

Phytochemical Profile and In Vitro Bioactivities of Plant-Based By-Products in View of a Potential Reuse and Valorization
 , ,
, , 
 and
and
Abstract
:1. Introduction
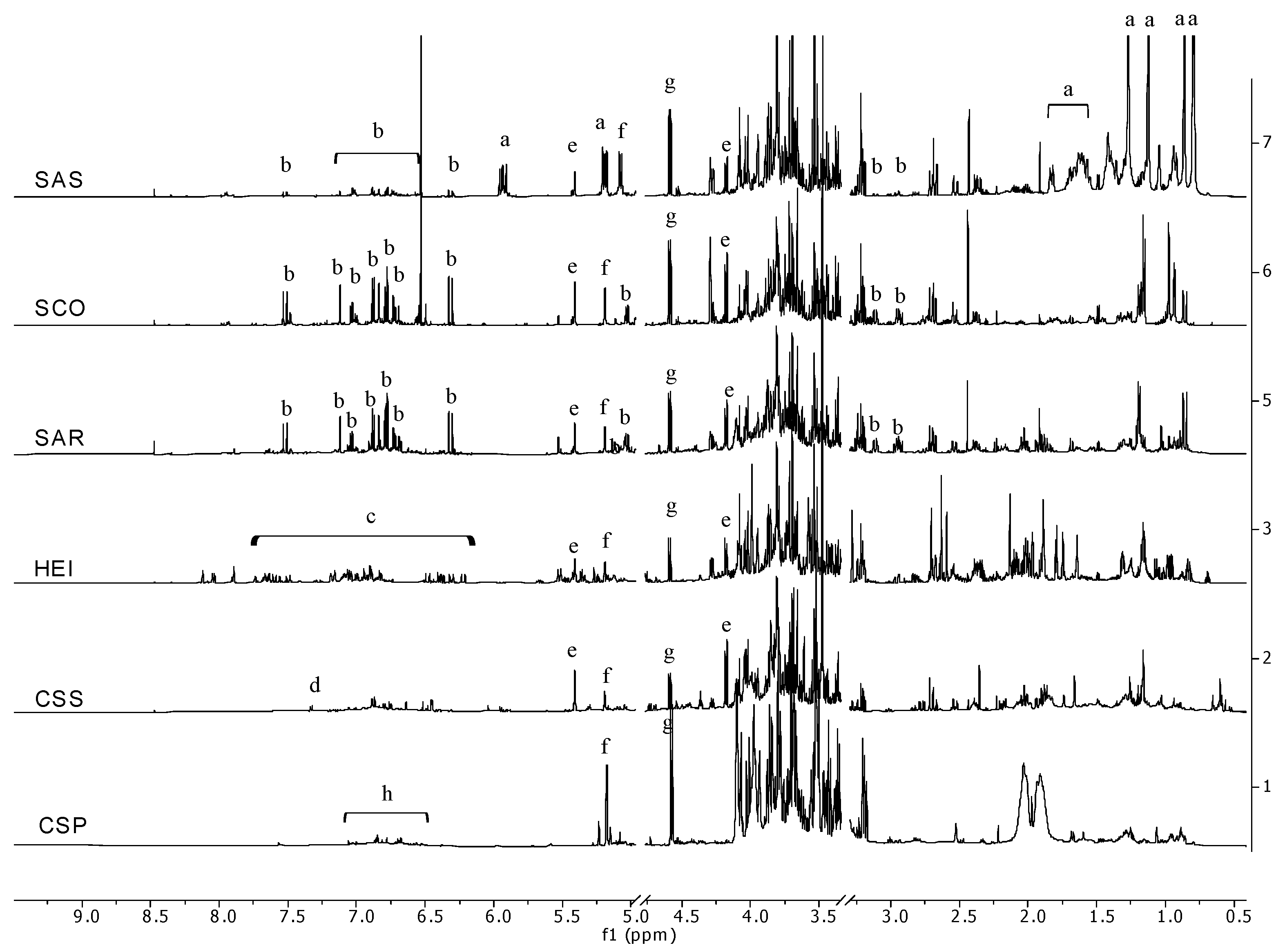
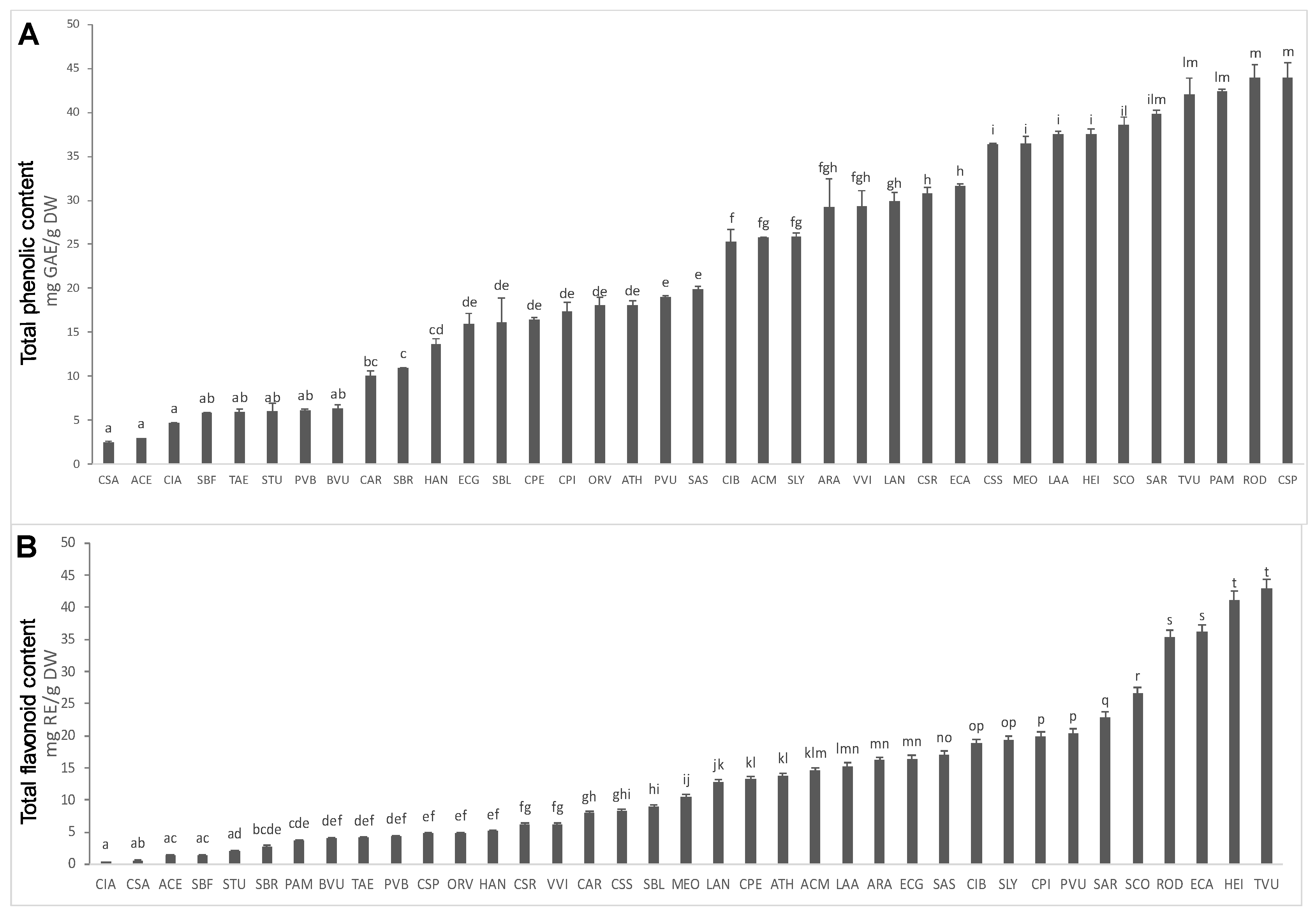
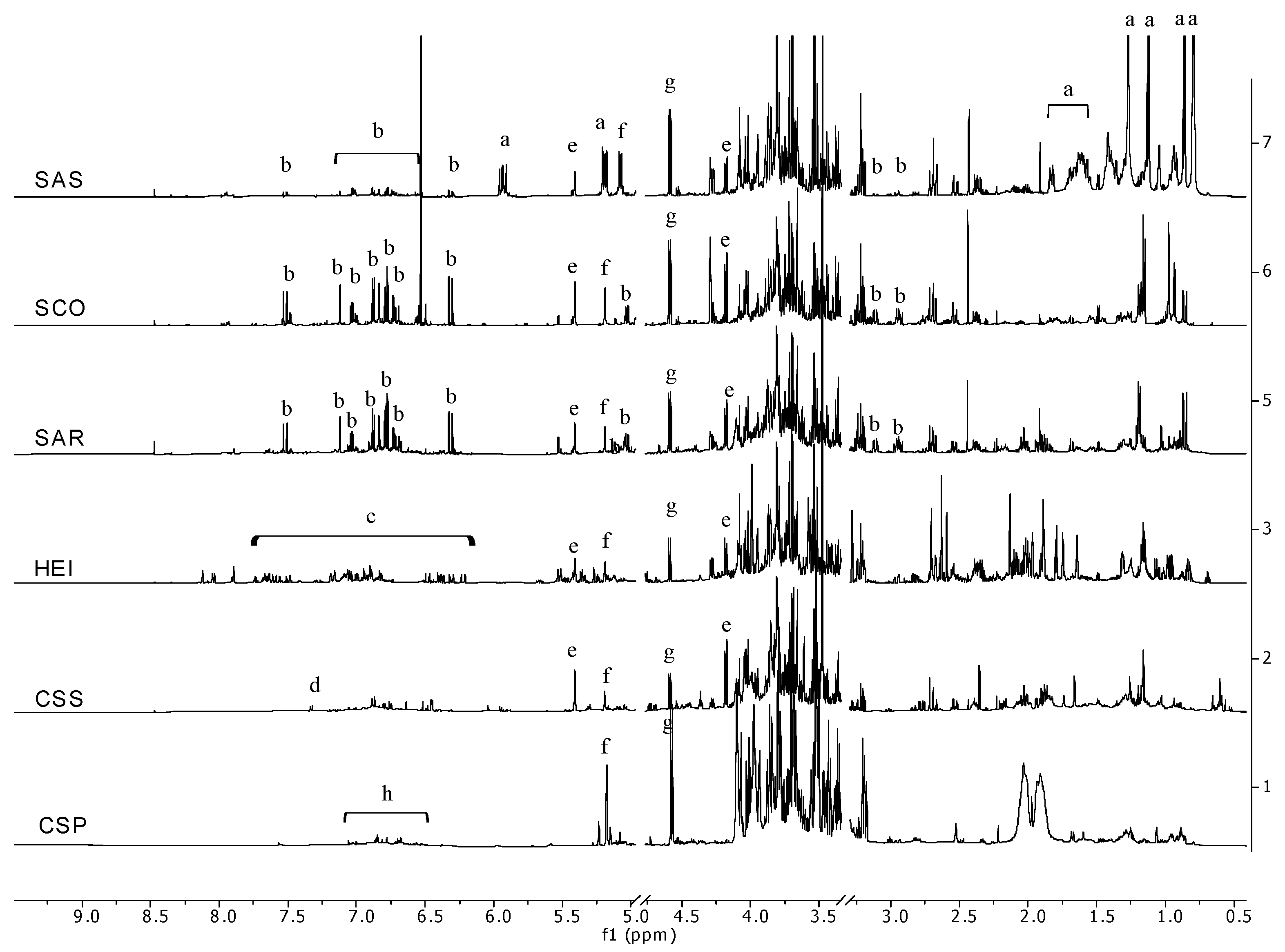
2. Results and Discussion
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material and Sample Treatment
3.3. Extracts Preparation
3.4. Total Phenolic Content and Total Flavonoid Content
3.5. Total Antioxidant Activity
3.6. Tyrosinase Inhibitory Assay
3.7. Antibacterial Activity
3.8. NMR Analysis
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stahel, W.R. The circular economy. Nat. News 2016, 531, 435. [Google Scholar] [CrossRef]
- European Commission Website. Available online: https://ec.europa.eu/environment/topics/circular-economy/first-circular-economy-action-plan_it (accessed on 1 November 2022).
