
Genetic Diversity Analysis of Brassica Yellows Virus Causing Aberrant Color Symptoms in Oilseed Rape

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. BrYV Was the Most Important Viral Pathogen in Oilseed Rape in Yangzhong, Jiangsu
2.2. BrYV Infection Causes Different Symptoms in the Field
2.3. The Relationship between Leaf Color and the Occurrence of the Virus
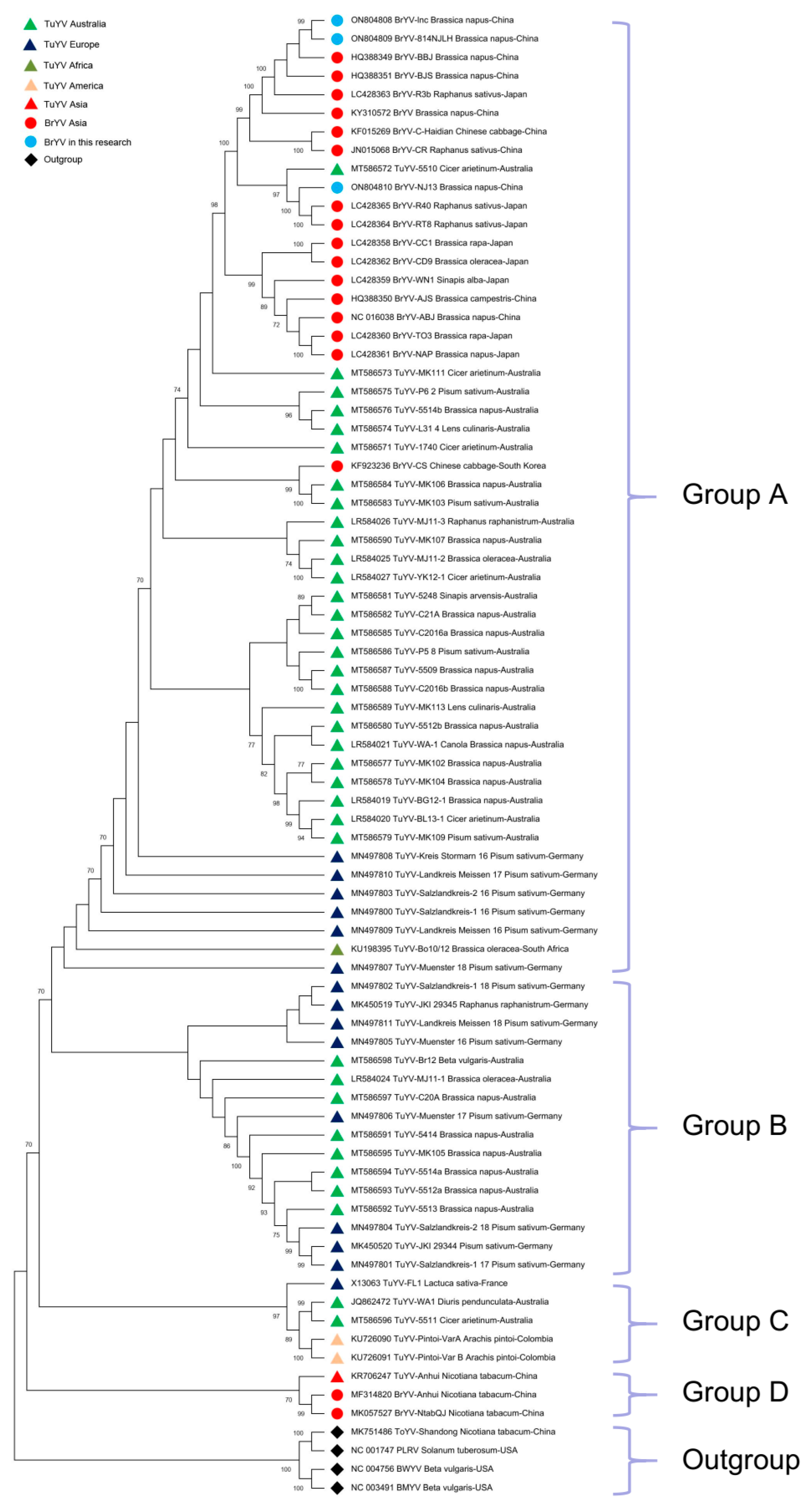
2.4. Phylogenetic Analysis of BrYV and TuYV Sequences
2.5. Pairwise Analysis of the BrYV Proteins
2.6. Recombination Analysis
3. Discussion
4. Materials and Methods
4.1. Plants and Sampling
4.2. RNA-Seq and Data Analysis
4.3. Phylogenetic Tree Construction, Recombinant Analysis
4.4. Virus Detection and Cloning
4.5. Leaf Image Capture, Color Quantification and Correlation Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stevens, M.; McGrann, G.R.D.; Clark, B. Turnip yellows virus (syn Beet western yellows virus): An emerging threat to European oilseed rape production? HGCA Res. Rev. 2008, 69, 1–36. [Google Scholar]
- Jones, R.A.C.; Coutts, B.A.; Hawkes, J. Yield-limiting potential of Beet western yellows virus in Brassica napus. Aust. J. Agric. Res. 2007, 58, 788–801. [Google Scholar] [CrossRef]
- Sõmera, M.; Fargette, D.; Hébrard, E.; Sarmiento, C. ICTV Report Consortium. ICTV Virus Taxonomy Profile: Solemoviridae 2021. J. Gen. Virol. 2021, 102, 001707. [Google Scholar] [CrossRef]
- Hackenberg, D.; Asare-Bediako, E.; Baker, A.; Walley, P.; Jenner, C.; Greer, S.; Bramham, L.; Batley, J.; Edwards, D.; Delourme, R.; et al. Identification and QTL mapping of resistance to Turnip yellows virus (TuYV) in oilseed rape, Brassica napus. Theor. Appl. Genet. 2020, 133, 383–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.Y.; Peng, Y.M.; Xiang, H.Y.; Wang, Y.; Li, D.W.; Yu, J.L.; Han, C.G. Incidence and prevalence levels of three aphid-transmitted viruses in crucifer crops in China. J. Integr. Agric. 2022, 21, 774–780. [Google Scholar] [CrossRef]
- Kamitani, M.; Nagano, A.J.; Honjo, M.N.; Kudoh, H. RNA-Seq reveals virus-virus and virus-plant interactions in nature. FEMS Microbiol. Ecol. 2016, 92, fiw176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.; Yoo, R.H.; Igori, D.; Zhao, F.; Kim, K.H.; Moon, J.S. Genome sequence of a recombinant brassica yellows virus infecting Chinese cabbage. Arch. Virol. 2015, 160, 597–600. [Google Scholar] [CrossRef]
- Xiang, H.Y.; Dong, S.W.; Shang, Q.X.; Zhou, C.J.; Li, D.W.; Yu, J.L.; Han, C.G. Molecular characterization of two genotypes of a new polerovirus infecting brassicas in China. Arch. Virol. 2011, 156, 2251–2255. [Google Scholar] [CrossRef]
- Gray, S.; Cilia, M.; Ghanim, M. Circulative, “nonpropagative” virus transmission: An orchestra of virus-, insect-, and plant-derived instruments. Adv. Virus Res. 2014, 89, 141–199. [Google Scholar]
