

Influence of Plant-Based Biostimulant (BORTAN) on Qualitative and Aromatic Traits of Rocket Salad (Diplotaxis tenuifolia L.)

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
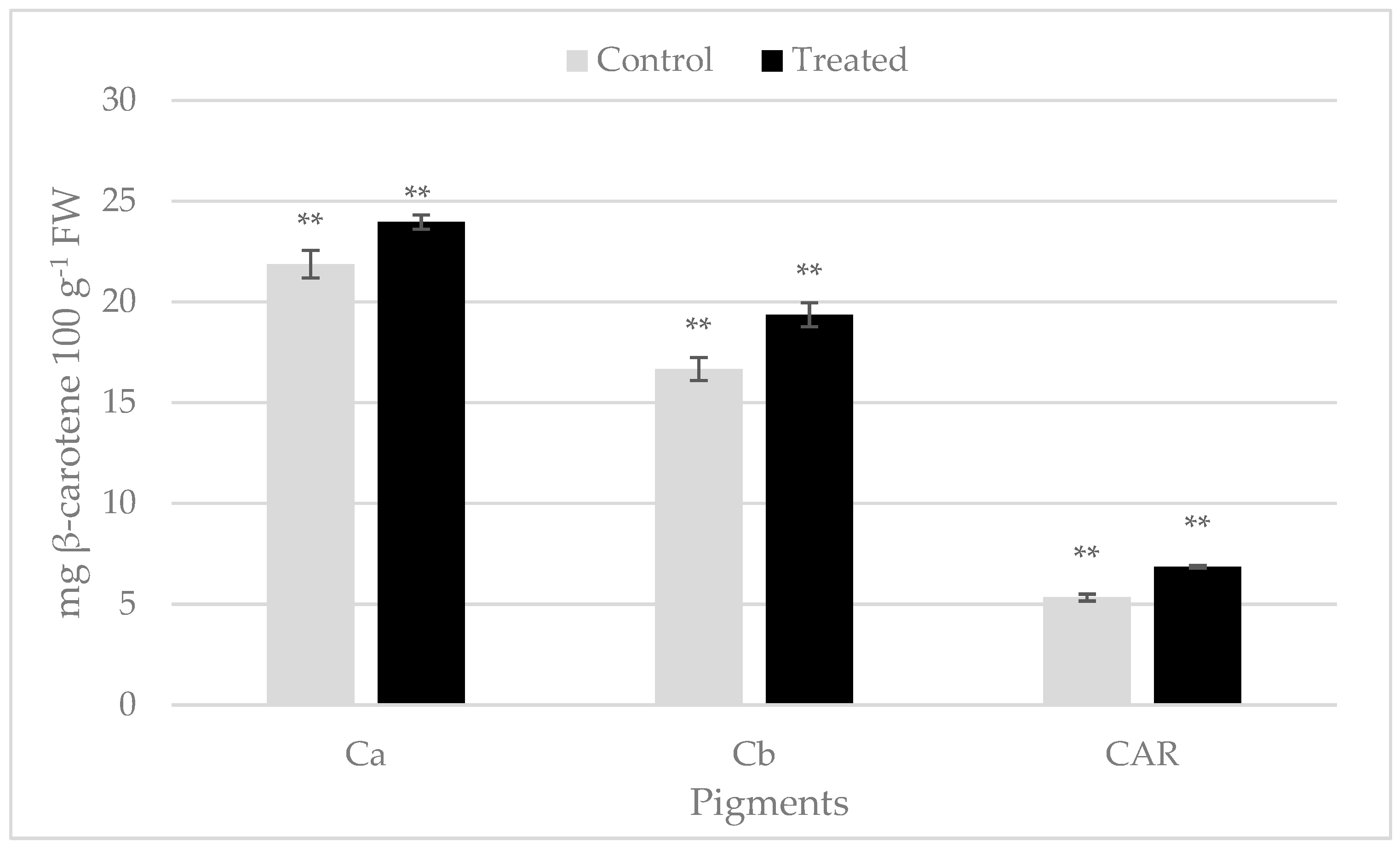
2.1. Colorimetric Parameters and Leaf Chlorophyll Content
2.2. Bioactive Compounds and Ascorbic Acid Content
2.3. Volatile Compounds Analysis
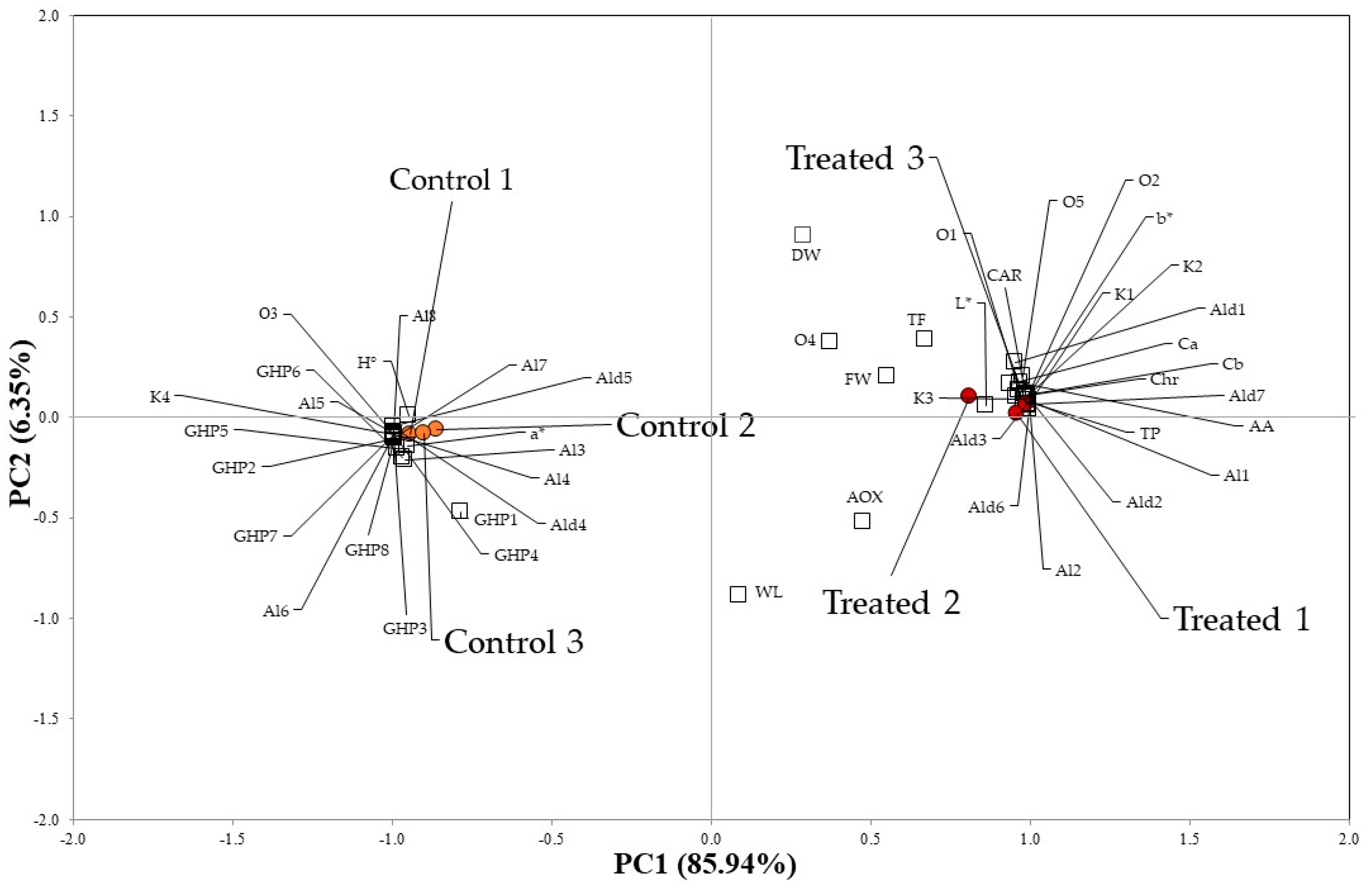
2.4. Correlation Analysis among Physico-Chemical, Biochemical and VOCs Data
3. Materials and Methods
3.1. Plant Material and Growth Conditions
3.2. Physical Parameters
3.3. Photosynthetic Pigments
3.4. Bioactive Compounds
3.5. Ascorbic Acid Content
3.6. Sample Preparation and SPME Procedure
3.7. Gas Chromatography–Quadrupole Mass Spectrometry Analysis (GC–qMS)
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Blattner, C. Just Transition for Agriculture? A Critical Step in Tackling Climate Change. J. Agric. Food Syst. Community Dev. 2020, 9, 53–58. [Google Scholar] [CrossRef]
- Del Buono, D. Can biostimulants be used to mitigate the effect of anthropogenic climate change on agriculture? It is time to respond. Sci. Total Environ. 2021, 751, 141763. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef]
- Malik, A.; Mor, V.S.; Tokas, J.; Punia, H.; Malik, S.; Malik, K.; Sangwan, S.; Tomar, S.; Singh, P.; Singh, N.; et al. Biostimulant-Treated Seedlings under Sustainable Agriculture: A Global Perspective Facing Climate Change. Agronomy 2021, 11, 14. [Google Scholar] [CrossRef]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Senatore, M.; Giordano, M.; El-Nakhel, C.; Sacco, A.; Rouphael, Y.; Colla, G.; Mori, M. Plant-Based Biostimulants Influence the Agronomical, Physiological, and Qualitative Responses of Baby Rocket Leaves under Diverse Nitrogen Conditions. Plants 2019, 8, 522. [Google Scholar] [CrossRef] [PubMed]
- Campobenedetto, C.; Agliassa, C.; Mannino, G.; Vigliante, I.; Contartese, V.; Secchi, F.; Bertea, C. A Biostimulant Based on Seaweed (Ascophyllum nodosum and Laminaria digitata) and Yeast Extracts Mitigates Water Stress Effects on Tomato (Solanum lycopersicum L.). Agriculture 2021, 11, 557. [Google Scholar] [CrossRef]