- Rico, X.; Gullón, B.; Alonso, J.L.; Yáñez, R. Recovery of high value-added compounds from pineapple, melon, watermelon and pumpkin processing by-products: An overview. Food Res. Int. 2020, 132, 109086. [Google Scholar] [CrossRef]
- Pleissner, D.; Qi, Q.; Gao, C.; Rivero, C.P.; Webb, C.; Lin, C.S.K.; Venus, J. Valorization of organic residues for the production of added value chemicals: A contribution to the bio-based economy. Biochem. Eng. J. 2016, 116, 3–16. [Google Scholar] [CrossRef]
- Chinou, I. Primary and secondary metabolites and their biological activity. Chromatogr. Sci. Ser. 2008, 99, 59. [Google Scholar]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef]
- Kubo, I.; Chen, Q.X.; Nihei, K.I. Molecular design of antibrowning agents: Antioxidative tyrosinase inhibitors. Food Chem. 2003, 81, 241–247. [Google Scholar] [CrossRef]
- Chiocchio, I.; Mandrone, M.; Tomasi, P.; Marincich, L.; Poli, F. Plant Secondary Metabolites: An Opportunity for Circular Economy. Molecules 2021, 26, 495. [Google Scholar] [CrossRef]
- Jordan, N.; Boody, G.; Broussard, W.; Glover, J.D.; Keeney, D.; McCown, B.H.; McIsaac, G.; Muller, M.; Murray, H.; Neal, J.; et al. Sustainable development of the agricultural bio-economy. Science 2007, 316, 1570. [Google Scholar] [CrossRef]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef]
- World Agricultural Production Website. Available online: http://www.worldagriculturalproduction.com/crops/corn.aspx (accessed on 1 October 2022).
- Khan, D.; Mohammed, M.; Upaganlawar, A.; Upasani, C.D.; Une, H.D. Ameliorative potential of Allium cepa Lam. leaves on diabetes induced and chronic constriction injury induced neuropathic pain in experimental rats. Indian J. Pharm. Educ. Res. 2020, 54, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Milner, S.E.; Brunton, N.P.; Jones, P.W.; O’Brien, N.M.; Collins, S.G.; Maguire, A.R. Bioactivities of glycoalkaloids and their aglycones from Solanum species. J. Agric. Food Chem. 2011, 59, 3454–3484. [Google Scholar] [CrossRef]
- Chhikara, N.; Kushwaha, K.; Sharma, P.; Gat, Y.; Panghal, A. Bioactive compounds of beetroot and utilization in food processing industry: A critical review. Food Chem. 2019, 272, 192–200. [Google Scholar] [CrossRef]
- Abugri, D.A.; Tiimob, B.J.; Apalangya, V.A.; Pritchett, G.; McElhenney, W.H. Bioactive and nutritive compounds in Sorghum bicolor (Guinea corn) red leaves and their health implication. Food Chem. 2013, 138, 718–723. [Google Scholar] [CrossRef]
- Zhou, J.M.; Ibrahim, R.K. Tricin-a potential multifunctional nutraceutical. Phytochem. Rev. 2010, 9, 413–424. [Google Scholar] [CrossRef]
- Chiocchio, I.; Prata, C.; Mandrone, M.; Ricciardiello, F.; Marrazzo, P.; Tomasi, P.; Angeloni, C.; Fiorentini, D.; Malaguti, M.; Poli, F.; et al. Leaves and spiny burs of Castanea sativa from an experimental chestnut grove: Metabolomic analysis and anti-neuroinflammatory activity. Metabolites 2020, 10, 408. [Google Scholar] [CrossRef]
- Silva, V.; Falco, V.; Dias, M.I.; Barros, L.; Silva, A.; Capita, R.; Alonso-Calleja, C.; Amaral, J.S.; Igrejas, G.; Ferreira, I.C.F.R.; et al. Evaluation of the phenolic profile of Castanea sativa Mill. by-products and their antioxidant and antimicrobial activity against multiresistant bacteria. Antioxidants 2020, 9, 87. [Google Scholar] [CrossRef]