- Zhang, X.Y.; Xiang, H.Y.; Zhou, C.J.; Li, D.W.; Yu, J.L.; Han, C.G. Complete genome sequence analysis identifies a new genotype of brassica yellows virus that infects cabbage and radish in China. Arch. Virol. 2014, 159, 2177–2180. [Google Scholar] [CrossRef]
- Yoshida, N.; Tamada, T. Host range and molecular analyses of Beet leaf yellowing virus, Beet western yellows virus-JP and Brassica yellows virus in Japan. Plant Pathol. 2019, 68, 1045–1058. [Google Scholar] [CrossRef]
- Taliansky, M.; Mayo, M.A.; Barker, H. Potato leafroll virus: A classic pathogen shows some new tricks. Mol. Plant Pathol. 2003, 4, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Derrien, B.; Baumberger, N.; Schepetilnikov, M.; Viotti, C.; De Cillia, J.; Ziegler-Graff, V.; Isono, E.; Schumacher, K.; Genschik, P. Degradation of the antiviral component ARGONAUTE1 by the autophagy pathway. Proc. Natl. Acad. Sci. USA 2012, 109, 15942–15946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Y.; Sun, Q.; Zhao, T.Y.; Xiang, H.Y.; Zhang, X.Y.; Wu, Z.Y.; Zhou, C.J.; Zhang, X.; Wang, Y.; Zhang, Y.L.; et al. Interaction between brassica yellows virus silencing suppressor P0 and plant SKP1 facilitates stability of P0 in vivo against degradation by proteasome and autophagy pathways. New Phytol. 2019, 222, 1458–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prüfer, D.; Kawchuk, L.; Monecke, M.; Nowok, S.; Fischer, R.; Rohde, W. Immunological analysis of potato leafroll luteovirus (PLRV) P1 expression identifies a 25kDa RNA-binding protein derived via P1 processing. Nucleic Acids Res. 1999, 27, 421–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Wilk, F.; Verbeek, M.; Dullemans, A.M.; van der Heuvel, J.F.J.M. The genome-linked protein of Potato leafroll virus is located downstream of the putative protease domain of the ORF1 product. Virology 1997, 234, 300–303. [Google Scholar] [CrossRef] [Green Version]
- Smirnova, E.; Firth, A.E.; Allen Miller, W.; Scheidecker, D.; Brault, V.; Reinbold, C.; Rakotondrafara, A.M.; Chung, B.Y.W.; Ziegler-Graff, V. Discovery of a small non-AUG-initiated ORF in poleroviruses and luteoviruses that is required for long-distance movement. PLoS Pathog. 2015, 11, e1004868. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, J.; Stussi-Garaud, C.; Tacke, E.; Prüfer, D.; Rohde, W.; Rohfritsch, O. In situ localization of the putative movement protein (pr17) from potato leafroll lutevirus (PLRV) in infected and transgenic potato plants. Virology 1997, 235, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y.; Zhao, T.Y.; Li, Y.Y.; Xiang, H.Y.; Dong, S.W.; Zhang, Z.Y.; Wang, Y.; Li, D.W.; Yu, J.L.; Han, C.G. The conserved proline18 in the Polerovirus P3a is important for brassica yellows virus systemic infection. Front. Microbiol. 2018, 9, 613. [Google Scholar] [CrossRef]