- Toscano, S.; Romano, D.; Massa, D.; Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulant applications in low input horticultural cultivation systems. Italus Hortus 2019, 25, 27–36. [Google Scholar] [CrossRef]
- Xu, L.; Geelen, D. Developing Biostimulants From Agro-Food and Industrial By-Products. Front. Plant Sci. 2018, 9, 1567. [Google Scholar] [CrossRef]
- Franzoni, G.; Cocetta, G.; Prinsi, B.; Ferrante, A.; Espen, L. Biostimulants on Crops: Their Impact under Abiotic Stress Conditions. Horticulturae 2022, 8, 189. [Google Scholar] [CrossRef]
- Assainar, S.K.; Abbott, L.; Mickan, B.S.; Whiteley, A.; Siddique, K.; Solaiman, Z.M. Response of Wheat to a Multiple Species Microbial Inoculant Compared to Fertilizer Application. Front. Plant Sci. 2018, 9, 1601. [Google Scholar] [CrossRef] [Green Version]
- Bell, L.; Oruna Concha, M.J.; Wagstaff, C. Identification and quantification of glucosinolate and flavonol compounds in rocket salad (Eruca sativa, Eruca vesicaria and Diplotaxis tenuifolia) by LC−MS: Highlighting the potential for improving nutritional value of rocket crops. Food Chem. 2015, 172, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Bell, L.; Wagstaff, C. Glucosinolates, myrosinase hydrolysis products, and flavonols found in rocket (Eruca sativa and Diplo-taxis tenuifolia). J. Agric. Food Chem. 2014, 62, 4481–4492. [Google Scholar] [CrossRef] [PubMed]
- Aprea, E.; Corollaro, M.L.; Betta, E.; Endrizzi, I.; Dematte, M.L.; Biasioli, F. Sensory and instrumental profiling of 18 apple cultivars to investigate the relation between perceived quality and odour and flavor. Food Res. Int. 2012, 49, 677–686. [Google Scholar] [CrossRef]
- Besada, C.; Sanchez, G.; Salvador, A.; Granell, A. Volatile compounds associated to the loss of astringency in persimmon fruit revealed by untargeted GC–MS analysis. Metabolomics 2013, 9, 157–172. [Google Scholar] [CrossRef]
- Palumbo, M.; Cozzolino, R.; Laurino, C.; Malorni, L.; Picariello, G.; Siano, F.; Stocchero, M.; Cefola, M.; Corvino, A.; Romaniello, R.; et al. Rapid and Non-Destructive Techniques for the Discrimination of Ripening Stages in Candonga Strawberries. Foods 2022, 11, 1534. [Google Scholar] [CrossRef] [PubMed]
