

In Vivo Antiphytoviral and Aphid Repellency Activity of Essential Oils and Hydrosols from Mentha suaveolens and Foeniculum vulgare to Control Zucchini Yellow Mosaic Virus and Its Vector Aphis gossypii
 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Chemical Composition of Plant Extracts
2.2. Effect of Plant Extracts on Virus Titer in ZYMV-Infected Plants
2.2.1. Treatments Applied at the Same Time as Inoculation
2.2.2. Treatments Applied after Inoculation
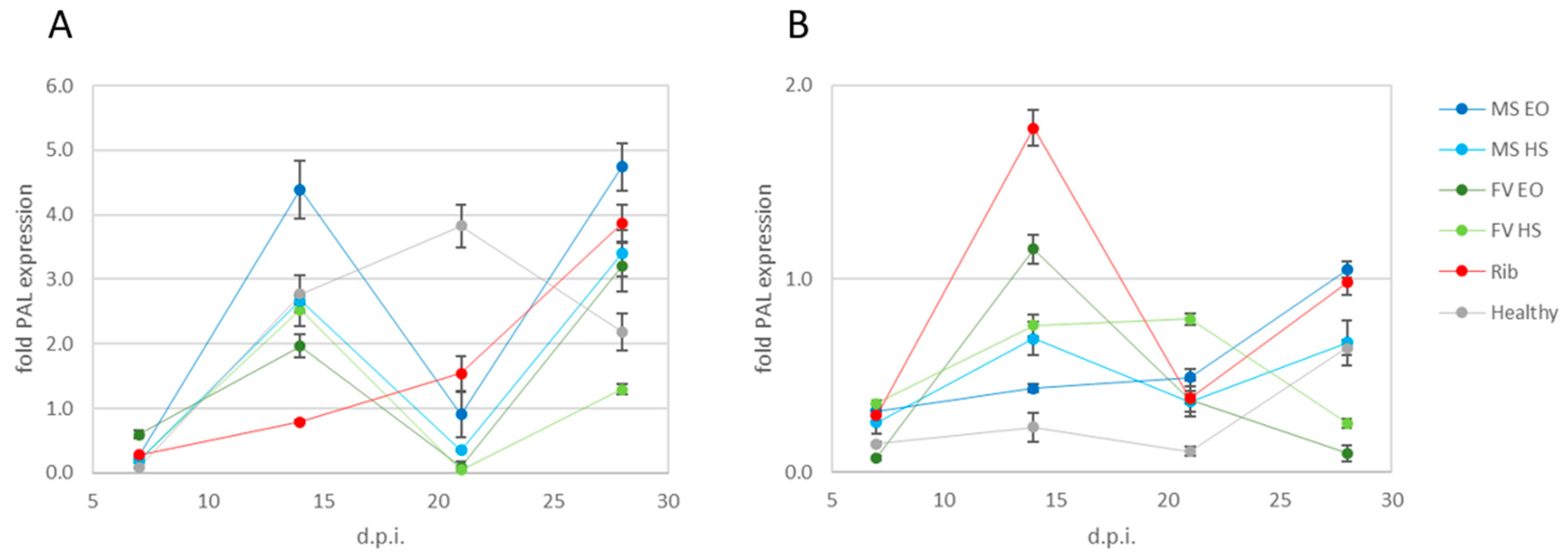
2.3. Effect of Plant Extracts on Phenylalanine Ammonia Lyase Gene Expression
2.3.1. Treatments Applied at the Same Time as Inoculation
2.3.2. Treatments Applied after Inoculation
2.4. Effect of Plant Extracts on Virus Vector Choice and Survival
2.4.1. Repellency
2.4.2. Toxicity and Fecundity
3. Discussion
4. Materials and Methods
4.1. Aromatic Plants
4.2. Essential Oil and Hydrosol Production and Characterization
4.2.1. Essential Oils
4.2.2. Hydrosol Preparation
4.2.3. Gas Chromatography–Mass Spectrometry Analysis
4.3. Experimental Plant Material
4.3.1. Plant Host
4.3.2. Virus Inoculum
4.4. Experimental Trials
4.4.1. Treatments Applied at the Same Time as Inoculation
4.4.2. Treatments Applied after Inoculation
4.4.3. RNA Extraction and Real-Time RT-PCR
4.4.4. Insect Bioassays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Baser, K.H.C.; Buchbauer, G. (Eds.) Handbook of Essential Oils; CRC Press: Boca Raton, FL, USA, 2015; ISBN 9780429155666. [Google Scholar]
- Božović, M.; Navarra, A.; Garzoli, S.; Pepi, F.; Ragno, R. Esential Oils Extraction: A 24-Hour Steam Distillation Systematic Methodology. Nat. Prod. Res. 2017, 31, 2387–2396. [Google Scholar] [CrossRef] [PubMed]
- Barton, P.; Hughes, R.E.; Hussein, M.M. Supercritical Carbon Dioxide Extraction of Peppermint and Spearmint. J. Supercrit. Fluids 1992, 5, 157–162. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological Effects of Essential Oils—A Review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Božović, M.; Garzoli, S.; Baldisserotto, A.; Romagnoli, C.; Pepi, F.; Cesa, S.; Vertuani, S.; Manfredini, S.; Ragno, R. Melissa Officinalis L. Subsp. Altissima (Sibth. & Sm.) Arcang. Essential Oil: Chemical Composition and Preliminary Antimicrobial Investigation of Samples Obtained at Different Harvesting Periods and by Fractionated Extractions. Ind. Crops Prod. 2018, 117, 317–321. [Google Scholar] [CrossRef]
- Sabatino, M.; Fabiani, M.; Božović, M.; Garzoli, S.; Antonini, L.; Marcocci, M.E.; Palamara, A.T.; de Chiara, G.; Ragno, R. Experimental Data Based Machine Learning Classification Models with Predictive Ability to Select in Vitro Active Antiviral and Non-Toxic Essential Oils. Molecules 2020, 25, 2452. [Google Scholar] [CrossRef]
- Papa, R.; Garzoli, S.; Vrenna, G.; Sabatino, M.; Sapienza, F.; Relucenti, M.; Donfrancesco, O.; Fiscarelli, E.; Artini, M.; Selan, L.; et al. Essential Oils Biofilm Modulation Activity, Chemical and Machine Learning Analysis—Application on Staphylococcus Aureus Isolates from Cystic Fibrosis Patients. Int. J. Mol. Sci. 2020, 21, 9258. [Google Scholar] [CrossRef]