- Xu, Y.; Ju, H.J.; Deblasio, S.; Carino, E.J.; Johnson, R.; MacCoss, M.J.; Heck, M.; Allen Miller, W.; Gray, S.M. A stem-loop structure in Potato leafroll virus open reading frame 5 (ORF5) is essential for readthrough translation of the coat protein ORF stop codon 700 bases upstream. J. Virol. 2018, 92, e01544-17. [Google Scholar] [CrossRef] [Green Version]
- Byrne, M.J.; Steele, J.F.C.; Hesketh, E.L.; Walden, M.; Thompson, R.F.; Lomonossoff, G.P.; Ranson, N.A. Combining transient expression and Cryo-EM to obtain high-resolution structures of Luteovirid particles. Structure 2019, 27, 1761–1770. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.R.; Wang, Y.; Zhao, H.H.; Zhang, X.Y.; Wang, X.B.; Li, D.W.; Yu, J.L.; Han, C.G. Brassica yellows virus’ movement protein upregulates anthocyanin accumulation, leading to the development of purple leaf symptoms on Arabidopsis thaliana. Sci. Rep. 2018, 8, 16273. [Google Scholar] [CrossRef] [Green Version]
- Umar, M.; Farooq, T.; Tegg, R.S.; Thangavel, T.; Wilson, C.R. Genomic characterisation of an isolate of Brassica yellows virus associated with brassica weed in Tasmania. Plants 2022, 11, 884. [Google Scholar] [CrossRef] [PubMed]
- Filardo, F.; Nancarrow, N.; Kehoe, M.; McTaggart, A.R.; Congdon, B.; Kumari, S.; Aftab, M.; Trębicki, P.; Rodoni, B.; Thomas, J.; et al. Genetic diversity and recombination between turnip yellows virus strains in Australia. Arch. Virol. 2021, 166, 813–829. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, A.D.; Menzel, W.; Lesemann, D.E.; Varrelmann, M.; Vetten, H.J. Chickpea chlorotic stunt virus: A new polerovirus infecting cool-season food legumes in Ethiopia. Virology 2006, 96, 437–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buxton-Kirk, A.; Adams, I.; Frew, L.; Ward, R.; Kelly, M.; Forde, S.; Skelton, A.; Harju, V.; Baucas, N.S.; Bas-Ilan, M.A.G.; et al. First report of Turnip yellows virus in cabbage in the Philippines. New Dis. Rep. 2021, 44, 1. [Google Scholar] [CrossRef]
- Gaafar, Y.Z.A.; Ziebell, H. Two divergent isolates of turnip yellows virus from pea and rapeseed and first report of turnip yellows virus-associated RNA in Germany. Microbiol. Resour. Announc. 2019, 8, e00214-19. [Google Scholar] [CrossRef] [Green Version]
- Milošević, D.; Ignjatov, M.; Nikolić, Z.; Stanković, I.; Bulajić, A.; Marjanović-Jeromela, A.; Krstić, B. The presence of Turnip yellows virus in oilseed rape (Brassica napus L.) in Serbia. Pestic. Phytomed. 2016, 31, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Slavíková, L.; Ibrahim, E.; Alquicer, G.; Tomašechová, J.; Šoltys, K.; Glasa, M.; Kundu, J.K. Weed hosts represent an important reservoir of turnip yellows virus and a possible source of virus introduction into oilseed rape crop. Viruses 2022, 14, 2511. [Google Scholar] [CrossRef]