- Mastrandrea, L.; Amodio, M.L.; Pati, S.; Colelli, G. Effect of modified atmosphere packaging and temperature abuse on flavor related volatile compounds of rocket leaves (Diplotaxis tenuifolia L.). J. Food Sci. Technol. 2017, 54, 2433–2442. [Google Scholar] [CrossRef]
- Cristofano, F.; El-Nakhel, C.; Rouphael, Y. Biostimulant Substances for Sustainable Agriculture: Origin, Operating Mechanisms and Effects on Cucurbits, Leafy Greens, and Nightshade Vegetables Species. Biomolecules 2021, 11, 1103. [Google Scholar] [CrossRef]
- Caruso, G.; De Pascale, S.; Cozzolino, E.; Giordano, M.; El-Nakhel, C.; Cuciniello, A.; Cenvinzo, V.; Colla, G.; Rouphael, Y. Protein Hydrolysate or Plant Extract-based Biostimulants Enhanced Yield and Quality Performances of Greenhouse Perennial Wall Rocket Grown in Different Seasons. Plants 2019, 5, 208. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, A.; Incrocci, L.; Maggini, R.; Serra, G.; Tognoni, F. Colour changes of fresh-cut leafy vegetables during storage. J. Food Agric. Env. 2004, 22, 40–44. [Google Scholar]
- Franzoni, G.; Bulgari, R.; Ferrante, A. Maceration Time Affects the Efficacy of Borage Extracts as Potential Biostimulant on Rocket Salad. Agronomy 2021, 11, 2182. [Google Scholar] [CrossRef]
- Zappia, A.; De Bruno, A.; Piscopo, A.; Poiana, M. Physico-chemical and microbiological quality of ready-to-eat rocket (Eruca vesicaria (L.) Cav.) treated with organic acids during storage in dark and light conditions. Food Sci. Biotechnol. 2019, 28, 965–973. [Google Scholar] [CrossRef]
- Elmardy, N.A.; Yousef, A.F.; Lin, K.; Zhang, X.; Ali, M.M.; Lamlom, S.F.; Kalaji, H.M.; Kowalczyk, K.; Xu, Y. Photosynthetic performance of rocket (Eruca sativa. Mill.) grown under different regimes of light intensity, quality, and photoperiod. PLoS ONE 2021, 16, e0257745. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Cheng, Z.-H.; Hayat, S.; Ahmad, H.; Ghani, M.I.; Liu, T. Foliar spraying of aqueous garlic bulb extract stimulates growth and antioxidant enzyme activity in eggplant (Solanum melongena L.). J. Integr. Agric. 2019, 18, 1001–1013. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Canaguier, R.; Kumar, P.; Colla, G. The effect of a plant-derived biostimulant on metabolic profiling and crop performance of lettuce grown under saline conditions. Sci. Hortic. 2015, 182, 124–133. [Google Scholar] [CrossRef]
