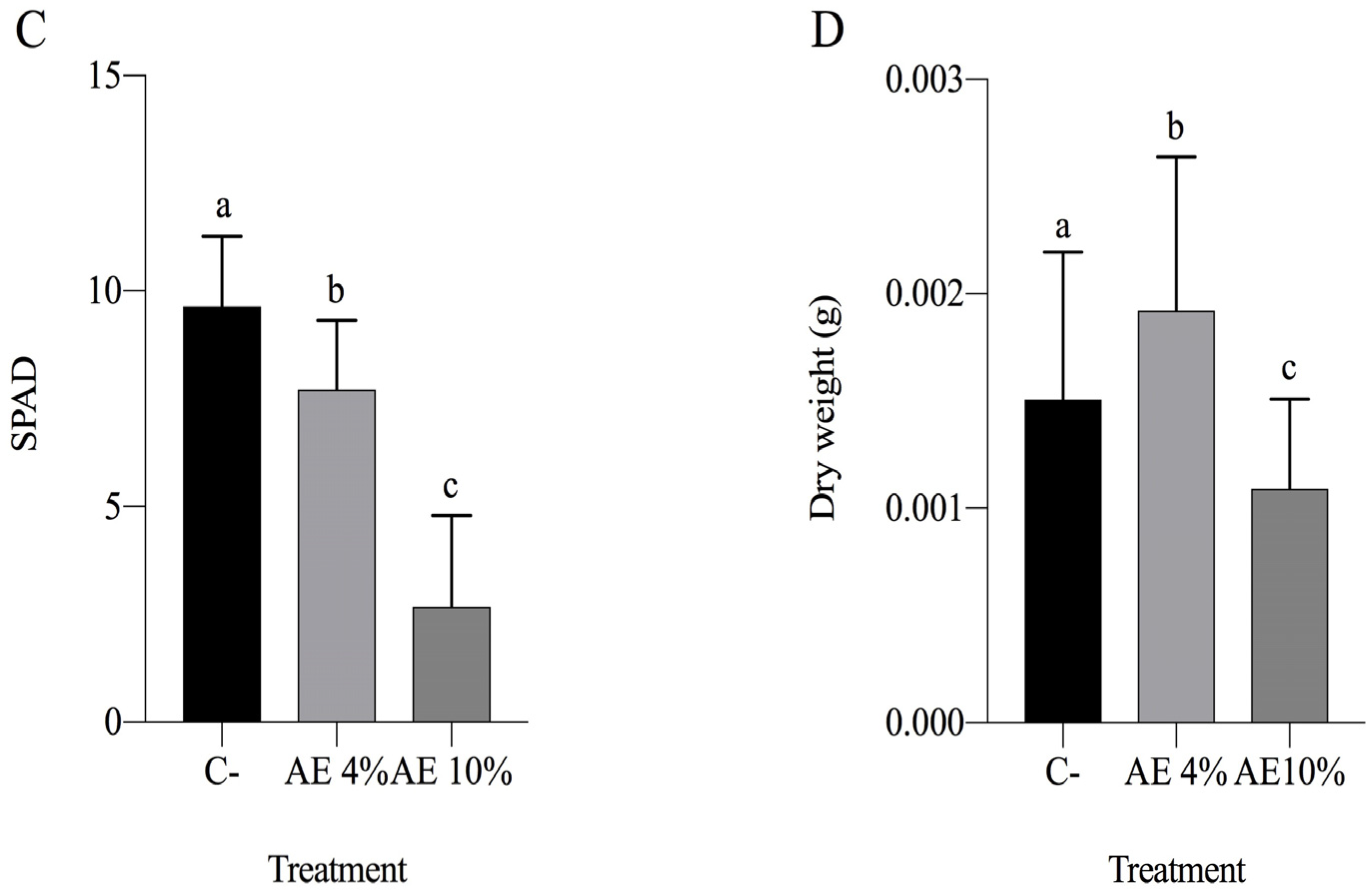
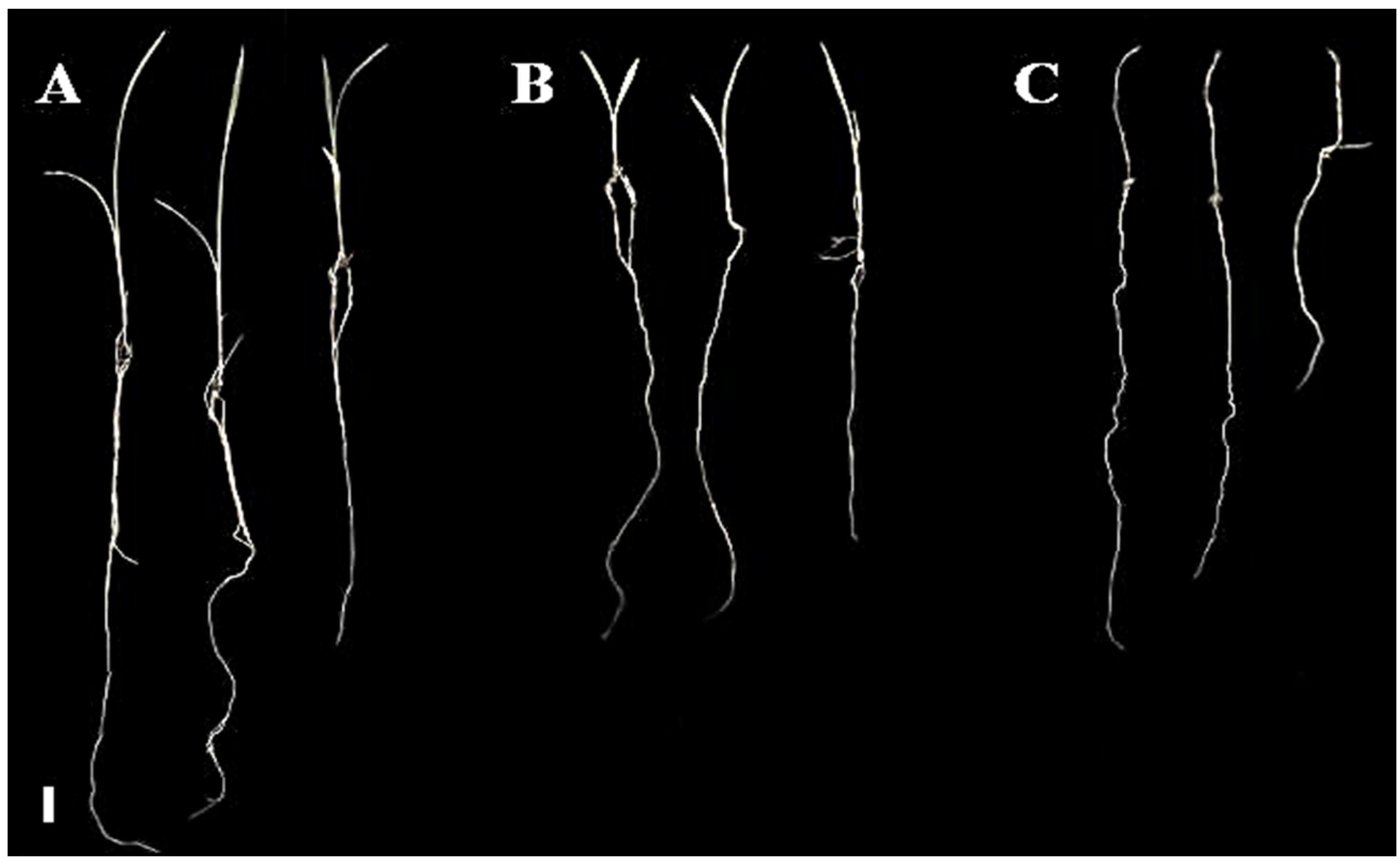
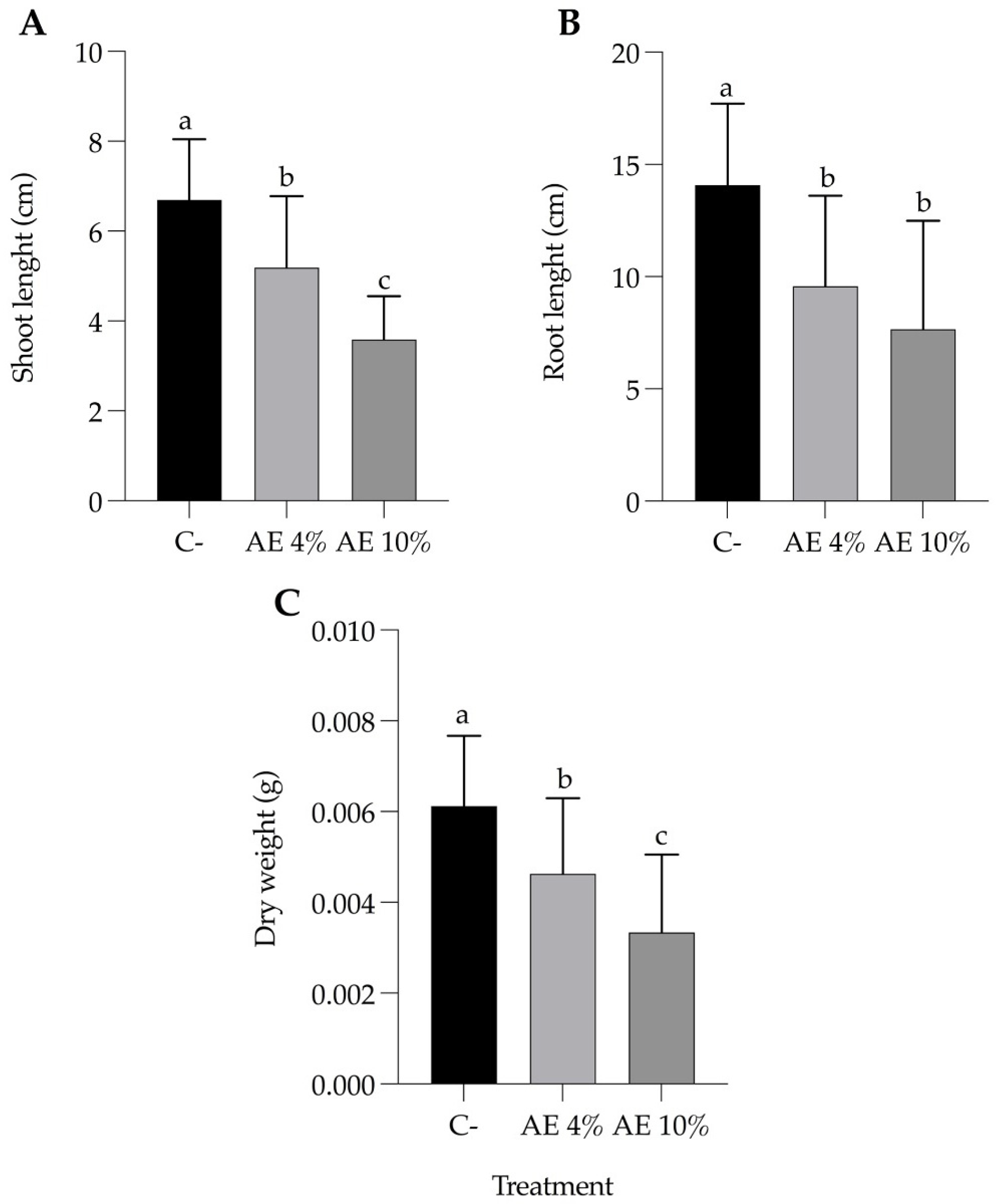
2.1.2. Pre-Emergence Bioassay
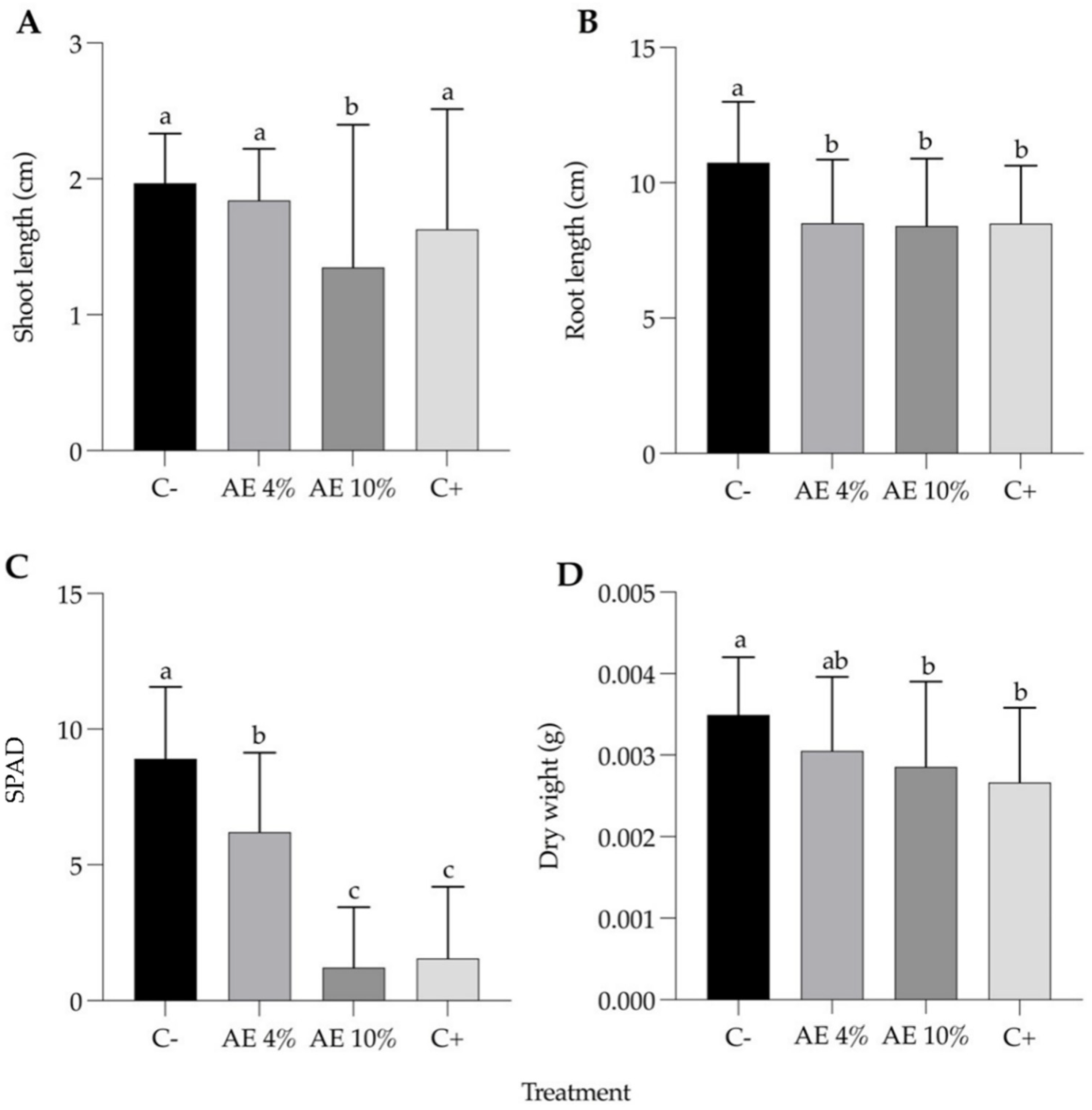
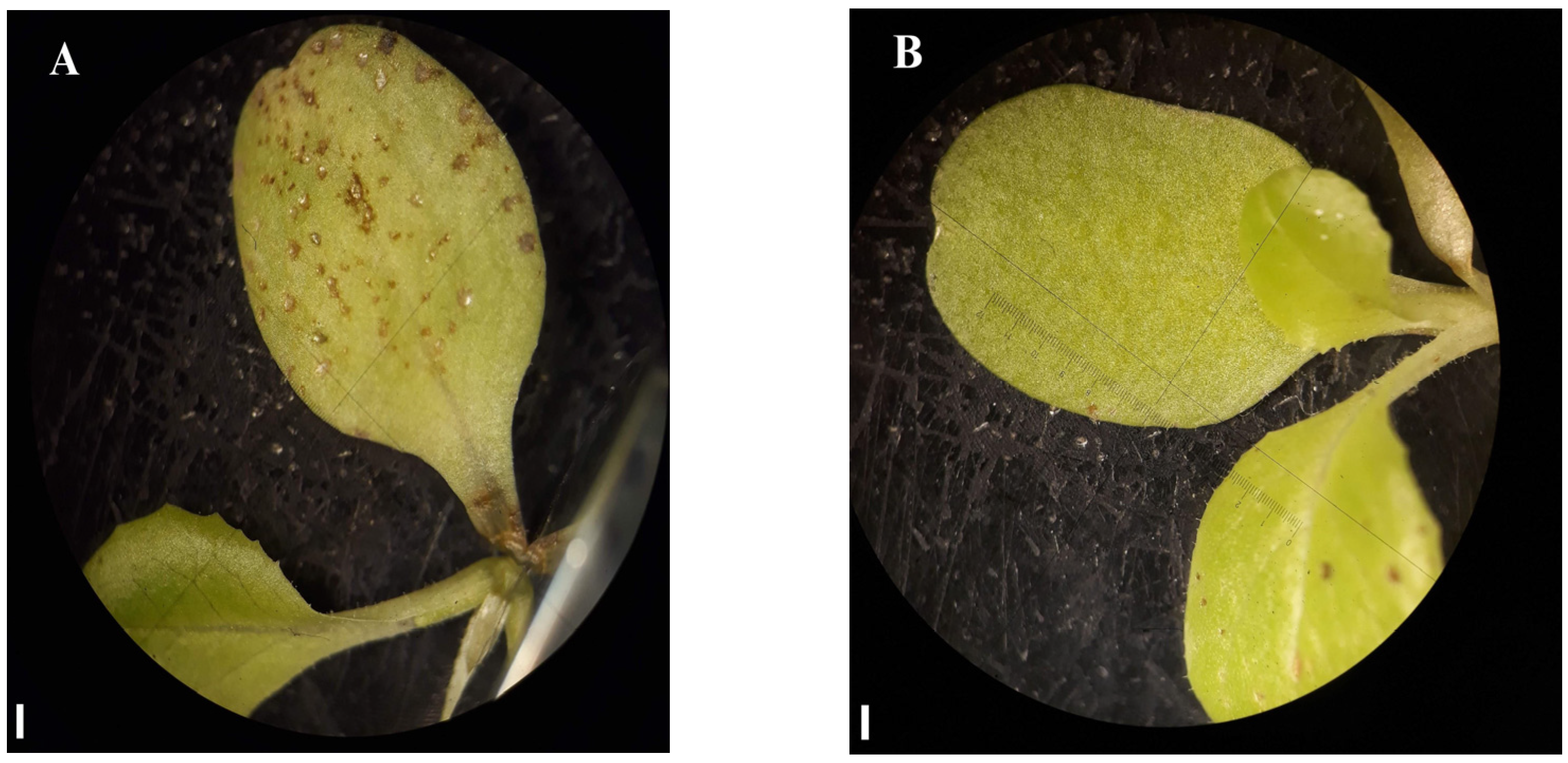
2.1.3. Post-Emergence Bioassay
2.1.4. Substrate Leaching Bioassay
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material and Extracts