- Barragán-Ocaña, A.; Silva-Borjas, P.; Olmos-Peña, S. Scientific and Technological Trajectories for Sustainable Agricultural Solutions. In Biopesticides; Elsevier: Amsterdam, The Netherlands, 2022; pp. 93–105. [Google Scholar]
- Labadie, C.; Cerutti, C.; Carlin, F. Fate and Control of Pathogenic and Spoilage Micro-Organisms in Orange Blossom (Citrus Aurantium) and Rose Flower (Rosa Centifolia) Hydrosols. J. Appl. Microbiol. 2016, 121, 1568–1579. [Google Scholar] [CrossRef]
- Xie, L.H.; Lin, Q.Y.; Wu, Z.J. Plant. Virus: Virology and Molecular Biology; Science Press: Bejing, China, 2009. [Google Scholar]
- Rubio, L.; Galipienso, L.; Ferriol, I. Detection of Plant Viruses and Disease Management: Relevance of Genetic Diversity and Evolution. Front. Plant Sci. 2020, 11, 1092. [Google Scholar] [CrossRef]
- Golino, D.A.; Fuchs, M.; Rwahnih, M.A.; Farrar, K.; Schmidt, A.; Martelli, G.P. Regulatory Aspects of Grape Viruses and Virus Diseases: Certification, Quarantine, and Harmonization. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 581–598. [Google Scholar]
- Fereres, A.; Raccah, B. Plant Virus Transmission by Insects; eLS. John Wiley & Sons: Chichester, UK, 2015. [Google Scholar]
- Bishop, C.D. Antiviral Activity of the Essential Oil of Melaleuca Alternifolia (Maiden Amp; Betche) Cheel (Tea Tree) Against Tobacco Mosaic Virus. J. Essent. Oil Res. 1995, 7, 641–644. [Google Scholar] [CrossRef]
- Dunkić, V.; Bezić, N.; Vuko, E.; Cukrov, D. Antiphytoviral Activity of Satureja Montana L. Ssp. Variegata (Host) P. W. Ball Essential Oil and Phenol Compounds on CMV and TMV. Molecules 2010, 15, 6713–6721. [Google Scholar] [CrossRef] [Green Version]
- Lu, M. In Vitro and In Vivo Anti-Tobacco Mosaic Virus Activities of Essential Oils and Individual Compounds. J. Microbiol. Biotechnol. 2013, 23, 771–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezić, N.; Vuko, E.; Dunkić, V.; Ruščić, M.; Blažević, I.; Burčul, F. Antiphytoviral Activity of Sesquiterpene-Rich Essential Oils from Four Croatian Teucrium Species. Molecules 2011, 16, 8119–8129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Shafi, S. Preliminary Studies on Antibacterial and Antiviral Activities of Five Medicinal Plants. J. Plant Pathol. Microbiol. 2013, 4, 190. [Google Scholar] [CrossRef] [Green Version]
- Vuko, E.; Rusak, G.; Dunkić, V.; Kremer, D.; Kosalec, I.; Rađa, B.; Bezić, N. Inhibition of Satellite RNA Associated Cucumber Mosaic Virus Infection by Essential Oil of Micromeria Croatica (Pers.) Schott. Molecules 2019, 24, 1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taglienti, A.; Donati, L.; Ferretti, L.; Tomassoli, L.; Sapienza, F.; Sabatino, M.; di Massimo, G.; Fiorentino, S.; Vecchiarelli, V.; Nota, P.; et al. In Vivo Antiphytoviral Activity of Essential Oils and Hydrosols From Origanum Vulgare, Thymus Vulgaris, and Rosmarinus Officinalis to Control Zucchini Yellow Mosaic Virus and Tomato Leaf Curl New Delhi Virus in Cucurbita pepo L. Front. Microbiol. 2022, 13, 840893. [Google Scholar] [CrossRef]