- Schiattone, M.I.; Boari, F.; Cantore, V.; Castronuovo, D.; Denora, M.; Di Venere, D.; Perniola, M.; Renna, M.; Sergio, L.; Candido, V. Effects of Nitrogen, Azoxystrobin and a Biostimulant Based on Brown Algae and Yeast on Wild Rocket Features at Harvest and During Storage. Agronomy 2021, 11, 2326. [Google Scholar] [CrossRef]
- Candido, V.; Boari, F.; Cantore, V.; Castronuovo, D.; Denora, M.; Sergio, L.; Todorovic, M.; Schiattone, M.I. Interactive effect of water regime, nitrogen rate and biostimulant application on physiological and biochemical traits of wild rocket. Agric. Water Manag. 2023, 277, 108075. [Google Scholar] [CrossRef]
- Schiattone, M.I.; Boari, F.; Cantore, V.; Castronuovo, D.; Denora, M.; Di Venere, D.; Perniola, M.; Sergio, L.; Todorovic, M.; Candido, V. Effect of water regime, nitrogen level and biostimulants application on yield and quality traits of wild rocket (Diplotaxis tenuifolia (L.) DC. Agric. Water Manag. 2023, 277, 108078. [Google Scholar] [CrossRef]
- Berti, M.; Wilckens, R.; Fischer, S.; Araos, R. Borage: A new crop for Southern Chile. In Trends New Crop New Uses; ASHS Press: Alexandria, VA, USA, 2002. [Google Scholar]
- Berti, M.; Fischer, S.; Wilckens, R.; Hevia, F.; Johnson, B.L. BORAGE (Borago officinalis L.) Response to N, P, K, and S fertilization in South Central Chile. Chil. J. Agric. Res. 2010, 70, 228–236. [Google Scholar] [CrossRef]
- Di Gioia, F.; Tzortzakis, N.; Rouphael, Y.; Kyriacou, M.C.; Sampaio, S.L.; Ferreira, I.C.; Petropoulos, S.A. Grown to Be Blue—Antioxidant Properties and Health Effects of Colored Vegetables. Part II: Leafy, Fruit, and Other Vegetables. Antioxidants 2020, 9, 97. [Google Scholar] [CrossRef]
- Pereira, M.; Amaro, A.; Oliveira, A.; Pintado, M. Bioactive compounds in ready-to-eat rocket leaves as affected by oxygen partial pressure and storage time: A kinetic modelling. Postharvest Biol. Technol. 