- Hassan, M.; Haq, S.M.; Rasool, A.; Fatima, S.; Ashraf, A.; Zulfajri, M.; Hanafiah, M.M. Ethnobotanical properties and traditional uses of medicinal plant Abutilon theophrasti Medik. In Medicinal and Aromatic Plants; Springer: Cham, Switzerland, 2021; pp. 271–285. [Google Scholar]
- Perović, J.; Šaponjac, V.T.; Kojić, J.; Krulj, J.; Moreno, D.A.; García-Viguera, C.; Bodroža-Solarov, M.; Ilić, N. Chicory (Cichorium intybus L.) as a food ingredient–nutritional composition, bioactivity, safety, and health claims: A review. Food Chem. 2021, 336, 127676. [Google Scholar] [CrossRef]
- Navarrete, A.; Herrero, M.; Martín, A.; Cocero, M.J.; Ibáñez, E. Valorization of solid wastes from essential oil industry. J. Food Eng. 2011, 104, 196–201. [Google Scholar] [CrossRef]
- Skendi, A.; Irakli, M.; Chatzopoulou, P.; Bouloumpasi, E.; Biliaderis, C.G. Phenolic extracts from solid wastes of the aromatic plant essential oil industry: Potential uses in food applications. Food Chem. Adv. 2022, 1, 100065. [Google Scholar] [CrossRef]
- Bordiga, M.; Travaglia, F.; Locatelli, M. Valorisation of grape pomace: An approach that is increasingly reaching its maturity—A review. Int. J. Food Sci. 2019, 54, 933–942. [Google Scholar] [CrossRef]
- Pinto, G.; De Pascale, S.; Aponte, M.; Scaloni, A.; Addeo, F.; Caira, S. Polyphenol profiling of chestnut pericarp, integument and curing water extracts to qualify these food by-products as a source of antioxidants. Molecules 2021, 26, 2335. [Google Scholar] [CrossRef]
- Kim, H.K.; Choi, Y.H.; Verpoorte, R. NMR-based metabolomic analysis of plants. Nat. Protoc. 2010, 5, 536–549. [Google Scholar] [CrossRef]
- Kim, H.K.; Verpoorte, R. Sample preparation for plant metabolomics. Phytochem. Anal. 2010, 21, 4–13. [Google Scholar] [CrossRef]
- Vasconcelos, M.C.B.M.; Bennett, R.N.; Quideau, S.; Jacquet, R.; Rosa, E.A.; Ferreira-Cardoso, J.V. Evaluating the potential of chestnut (Castanea sativa Mill.) fruit pericarp and integument as a source of tocopherols, pigments and polyphenols. Ind. Crops Prod. 2010, 31, 301–311. [Google Scholar] [CrossRef]
- Wijeratne, S.S.; Amarowicz, R.; Shahidi, F. Antioxidant activity of almonds and their by-products in food model systems. J. Am. Oil Chem. Soc. 2006, 83, 223–230. [Google Scholar] [CrossRef]
- Esfahlan, A.J.; Jamei, R.; Esfahlan, R.J. The importance of almond (Prunus amygdalus L.) and its by-products. Food Chem. 2010, 120, 349–360. [Google Scholar] [CrossRef]
- Pinelo, M.; Rubilar, M.; Sineiro, J.; Nunez, M.J. Extraction of antioxidant phenolics from almond hulls (Prunus amygdalus) and pine sawdust (Pinus pinaster). Food Chem. 2004, 85, 267–273. [Google Scholar] [CrossRef]
- Önder, S.; Tonguç, M.; Erbaş, S.; Önder, D.; Mutlucan, M. Investigation of phenological, primary and secondary metabolites changes during flower developmental of Rosa damascena. Plant Physiol. Biochem. 2022, 192, 20–34. [Google Scholar] [CrossRef]
- Flórez-Fernández, N.; Torres, M.D.; Gómez, S.; Couso, S.; Domínguez, H. Potential of chestnut wastes for cosmetics and pharmaceutical applications. Waste Biomass Valorization 2020, 11, 4721–4730. [Google Scholar] [CrossRef]