5.1.1. Plant Source and Extract Preparation
5.1.2. Extract and Saponin Chemical Characterization
5.2. Phytotoxicity Assay
5.2.1. Target Species
5.2.2. Extract Osmotic Potential and pH Measurements
5.2.3. Pre-Emergence Bioassay
5.2.4. Bioassay for Post-Emergence
5.2.5. Substrate Leaching Bioassay
5.2.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rehman, A.; Farooq, M.; Lee, D.J.; Siddique, K.H.M. Sustainable agricultural practices for food security and ecosystem services. Environ. Sci. Pollut. Res. 2022, 29, 84076–84095. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations. FAOSTAT-Pesticides Use. 2018. Available online: http://www.fao.org/faostat/en/#data/RP/visualize (accessed on 25 May 2019).
- Pignati, W.A.; de Lima, F.A.N.S.; de Lara, S.S.; Correa, M.L.M.; Barbosa, J.R.; da Leão, L.H.C.; Pignatti, M.G. Distribuição espacial do uso de agrotóxicos no Brasil: Uma ferramenta para a Vigilância em Saúde (Spacial distribution of agrotoxics use in Brazil: A tool for Health Surveillance). Cien. Saude Colet. 2017, 22, 3281–3293. [Google Scholar] [CrossRef]
- Bebber, D.P.; Holmes, T.; Gurr, S.J. The global spread of crop pests and pathogens. Glob. Ecol. Biogeogr. 2014, 23, 1398–1407. [Google Scholar] [CrossRef]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Stewart, C.N. Becoming weeds. Nat. Genet. 2017, 49, 654–655. [Google Scholar] [CrossRef] [PubMed]
- Weed Science Society of America. Weeds. 2016. Available online: http://wssa.net/wssa/weed/ (accessed on 25 May 2019).