2019, 158, 110985. [Google Scholar] [CrossRef]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic compounds in plants and agri-industrialby-products: Antioxidant activity, occurrence and potential uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Conversa, G.; Bonasia, A.; Lazzizera, C.; Elia, A. Pre-harvest nitrogen and azoxystrobin application enhances raw product quality and post-harvest shelf-life of baby spinach (Spinacia oleracea L.). J. Sci. Food Agric. 2014, 94, 3263–3272. [Google Scholar] [CrossRef]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.C.; Bonini, P.; Colla, G. Plant and sea-weed-based extracts increase yield but differentially modulate nutritional quality of greenhouse spinach through biostimulant action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef]
- Trejo Valencia, R.; Sanchez Acosta, L.; Fortis Hernandez, M.; Preciado Rangel, P.; Gallegos Robles, M.A.; Antonio Cruz, R.D.C.; Vazquez Vazquez, C. Effect of seaweed aqueous extracts and compost on vegetative growth, yield, and nutraceutical quality of cucumber (Cucumis sativus L.) fruit. Agronomy 2018, 8, 264. [Google Scholar] [CrossRef]
- Kocira, A.; Lamorska, J.; Kornas, R.; Nowosad, N.; Tomaszewska, M.; Leszczynska, D.; Kozłowicz, K.; Tabor, S. Changes in biochemistry and yield in response to biostimulants applied in bean (Phaseolus vulgaris L.). Agronomy 2020, 10, 189. [Google Scholar] [CrossRef]
- Lawal, O.O.; Essien, N.C.; Essien, N.M.; Ochalla, F. Vitamin C content of some processed green leafy vegetables. Eur. J. Exp. Biol. 2015, 5, 110–112. [Google Scholar]
- Matev, G.; Dimitrova, P.; Petkova, N.; Ivanov, I.; Mihaylova, D. Antioxidant activity and mineral content of rocket (Eruca sativa) plant from italian and bulgarian origins. J. Microbiol. Biotechnol. Food Sci. 2018, 8, 756–759. [Google Scholar] [CrossRef]
- Jirovetz, L.; Smith, D.; Buchbauer, G. Aroma compound analysis of Eruca sativa (Brassicaceae) SPME headspace leaf samples using GC, GC−MS, and olfactometry. J. Agric. Food Chem. 2002, 50, 4643–4646. [Google Scholar] [CrossRef]
- Blaževic, I.; Mastelic, J. Free and bound volatiles of rocket (Eruca sativa Mill.). J. Flavour Fragr. 2008, 23, 278–285. [Google Scholar] [CrossRef]
- Spadafora, N.D.; Amaro, A.L.; Pereira, M.J.; Müller, C.T.; Pintado, M.; Rogers, H.J. Multi-trait analysis of post-harvest storage in rocket salad (Diplotaxis tenuifolia) links sensorial, volatile and nutritional data. Food Chem. 2016, 211, 114–123. [Google Scholar] [CrossRef]