- D’Amato, S.; Serio, A.; López, C.C.; Paparella, A. Hydrosols: Biological Activity and Potential as Antimicrobials for Food Applications. Food Control 2018, 86, 126–137. [Google Scholar] [CrossRef]
- Saǧdıç, O.; Özcan, M. Antibacterial Activity of Turkish Spice Hydrosols. Food Control 2003, 14, 141–143. [Google Scholar] [CrossRef]
- Tornuk, F.; Cankurt, H.; Ozturk, I.; Sagdic, O.; Bayram, O.; Yetim, H. Efficacy of Various Plant Hydrosols as Natural Food Sanitizers in Reducing Escherichia Coli O157:H7 and Salmonella Typhimurium on Fresh Cut Carrots and Apples. Int. J. Food Microbiol. 2011, 148, 30–35. [Google Scholar] [CrossRef]
- Ozturk, I.; Tornuk, F.; Caliskan-Aydogan, O.; Durak, M.Z.; Sagdic, O. Decontamination of Iceberg Lettuce by Some Plant Hydrosols. LWT 2016, 74, 48–54. [Google Scholar] [CrossRef]
- Li, Y.; Xu, X.; Wu, Y.; Zhuang, J.; Zhang, X.; Zhang, H.; Lei, B.; Hu, C.; Liu, Y. A Review on the Effects of Carbon Dots in Plant Systems. Mater. Chem. Front. 2020, 4, 437–448. [Google Scholar] [CrossRef]
- Servin, A.; Elmer, W.; Mukherjee, A.; de la Torre-Roche, R.; Hamdi, H.; White, J.C.; Bindraban, P.; Dimkpa, C. A Review of the Use of Engineered Nanomaterials to Suppress Plant Disease and Enhance Crop Yield. J. Nanopart. Res. 2015, 17, 92. [Google Scholar] [CrossRef]
- Somala, N.; Laosinwattana, C.; Teerarak, M. Formulation Process, Physical Stability and Herbicidal Activities of Cymbopogon Nardus Essential Oil-Based Nanoemulsion. Sci. Rep. 2022, 12, 10280. [Google Scholar] [CrossRef] [PubMed]
- Abdollahdokht, D.; Gao, Y.; Faramarz, S.; Poustforoosh, A.; Abbasi, M.; Asadikaram, G.; Nematollahi, M.H. Conventional Agrochemicals towards Nano-Biopesticides: An Overview on Recent Advances. Chem. Biol. Technol. Agric. 2022, 9, 13. [Google Scholar] [CrossRef]
- Kumar, A.; Choudhary, A.; Kaur, H.; Mehta, S.; Husen, A. Smart Nanomaterial and Nanocomposite with Advanced Agrochemical Activities. Nanoscale Res. Lett. 2021, 16, 156. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Mallya, R.; Suvarna, V.; Khan, T.A.; Momin, M.; Omri, A. Nanoparticles—Attractive Carriers of Antimicrobial Essential Oils. Antibiotics 2022, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Nerio, L.S.; Olivero-Verbel, J.; Stashenko, E. Repellent Activity of Essential Oils: A Review. Bioresour. Technol. 2010, 101, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Khursheed, A.; Rather, M.A.; Jain, V.; Wani, A.R.; Rasool, S.; Nazir, R.; Malik, N.A.; Majid, S.A. Plant Based Natural Products as Potential Ecofriendly and Safer Biopesticides: A Comprehensive Overview of Their Advantages over Conventional Pesticides, Limitations and Regulatory Aspects. Microb. Pathog. 2022, 173, 105854. [Google Scholar] [CrossRef]
- Traka, C.K.; Petrakis, E.A.; Kimbaris, A.C.; Polissiou, M.G.; Perdikis, D.C. Effects of Ocimum Basilicumand Ruta Chalepensis Hydrosols on Aphis Gossypii and Tetranychus Urticae. J. Appl. Entomol. 2018, 142, 413–420. [Google Scholar] [CrossRef]
- Petrakis, E.A.; Kimbaris, A.C.; Lykouressis, D.P.; Polissiou, M.G.; Perdikis, D.C. Hydrosols Evaluation in Pest Control: Insecticidal and Settling Inhibition Potential against Myzus Persicae (Sulzer). J. Appl. Entomol. 2015, 139, 260–267. [Google Scholar] [CrossRef]