- Zimdahl, R.L. Fundamentals of Weed Science, 5th ed.; Academic Press: London, UK, 2018; 735p. [Google Scholar]
- Schonbeck, M. Principles of Sustainable Weed Management in Organic Cropping Systems, Workshop for Farmers and Agricultural Professionals on Sustainable Weed Management, 3rd ed.; Clemson University: Clemson, SC, USA, 2011; 20p. [Google Scholar]
- Brühl, C.A.; Zaller, J.G. Biodiversity decline as a consequence of an inappropriate environmental risk assessment of pesticides. Front. Environ. Sci. 2019, 7, 177. [Google Scholar] [CrossRef]
- Abbas, T.; Zahir, Z.A.; Naveed, M.; Kremer, R.J. Limitations of existing weed control practices necessitate development of alternative techniques based on biological approaches. Adv. Agron. 2018, 147, 239–280. [Google Scholar] [CrossRef]
- Cordeau, S.; Triolet, M.; Wayman, S.; Steinberg, C.; Guillemin, J.P. Bioherbicides: Dead in the water? A review of the existing products for integrated weed management. Crop Prot. 2016, 87, 44–49. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Mahajan, G. Recent Advances in Weed Management, 1st ed.; Springer: New York, NY, USA, 2014; 411p. [Google Scholar] [CrossRef]
- Dayan, F.E.; Duke, S.O. Natural compounds as next-generation herbicides. Plant Physiol. 2014, 166, 1090–1105. [Google Scholar] [CrossRef] [Green Version]
- Duke, S.O.; Pan, Z.; Bajsa-Hirschel, J.; Boyette, C.D. The potential future roles of natural compounds and microbial bioherbicides in weed management in crops. Adv. Weed Sci. 2022, 40, e020210054. [Google Scholar] [CrossRef]
- Puig, C.G.; Reigosa, M.J.; Valentão, P.; Andrade, P.B.; Pedrol, N. Unravelling the bioherbicide potential of Eucalyptus globulus Labill: Biochemistry and effects of its aqueous extract. PLoS ONE 2018, 13, e0192872. [Google Scholar] [CrossRef] [PubMed]
- De Costa, F.; Fleck, J.; Yendo, A.C.A.; Gosmann, G.; Fett-Neto, A.G. Accumulation of a bioactive triterpene saponin fraction of Quillaja brasiliensis leaves is associated with abiotic and biotic stresses. Plant Physiol. Biochem. 2013, 66, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Patterson, N.; Zaharia, L.I. Saponin biosynthesis in pulses. Plants 2022, 11, 3505. [Google Scholar] [CrossRef] [PubMed]
- Magedans, Y.V.S.; Phillips, M.A.; Fett-Neto, A.G. Production of plant bioactive triterpenoid saponins: From metabolites to genes and back. Phytochem. Rev. 2021, 20, 461–482. [Google Scholar] [CrossRef]
- Faizal, A.; Geelen, D. Saponins and their role in biological processes in plants. Phytochem. Rev. 2013, 12, 877–893. [Google Scholar] [CrossRef]
- Hoagland, R.E.; Zablotowicz, R.M.; Reddy, K.N. Studies of the phytotoxicity of saponins on weed and crop plants. In Saponins Used in Food and Agriculture; Waller, G.R., Yamasaki, K., Eds.; Plenum Press: New York, NY, USA, 1996; pp. 57–73. [Google Scholar]
- Jelassi, A.; Ayeb-Zakhama, A.E.; Nejma, A.B.; Chaari, A.; Harzallah-Skhiri, F.; Jannet, H.B. Phytochemical composition and allelopathic potential of three Tunisian Acacia species. Ind. Crops Prod. 2016, 83, 339–345. [Google Scholar] [CrossRef]
- Pérez, A.J.; Simonet, A.M.; Pecio, Ł.; Kowalczyk, M.; Calle, J.M.; Macías, F.A.; Oleszec, W.; Stochmal, A. Triterpenoid saponins from the aerial parts of Trifolium argutum Sol. and their phytotoxic evaluation. Phytochem. Lett. 2015, 13, 165–170. [Google Scholar] [CrossRef]