- Raffo, A.; Masci, M.; Moneta, E.; Nicoli, S.; del Pulgar, J.S.; Paoletti, F. Characterization of volatiles and identification of odor-active compounds of rocket leaves. Food Chem. 2018, 240, 1161–1170. [Google Scholar] [CrossRef]
- Abouelenein, D.; Angeloni, S.; Caprioli, G.; Genovese, J.; Mustafa, A.M.; Nzekoue, F.K.; Petrelli, R.; Rocculi, P.; Sagratini, G. Tappi, Effect of Plasma Activated Water on Selected Chemical Compounds of Rocket-Salad (Eruca sativa Mill.) Leaves. Molecules 2021, 26, 7691. [Google Scholar] [CrossRef] [PubMed]
- Bell, L.; Kitsopanou, E.; Oloyede, O.O.; Lignou, S. Important Odorants of Four Brassicaceae Species, and Discrepancies between Glucosinolate Profiles and Observed Hydrolysis Products. Foods 2021, 10, 1055. [Google Scholar] [CrossRef]
- Arora, R.; Kumar, R.; Mahajan, J.; Vig, A.P.; Singh, B.; Singh, B.; Arora, S. 3-Butenyl isothiocyanate: A hydrolytic product of glucosinolate as a potential cytotoxic agent against human cancer cell lines. J. Food Sci. Technol. 2016, 53, 3437–3445. [Google Scholar] [CrossRef] [PubMed]
- Bell, L.; Methven, L.; Signore, A.; Oruna-Concha, M.J.; Wagstaff, C. Analysis of seven salad rocket (Eruca sativa) accessions: The relationships between sensory attributes and volatile and non-volatile compounds. Food Chem. 2017, 218, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Zheng, F.; Tian, X.; Feng, T.; Yao, L.; Sun, M.; Shi, L. Evolution Analysis of Free Fatty Acids and Aroma-Active Compounds during Tallow Oxidation. Molecules 2022, 27, 352. [Google Scholar] [CrossRef]
- Huseby, S.; Koprivova, A.; Lee, B.-R.; Saha, S.; Mithen, R.; Wold, A.-B.; Bengtsson, G.; Kopriva, S. Diurnal and light regulation of sulphur assimilation and glucosinolate biosynthesis in Arabidopsis. J. Exp. Bot. 2013, 64, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Magri, A.; Petriccione, M. Melatonin treatment reduces qualitative decay and improves antioxidant system in highbush blueberry fruit during cold storage. J. Sci. Food Agric. 2022, 102, 4229–4237. [Google Scholar] [CrossRef]