- Zekri, N.; Handaq, N.; el Caidi, A.; Zair, T.; Alaoui El Belghiti, M. Insecticidal Effect of Mentha Pulegium L. and Mentha Suaveolens Ehrh. Hydrosols against a Pest of Citrus, Toxoptera Aurantii (Aphididae). Res. Chem. Intermed. 2016, 42, 1639–1649. [Google Scholar] [CrossRef]
- Božović, M.; Pirolli, A.; Ragno, R. Mentha Suaveolens Ehrh. (Lamiaceae) Essential Oil and Its Main Constituent Piperitenone Oxide: Biological Activities and Chemistry. Molecules 2015, 20, 8605–8633. [Google Scholar] [CrossRef] [Green Version]
- Ruberto, G.; Baratta, M.T.; Deans, S.G.; Dorman, H.J.D. Antioxidant and Antimicrobial Activity of Foeniculum Vulgare and Crithmum Maritimum Essential Oils. Planta Med. 2000, 66, 687–693. [Google Scholar] [CrossRef] [PubMed]
- lo Cantore, P.; Iacobellis, N.S.; de Marco, A.; Capasso, F.; Senatore, F. Antibacterial Activity of Coriandrum Sativum L. and Foeniculum Vulgare Miller Var. Vulgare (Miller) Essential Oils. J. Agric. Food Chem. 2004, 52, 7862–7866. [Google Scholar] [CrossRef] [PubMed]
- Kalleli, F.; Abid, G.; ben Salem, I.; Boughalleb-M’hamdi, N.; M’hamdi, M. Essential Oil from Fennel Seeds (Foeniculum Vulgare) Reduces Fusarium Wilt of Tomato (Solanum Lycopersicon). Phytopathol. Mediterr. 2020, 59, 63–76. [Google Scholar]
- Pavela, R. Essential Oils from Foeniculum Vulgare Miller as a Safe Environmental Insecticide against the Aphid Myzus Persicae Sulzer. Environ. Sci. Pollut. Res. 2018, 25, 10904–10910. [Google Scholar] [CrossRef] [PubMed]
- Gal-On, A. Zucchini Yellow Mosaic Virus: Insect Transmission and Pathogenicity? The Tails of Two Proteins. Mol. Plant Pathol. 2007, 8, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Walkey, D.G.A.; Lecoq, H.; Collier, R.; Dobson, S. Studies on the Control of Zucchini Yellow Mosaic Virus in Courgettes by Mild Strain Protection. Plant Pathol. 1992, 41, 762–771. [Google Scholar] [CrossRef]
- Desbiez, C.; Lecoq, H. Zucchini Yellow Mosaic Virus. Plant Pathol. 1997, 46, 809–829. [Google Scholar] [CrossRef]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification and Information Guide, 2nd ed.; John Wiley & Sons: Chichester, UK, 2000. [Google Scholar]
- Shi, X.; Jiang, L.; Wang, H.; Qiao, K.; Wang, D.; Wang, K. Toxicities and Sublethal Effects of Seven Neonicotinoid Insecticides on Survival, Growth and Reproduction of Imidacloprid-Resistant Cotton Aphid, Aphis Gossypii. Pest. Manag. Sci. 2011, 67, 1528–1533. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Hwang, B.K. An Important Role of the Pepper Phenylalanine Ammonia-Lyase Gene (PAL1) in Salicylic Acid-Dependent Signalling of the Defence Response to Microbial Pathogens. J. Exp. Bot. 2014, 65, 2295–2306. [Google Scholar] [CrossRef] [Green Version]
- Abdelkhalek, A.; Dessoki, E.S.; Hafez, E. Polyphenolic Genes Expression Pattern and Their Role in Viral Resistance in Tomato Plant Infected with Tobacco Mosaic Virus. Biosci. Res. 2018, 15, 3349–3356. [Google Scholar]