- Akinyede, K.A.; Cupido, C.N.; Hughes, G.D.; Oguntibeju, O.O.; Ekpo, O.E. Medicinal properties and In vitro biological activities of selected Helichrysum species from South Africa: A review. Plants 2021, 10, 1566. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, S.; Moreira, E.; Grosso, C.; Andrade, P.B.; Valentão, P.; Romano, A. Phenolic profile, antioxidant activity and enzyme inhibitory activities of extracts from aromatic plants used in Mediterranean diet. J. Food Sci. Technol. 2017, 54, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Mollica, A.; Aktumsek, A.; Picot, C.M.N.; Mahomoodally, M.F. In vitro and in silico insights of Cupressus sempervirens, Artemisia absinthium and Lippia triphylla: Bridging traditional knowledge and scientific validation. Eur. J. Integr. Med. 2017, 12, 135–141. [Google Scholar] [CrossRef]
- El Aanachi, S.; Gali, L.; Rammali, S.; Bensouici, C.; Aassila, H.; Dari, K. In vitro study of the antioxidant, photoprotective, anti-tyrosinase, and anti-urease effects of methanolic extracts from leaves of six Moroccan Lamiaceae. J. Food Meas. Charact. 2021, 15, 1785–1795. [Google Scholar] [CrossRef]
- Moreno, S.; Scheyer, T.; Romano, C.S.; Vojnov, A.A. Antioxidant and antimicrobial activities of rosemary extracts linked to their polyphenol composition. Free Radic. Res. 2006, 40, 223–231. [Google Scholar] [CrossRef]
- Kang, H.S.; Kim, H.R.; Byun, D.S.; Park, H.J.; Choi, J.S. Rosmarinic acid as a tyrosinase inhibitors from Salvia miltiorrhiza. Nat. Prod. Sci. 2004, 10, 80–84. [Google Scholar]
- Cappadone, C.; Mandrone, M.; Chiocchio, I.; Sanna, C.; Malucelli, E.; Bassi, V.; Picone, G.; Poli, F. Antitumor potential and phytochemical profile of plants from Sardinia (Italy), a hotspot for biodiversity in the mediterranean basin. Plants 2020, 9, 26. [Google Scholar] [CrossRef]
- Chiocchio, I.; Mandrone, M.; Sanna, C.; Maxia, A.; Tacchini, M.; Poli, F.J.I.C. Screening of a hundred plant extracts as tyrosinase and elastase inhibitors, two enzymatic targets of cosmetic interest. Ind. Crops Prod. 2018, 122, 498–505. [Google Scholar] [CrossRef]
- Marrelli, M.; Russo, N.; Chiocchio, I.; Statti, G.; Poli, F.; Conforti, F. Potential use in the treatment of inflammatory disorders and obesity of selected wild edible plants from Calabria region (Southern Italy). S. Afr. J. Bot. 2020, 128, 304–311. [Google Scholar] [CrossRef]
- Mandrone, M.; Marincich, L.; Chiocchio, I.; Petroli, A.; Gođevac, D.; Maresca, I.; Poli, F. NMR-based metabolomics for frauds detection and quality control of oregano samples. Food Control 2021, 127, 108141. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Common Name | Plant Part | Type of Waste/By-Product | Sample Tag | Voucher Number |
|---|---|---|---|---|---|
| Abutilon theophrasti Medik. | Velvetleaf | Aerial parts | Pest plant | Ath | WST25 |
| Achillea millefolium L. | Common yarrow | Aerial parts | Solid waste from distillation | Acm | WST35 |
| Allium cepa L. | Onion | Dry aerial parts | Agricultural residue | Ace | WST1 |
| Artemisia absinthium L. | Wormwood | Aerial parts | Solid waste from distillation | Ara | WST36 |
| Beta vulgaris L. | Sugar beet | Aerial parts | Agricultural residue | Bvu | WST10 |
| Camelina sativa (L.) Crantz | Camelina | Dry aerial parts | Agricultural residue | Csa | WST2 |
| Castanea sativa Mill. | Chestnut | Pericarp | Food Industry by-product | Csp | WST16 |
| Spiny burs | Agricultural residue | Csr | WST3 | ||
| Cicer arietinum L. | Chickpea | Aerial parts | Agricultural residue | Car | WST11 |
| Cichorium intybus L. | Chicory | Apical flowering aerial parts | Pest plant | Cia | WST22 |
| Basal flowering aerial parts | Pest plant | Cib | WST23 | ||