- Ribeiro, R.C.; de Carvalho, M.G.; de Moraes, M.L.L.; Rossiello, R.O.P.; de Oliveira, D.R.; de Amorim, R.M.Q.; Barbieri, E., Jr. Chemical screening of Urochloa humidicola: Methods for characterizing secondary metabolites and allelopathic activity on forage legumes. Am. J. Plant Sci. 2018, 9, 1260–1278. [Google Scholar] [CrossRef]
- Stavropoulou, M.I.; Angelis, A.; Aligiannis, N.; Kalpoutzakis, E.; Mitakou, S.; Duke, S.O.; Fokialakis, N. Phytotoxic triterpene saponins from Bellis longifolia, an endemic plant of Crete. Phytochemistry 2017, 144, 71–77. [Google Scholar] [CrossRef]
- Wyman-Simpson, C.L.; Waller, G.R.; Jurzysta, M.; McPherson, J.K.; Young, C.C. Biological activity and chemical isolation of root saponins of six cultivars of alfalfa (Medicago sativa L.). Plant Soil. 1991, 135, 83–94. [Google Scholar] [CrossRef]
- Reitz, R.; Reis, A.; Klein, R.M. Flora Ilustrada Catarinense—Rosáceas (Illustrated Flora of the State of Santa Catarina—Rosaceae); Herbário Barbosa Rodrigues: Itajaí, Brazil, 1996; pp. 113–116. [Google Scholar]
- Fleck, J.D.; Kauffmann, C.; Spilki, F.; Lencina, C.L.; Roehe, P.M.; Gosmann, G. Adjuvant activity of Quillaja brasiliensis saponins on the immune responses to bovine herpesvirus type 1 in mice. Vaccine 2006, 24, 7129–7134. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sharma, A.; Tirpude, N.V.; Padwad, Y.; Hallan, V.; Kumar, S. Plant-derived immuno-adjuvants in vaccines formulation: A promising avenue for improving vaccines efficacy against SARS-CoV-2 virus. Pharmacol. Rep. 2022, 74, 1238–1254. [Google Scholar] [CrossRef] [PubMed]
- Yendo, A.C.A. Saponinas de Quillaja brasiliensis: Purificação e atividade biológica. (Saponins of Quillaja brasiliensis: Purification and biological activity). Ph.D. Thesis, Graduate Program in Botany—Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre, Brazil, 2016. [Google Scholar]
- Kauffmann, C.; Machado, A.M.; Fleck, J.D.; Provensi, G.; Pires, V.S.; Guillaume, D.; Sonnet, P.; Reginatto, F.H.; Shenkel, E.P.; Gosmann, G. Constituents from leaves of Quillaja brasiliensis. Nat. Prod. Res. 2004, 18, 153–157. [Google Scholar] [CrossRef]
- González-Cruz, D.; Martín, R.S. Molluscicidal effects of saponin-rich plant extracts on the grey field slug. Cien. Investig. Agr. 2013, 40, 341–349. [Google Scholar] [CrossRef]
- De Geyter, E.; Smagghe, G.; Rahbe, Y.; Geelen, D. Triterpene saponins of Quillaja saponaria show strong aphicidal and deterrent activity against the pea aphid Acyrthosiphon pisum. Pest Manag. Sci. 2011, 68, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Moya, E.; San, R.M.; Gamboa, M.; Hidalgo, G.E.A. Evaluation of a Quillaja saponaria saponin extract for control of powdery mildew of wheat and squash evaluación de un extracto de saponinas de Quillaja saponaria para el control de oídios de trigo y zapallo. Agro Sur 2010, 38, 87–96. [Google Scholar] [CrossRef]
- González-Castillo, J.A.; Quezada-D’Angelo, T.P.; Silva-Aguayo, G.I.; Moya, E.A. Effect of saponins of Quillaja saponaria extracts in combination with Pseudomonas protegens to control Gaeumannomyces graminis var. tritici in wheat. Chil. J. Agric Res. 2018, 78, 378–390. [Google Scholar] [CrossRef]
- Giannakou, I.O. Efficacy of a formulated product containing Quillaja saponaria plant extracts for the control of root-knot nematodes. Eur. J. Plant Pathol. 2011, 130, 587. [Google Scholar] [CrossRef]
- Martín, R.; Briones, R. Industrial uses and sustainable supply of Quillaja saponaria (Rosaceae) saponins. Econ. Bot. 1999, 53, 302–311. [Google Scholar] [CrossRef]