- Magri, A.; Adiletta, G.; Petriccione, M. Evaluation of Antioxidant Systems and Ascorbate-Glutathione Cycle in Feijoa Edible Flowers at Different Flowering Stages. Foods 2020, 9, 95. [Google Scholar] [CrossRef]
- Goffi, V.; Magri, A.; Botondi, R.; Petriccione, M. Response of antioxidant system to postharvest ozone treatment in ‘Soreli’ kiwifruit. J. Sci. Food Agric. 2020, 100, 961–968. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Samples | L* | A* | B* | Chr | H° | WL (%) |
|---|---|---|---|---|---|---|
| Treated | 39.97 ± 1.96 | −8.23 ± 0.57 | 19.38 ± 1.40 | 21.00 ± 1.55 | 115.62 ± 0.90 | 97.80 ± 0.04 |
| Control | 34.59 ± 2.48 | −7.19 ± 0.61 | 14.98 ± 1.29 | 16.62 ± 1.40 | 113.10 ± 0.46 | 97.80 ± 0.84 |
| p | ** | ** | *** | *** | ** | ns |
| Samples | TP (mg GAE 100 g FW) | TF (mg CE 100 g FW) | AA (mg AA 100 g FW) | AOX (µmol TE g FW) |
|---|---|---|---|---|
| Treated | 91.94 ± 5.04 | 102.51 ± 7.00 | 16.63 ± 0.69 | 9.99 ± 0.20 |
| Control | 68.56 ± 3.45 | 81.56 ± 2.29 | 14.04 ± 0.05 | 8.61 ± 0.49 |
| p | *** | *** | ** | ** |
| Metabolites | Code | RIcal/RIt | ID | Control | Treated | p |
|---|---|---|---|---|---|---|
| Ketones | ||||||
| 3-Pentanone | K1 | 980/980 | RI/MS/S | 2.86 | 4.07 | ** |
| 1-Penten-3-one | K2 | 1026/1026 | RI/MS/S | 4.66 | 15.55 | ** |
| 3-Octanone | K3 | 1272/1272 | RI/MS/S | 7.46 | 15.41 | ** |
| 6-Methyl-5-hepten-2-one | K4 | 1350/1348 | RI/MS/S | 3.30 | 0.00 | ** |
| Aldehydes | ||||||
| Hexanal | Ald1 | 1084/1086 | RI/MS/S | 11.45 | 14.38 | ** |
| cis-3-Hexanal | Ald2 | 1148/1148 | RI/MS | 6.45 | 58.64 | ** |
| 2-Hexenal | Ald3 | 1242/1248 | RI/MS/S | 245.48 | 722.66 | ** |
| Octanal | Ald4 | 1309/1308 | RI/MS/S | 1.79 | 0.00 | ** |
| Nonanal | Ald5 | 1401/1401 | RI/MS/S | 6.83 | 1.18 | ** |
| 2,4-Heptadienal | Ald6 | 1401/1401 | RI/MS/S | 0.62 | 2.59 | ** |
| Decanal | Ald7 | 1506/1505 | RI/MS/S | 7.86 | 10.08 | ** |
| Alcohols | ||||||
| 1-Penten-3-ol | Al1 | 1188/1189 | RI/MS/S | 18.53 | 48.78 | ** |
| cis-2-penten-1-ol | Al2 | 1272/1272 | RI/MS/S | 0.00 | 21.60 | ** |
| 1-Hexanol | Al3 | 1340/1339 | RI/MS/S | 8.56 | 6.55 | ** |
| trans-3-Hexen-1-ol | Al4 | 1367/1366 | RI/MS/S | 8.65 | 2.76 | ** |
| cis-3-Hexen-1-ol | Al5 | 1374/1374 | RI/MS/S | 382.28 | 148.21 | ** |
| trans-2-Hexen-1-ol | Al6 | 1394/1394 | RI/MS/S | 60.91 | 23.61 | ** |