- Nancarrow, N.; Aftab, M.; Hollaway, G.; Rodoni, B.; Trębicki, P. Symptomless turnip yellows virus infection causes grain yield loss in lentil and field pea: A three-year field study in south-eastern Australia. Front. Plant Sci. 2022, 13, 1049905. [Google Scholar] [CrossRef] [PubMed]
- Umar, M.; Tegg, R.S.; Farooq, T.; Thangavel, T.; Wilson, C.R. Abundance of Poleroviruses within Tasmanian pea cops and surrounding weeds, and the genetic diversity of TuYV isolates found. Viruses 2022, 14, 1690. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.D. The investigation of plant diseases. In Research Methods of Plant Pathology; Zhang, H.G., Ed.; China Agriculture Press: Beijing, China, 1998; pp. 12–13. [Google Scholar]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Xia, R. Tbtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Peng, Y.M.; Wang, Y.; Zhang, Z.Y.; Li, D.W.; Yu, J.L.; Han, C.G. Simultaneous detection and differentiation of three genotypes of Brassica yellows virus by multiplex reverse transcription-polymerase chain reaction. Virol. J. 2016, 13, 189. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.Y.; Zhang, C.W.; Hou, X.L.; Sanfaçon, H.; Wang, A.M. The SNARE protein Syp71 is essential for Turnip mosaic virus infection by mediating fusion of virus-induced vesicles with chloroplasts. PLoS Pathog. 2013, 9, e1003378. [Google Scholar] [CrossRef] [Green Version]
- Shang, Q.X.; Xiang, H.Y.; Han, C.G.; Li, D.W.; Yu, J.L. Partial sequence analysis of two isolates of Cucurbit aphid-borne yellows virus from Hubei and Yunnan in China. Acta Phytopathol. Sin. 2008, 38, 64–68. [Google Scholar]
- Wang, Y.; Wang, D.J.; Shi, P.H.; Omasa, K. Estimating rice chlorophyll content and leaf nitrogen concentration with a digital still color camera under natural light. Plant Methods 2014, 10, 36. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fields | Total Samples | BrYV-A | BrYV-B | AB Mix | BrYV | BrYV Incidence (%) | TuYV | TuYV Incidence (%) | TuMV | TuMV Incidence (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| A | 20 | 3 | 0 | 1 | 4 | 20.00 | 0 | 0.00 | 11 | 55.00 |
| B | 24 | 2 | 1 | 2 | 5 | 20.83 | 1 | 4.17 | 8 | 33.33 |
| C | 23 | 1 | 2 | 3 | 6 | 26.09 | 0 | 0.00 | 3 | 13.04 |
| D | 19 | 0 | 4 | 5 | 9 | 47.37 | 0 | 0.00 | 3 | 15.79 |
| E | 17 | 7 | 1 | 1 | 9 | 52.94 | 5 | 29.41 | 0 | 0.00 |
| Total | 103 | 13 | 8 | 12 | 33 | 32.04 | 6 | 5.83 | 25 | 24.27 |
| P0 (%) | P1 (%) | P2 (%) | P3 (%) | P3a (%) | P4 (%) | P5 (%) | |
|---|---|---|---|---|---|---|---|
| BrVY | 91.97~100 | 88.3~100 | 97.3~100 | 94.06~100 | 82.22~100 | 86.89~100 | 51.55~100 |