| Cucurbita pepo L. | Courgette | Leaves | Agricultural residue | Cpe | WST12 |
| Aerial parts | Agricultural residue | Cpi | WST15 | ||
| Cupressus sempervirens L. | Cupressus | Leaves | Solid waste from distillation | Css | WST26 |
| Echinochloa crus-galli (L.) P.Beauv. | Cockspur | Flowering aerial parts | Pest plant | Ecg | WST24 |
| Erigeron canadensis L. | Horseweed | Aerial parts | Pest plant | Eca | WST21 |
| Helianthus annuus L. | Sunflower | Leaves | Agricultural residue | Han | WST4 |
| Helichrysum italicum (Roth) G. Don | Curry plant | Aerial parts | Solid waste from distillation | Hei | WST27 |
| Laurus nobilis L. | Laurel | Leaves | Solid waste from distillation | Lan | WST28 |
| Lavandula angustifolia Mill. | Lavender | Aerial parts | Solid waste from distillation | Laa | WST37 |
| Melissa officinalis L. | Lemon balm | Aerial parts | Solid waste from distillation | Meo | WST29 |
| Origanum vulgare L. | Oregano | Stems | Food Industry by-product | Orv | WST19 |
| Phaseolus vulgaris L. | Bean | Husks | Food Industry by-product | Pvb | WST18 |
| Aerial parts | Agricultural residue | Pvu | WST13 | ||
| Prunus amygdalus Batsch | Almond | Exocarp and mesocarp | Food Industry by-product | Pam | WST17 |
| Rosa damascena | Damask rose | Buds | Solid waste from distillation | Rod | WST30 |
| Salvia officinalis L. | Sage | Aerial parts | Solid waste from distillation | Sco | WST33 |
| Salvia rosmarinus Schleid. | Rosemary | Aerial parts | Solid waste from distillation | Sar | WST31 |
| Salvia sclarea L. | Clary sage | Aerial parts | Solid waste from distillation | Sas | WST32 |
| Solanum lycopersicum L. | Tomato | Basal leaves | Agricultural residue | Sly | WST14 |
| Solanum tuberosum L. | Potato | Leaves | Agricultural residue | Stu | WST8 |
| Sorghum bicolor (L.) Moench | Sorghum | Leaves | Agricultural residue | Sbl | WST6 |
| Roots | Agricultural residue | Sbr | WST7 | ||
| Stems | Agricultural residue | Sbf | WST5 | ||
| Thymus vulgaris L. | Common thyme | Aerial parts | Solid waste from distillation | Tvu | WST34 |
| Triticum aestivum L. | Wheat | Dry aerial parts | Agricultural residue | Tae | WST9 |
| Vitis vinifera L. | Grape | Pomace | Food Industry by-product | Vvi | WST20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiocchio, I.; Mandrone, M.; Tacchini, M.; Guerrini, A.; Poli, F. Phytochemical Profile and In Vitro Bioactivities of Plant-Based By-Products in View of a Potential Reuse and Valorization. Plants 2023, 12, 795. https://doi.org/10.3390/plants12040795
Chiocchio I, Mandrone M, Tacchini M, Guerrini A, Poli F. Phytochemical Profile and In Vitro Bioactivities of Plant-Based By-Products in View of a Potential Reuse and Valorization. Plants. 2023; 12(4):795. https://doi.org/10.3390/plants12040795
Chicago/Turabian StyleChiocchio, Ilaria, Manuela Mandrone, Massimo Tacchini, Alessandra Guerrini, and Ferruccio Poli. 2023. "Phytochemical Profile and In Vitro Bioactivities of Plant-Based By-Products in View of a Potential Reuse and Valorization" Plants 12, no. 4: 795. https://doi.org/10.3390/plants12040795
APA StyleChiocchio, I., Mandrone, M., Tacchini, M., Guerrini, A., & Poli, F. (2023). Phytochemical Profile and In Vitro Bioactivities of Plant-Based By-Products in View of a Potential Reuse and Valorization. Plants, 12(4), 795. https://doi.org/10.3390/plants12040795