- Meneses, C.H.S.G.; Lima, L.H.G.M.; Lima, M.M.A.; Pereira, W.E.; Bruno, R.L.A.; Vidal, M.S. Potencial hídrico induzido por Polietilenoglicol-6000 na viabilidade de sementes de algodão (Water potential induced by Polyethylene glycol-6000 on cotton seed viability). Embrapa Agrobiol. 2007, 19, 1–22. [Google Scholar]
- Sadeghloo, A.; Asghari, J.; Ghaderi-Far, F. Seed germination and seedling emergence of velvetleaf (Abutilon theophrasti) and Barnyardgrass (Echinochloa crus-galli). Planta Daninha 2013, 31, 259–266. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press Inc.: New York, NY, USA, 1984; 422p. [Google Scholar]
- Yendo, A.C.A.; De Costa, F.; Da Costa, C.T.; Colling, L.C.; Gosmann, G.; Fett-Neto, A.G. Biosynthesis of plant triterpenoid saponins: Genes, enzymes, and their regulation. Mini Rev. Org. Chem. 2014, 11, 292–306. [Google Scholar] [CrossRef]
- Poonpaiboonpipat, T.; Pangnakorn, U.; Suvunnamek, U.; Teerarak, M.; Charoenying, P.; Laosinwattana, C. Phytotoxic effects of essential oil from Cymbopogon citratus and its physiological mechanisms on barnyardgrass (Echinochloa crus-galli). Ind. Crop. Prod. 2013, 41, 403–407. [Google Scholar] [CrossRef]
- De Costa, F.; Yendo, A.C.A.; Fleck, J.D.; Gosmann, G.; Fett-Neto, A.G. Immunoadjuvant and anti-inflammatory plant saponins: Characteristics and biotechnological approaches towards sustainable production. Mini Rev. Med. Chem. 2011, 11, 857–880. [Google Scholar] [CrossRef]
- De Groot, C.; Müller-Goymann, C.C. Saponin interactions with model membrane systems—Langmuir monolayer studies, hemolysis and formation of ISCOMs. Planta Med. 2016, 82, 1496–1512. [Google Scholar] [CrossRef]
- Wallace, F.; Bennadji, Z.; Ferreira, F.; Olivaro, C. Analysis of an immunoadjuvant saponin fraction from Quillaja brasiliensis leaves by electrospray ionization ion trap multiple-stage mass spectrometry. Phytochem. Lett. 2017, 20, 228–233. [Google Scholar] [CrossRef]
- Magedans, Y.V.S.; Phillips, M.A. Soapbark Triterpenes: Quillaja brasiliensis cell culture sapogenin and free sterol analysis by GCMS. Methods Mol. Biol. 2022, 2469, 119–128. [Google Scholar] [CrossRef]
- Popinigis, F. Fisiologia da Semente (Seed Physiology); Brazilian Ministry of Agriculture: Brasíla, Brazil; Inter-American Development Bank (BID): Washington, DC, USA, 1977; pp. 275–277.
- Newcombe, R.G. Two-sided confidence intervals for the single proportion: Comparison of seven methods. Statist. Med. 1998, 17, 857–872. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, M.E.M.; de Carvalho, A.C.; Yendo, A.C.A.; Magedans, Y.V.S.; Zachert, E.; Fett-Neto, A.G. Phytotoxicity of Quillaja lancifolia Leaf Saponins and Their Bioherbicide Potential. Plants 2023, 12, 663. https://doi.org/10.3390/plants12030663
Marques MEM, de Carvalho AC, Yendo ACA, Magedans YVS, Zachert E, Fett-Neto AG. Phytotoxicity of Quillaja lancifolia Leaf Saponins and Their Bioherbicide Potential. Plants. 2023; 12(3):663. https://doi.org/10.3390/plants12030663
Chicago/Turabian StyleMarques, Maria E. M., Ana C. de Carvalho, Anna C. A. Yendo, Yve V. S. Magedans, Eliane Zachert, and Arthur G. Fett-Neto. 2023. "Phytotoxicity of Quillaja lancifolia Leaf Saponins and Their Bioherbicide Potential" Plants 12, no. 3: 663. https://doi.org/10.3390/plants12030663
APA StyleMarques, M. E. M., de Carvalho, A. C., Yendo, A. C. A., Magedans, Y. V. S., Zachert, E., & Fett-Neto, A. G. (2023). Phytotoxicity of Quillaja lancifolia Leaf Saponins and Their Bioherbicide Potential. Plants, 12(3), 663. https://doi.org/10.3390/plants12030663