| 2,6-Dimethylcyclohexanol | Al7 | - | MS/S | 1.14 | 0.00 | ** |
| Terpinen-4-ol | Al8 | 1609/1609 | RI/MS/S | 1.53 | 0.00 | ** |
| Glucosinolate Hydrolysis Products (GHPs) | ** | |||||
| Methyl ITC | GHP1 | 1282/1278 | RI/MS/S | 6.30 | 5.55 | * |
| 3-Butenyl ITC | GHP2 | 1479/1463 | RI/MS | 2.41 | 0.00 | ** |
| Pentyl ITC | GHP3 | 1511/1540 | RI/MS | 15.32 | 3.13 | ** |
| 4-Methylpentyl ITC | GHP4 | 1487/1490 | RI/MS | 84.32 | 70.32 | ** |
| Hexyl ITC | GHP5 | 1582/1588 | RI/MS/S | 18.59 | 15.35 | ** |
| 3-Methylthiopropyl ITC | GHP6 | 1970/1979 | RI/MS/S | 11.55 | 10.28 | ** |
| Benzyl ITC | GHP7 | 2109/2109 | RI/MS/S | 7.03 | 1.90 | ** |
| 2-Ethylphenyl ITC | GHP8 | 2230/2233 | RI/MS | 4.53 | 2.21 | ** |
| Others | ** | |||||
| 2-Ethylfuran | O1 | 960/965 | RI/MS/S | 4.72 | 7.60 | ** |
| 1-Pentene-3-ethyl-2-methyl | O2 | - | MS/S | 5.46 | 12.29 | ** |
| cis-3-Hexen-1-ol acetate | O3 | 1320/1320 | RI/MS/S | 38.09 | 1.64 | ** |
| Anisole | O4 | 1350/1350 | RI/MS/S | 10.34 | 10.43 | ns |
| 5-Methylhexanenitrile | O5 | 1362/1358 | RI/MS | 2.54 | 5.61 | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malorni, L.; Cozzolino, R.; Magri, A.; Zampella, L.; Petriccione, M. Influence of Plant-Based Biostimulant (BORTAN) on Qualitative and Aromatic Traits of Rocket Salad (Diplotaxis tenuifolia L.). Plants 2023, 12, 730. https://doi.org/10.3390/plants12040730
Malorni L, Cozzolino R, Magri A, Zampella L, Petriccione M. Influence of Plant-Based Biostimulant (BORTAN) on Qualitative and Aromatic Traits of Rocket Salad (Diplotaxis tenuifolia L.). Plants. 2023; 12(4):730. https://doi.org/10.3390/plants12040730
Chicago/Turabian StyleMalorni, Livia, Rosaria Cozzolino, Anna Magri, Luigi Zampella, and Milena Petriccione. 2023. "Influence of Plant-Based Biostimulant (BORTAN) on Qualitative and Aromatic Traits of Rocket Salad (Diplotaxis tenuifolia L.)" Plants 12, no. 4: 730. https://doi.org/10.3390/plants12040730
APA StyleMalorni, L., Cozzolino, R., Magri, A., Zampella, L., & Petriccione, M. (2023). Influence of Plant-Based Biostimulant (BORTAN) on Qualitative and Aromatic Traits of Rocket Salad (Diplotaxis tenuifolia L.). Plants, 12(4), 730. https://doi.org/10.3390/plants12040730