| Br to Tu-FL1 | 86.75~89.96 | 89.31~91.6 | 96.22~97.84 | 95.05~98.02 | 86.67~97.78 | 90.16~96.57 | 46.84~77.48 |
| 814NJLH to Br | 94.38~100 | 89.07~99.67 | 97.84~100 | 96.04~100 | 84.44~100 | 89.62~100 | 57.11~100 |
| NJ13 to Br | 93.17~100 | 88.65~99.67 | 97.84~100 | 96.04~100 | 84.44~100 | 89.62~100 | 56.91~99.75 |
| Events | Recombinant | Major Parent | Minor Parent | Recombination Site | p-Values of Different Detection Method | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Begin | End | R | G | B | M | C | S | T | ||||
| 1 | TuYV-C20A | TuYV-Br12 | TuYV-MJ11-1 | 1492 | 2663 | 1.99 × 10−19 | 1.70 × 10−15 | 2.00 × 10−18 | 1.15 × 10−14 | 7.92 × 10−16 | 6.92 × 10−16 | 3.89 × 10−15 |
| 2 | TuYV-C2016a | TuYV-YK12-1 | TuYV-5509 | 2668 | 5654 | 6.43 × 10−47 | 2.37 × 10−43 | 3.23 × 10−48 | 4.85 × 10−19 | 4.97 × 10−22 | 7.62 × 10−37 | 6.64 × 10−51 |
| 3 | TuYV-MK111 | TuYV-MK107 | BrYV-NJ13 | 3368 | 5508 | 5.01 × 10−60 | 2.81 × 10−59 | 1.67 × 10−63 | 2.72 × 10−36 | 2.75 × 10−30 | 1.12 × 10−53 | 1.27 × 10−13 |
| 4 | TuYV-MJ11-2 | TuYV-MJ11-3 | TuYV-JKI | 3316 | 4277 | 1.08 × 10−49 | 2.58 × 10−50 | 5.99 × 10−47 | 2.74 × 10−20 | 1.83 × 10−20 | 2.60 × 10−21 | 5.11 × 10−13 |
| 5 | TuYV-1740 | TuYV-MK107 | TuYV-5510 | 3496 | 5624 | 9.20 × 10−63 | 2.12 × 10−56 | 6.52 × 10−64 | 5.45 × 10−33 | 5.02 × 10−22 | 6.61 × 10−58 | 1.27 × 10−13 |
| 6 | TuYV-5514b | TuYV-MK113 | BrYV-CR | 3842 | 4872 | 3.88 × 10−70 | 4.34 × 10−68 | 5.92 × 10−68 | 3.37 × 10−36 | 2.90 × 10−36 | 8.55 × 10−41 | 1.48 × 10−13 |
| 7 | TuYV-P6_2 | TuYV-Salzlandkreis-1 | BrYV-R3b | 3878 | 5628 | 1.33 × 10−101 | 6.41 × 10−76 | 3.33 × 10−94 | 9.24 × 10−44 | 2.92 × 10−13 | 4.18 × 10−69 | 9.53 × 10−56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Q.; Li, W.; Zhou, X.; Sun, C.; Hou, Y.; Hu, M.; Fu, S.; Zhang, J.; Kundu, J.K.; Lei, L. Genetic Diversity Analysis of Brassica Yellows Virus Causing Aberrant Color Symptoms in Oilseed Rape. Plants 2023, 12, 1008. https://doi.org/10.3390/plants12051008
Peng Q, Li W, Zhou X, Sun C, Hou Y, Hu M, Fu S, Zhang J, Kundu JK, Lei L. Genetic Diversity Analysis of Brassica Yellows Virus Causing Aberrant Color Symptoms in Oilseed Rape. Plants. 2023; 12(5):1008. https://doi.org/10.3390/plants12051008
Chicago/Turabian StylePeng, Qi, Wei Li, Xiaoying Zhou, Chengming Sun, Yan Hou, Maolong Hu, Sanxiong Fu, Jiefu Zhang, Jiban Kumar Kundu, and Lei Lei. 2023. "Genetic Diversity Analysis of Brassica Yellows Virus Causing Aberrant Color Symptoms in Oilseed Rape" Plants 12, no. 5: 1008. https://doi.org/10.3390/plants12051008
APA StylePeng, Q., Li, W., Zhou, X., Sun, C., Hou, Y., Hu, M., Fu, S., Zhang, J., Kundu, J. K., & Lei, L. (2023). Genetic Diversity Analysis of Brassica Yellows Virus Causing Aberrant Color Symptoms in Oilseed Rape. Plants, 12(5), 1008. https://doi.org/10.3390/plants12051008