- Zhao, L.; Feng, C.; Wu, K.; Chen, W.; Chen, Y.; Hao, X.; Wu, Y. Advances and Prospects in Biogenic Substances against Plant Virus: A Review. Pestic. Biochem. Physiol. 2017, 135, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential Oils as Potential Alternative Biocontrol Products against Plant Pathogens and Weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isman, M.B. Bioinsecticides Based on Plant Essential Oils: A Short Overview. Z. Für. Nat. C 2020, 75, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Barra, A. Factors Affecting Chemical Variability of Essential Oils: A Review of Recent Developments. Nat. Prod. Commun. 2009, 4, 1147–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutour, S.; Bradesi, P.; Casanova, J.; Tomi, F. Composition and Chemical Variability of Mentha Suaveolens Ssp. Suaveolens and M. Suaveolens Ssp. Insularis from Corsica. Chem. Biodivers. 2010, 7, 1002–1008. [Google Scholar] [CrossRef]
- Lorenzo, D.; Paz, D.; Dellacassa, E.; Davies, P.; Vila, R.; Cañigueral, S. Essential Oils of Mentha Pulegium and Mentha Rotundifolia from Uruguay. Braz. Arch. Biol. Technol. 2002, 45, 519–524. [Google Scholar] [CrossRef]
- Anwar, F.; Ali, M.; Hussain, A.I.; Shahid, M. Antioxidant and Antimicrobial Activities of Essential Oil and Extracts of Fennel (Foeniculum Vulgare Mill.) Seeds from Pakistan. Flavour. Fragr. J. 2009, 24, 170–176. [Google Scholar] [CrossRef]
- Civitelli, L.; Panella, S.; Marcocci, M.E.; de Petris, A.; Garzoli, S.; Pepi, F.; Vavala, E.; Ragno, R.; Nencioni, L.; Palamara, A.T.; et al. In Vitro Inhibition of Herpes Simplex Virus Type 1 Replication by Mentha Suaveolens Essential Oil and Its Main Component Piperitenone Oxide. Phytomedicine 2014, 21, 857–865. [Google Scholar] [CrossRef]
- Gulfraz, M.; Mehmood, S.; Minhas, N.; Jabeen, N.; Kausar, R.; Jabeen, K.; Arshad, G. Composition and Antimicrobial Properties of Essential Oil of Foeniculum Vulgare. Afr. J. Biotechnol. 2008, 7, 4364–4368. [Google Scholar]
- Šilha, D.; Švarcová, K.; Bajer, T.; Královec, K.; Tesařová, E.; Moučková, K.; Pejchalová, M.; Bajerová, P. Chemical Composition of Natural Hydrolates and Their Antimicrobial Activity on Arcobacter-Like Cells in Comparison with Other Microorganisms. Molecules 2020, 25, 5654. [Google Scholar] [CrossRef] [PubMed]
- la Camera, S.; Gouzerh, G.; Dhondt, S.; Hoffmann, L.; Fritig, B.; Legrand, M.; Heitz, T. Metabolic Reprogramming in Plant Innate Immunity: The Contributions of Phenylpropanoid and Oxylipin Pathways. Immunol. Rev. 2004, 198, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Gutha, L.R.; Casassa, L.F.; Harbertson, J.F.; Naidu, R.A. Modulation of Flavonoid Biosynthetic Pathway Genes and Anthocyanins Due to Virus Infection in Grapevine (Vitis Vinifera L.) Leaves. BMC Plant Biol. 2010, 10, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Cai, N.; Chen, J.; Wan, C. Clove Essential Oil as an Alternative Approach to Control Postharvest Blue Mold Caused by Penicillium Italicum in Citrus Fruit. Biomolecules 2019, 9, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Politi, M.; Menghini, L.; Conti, B.; Bedini, S.; Farina, P.; Cioni, P.L.; Braca, A.; de Leo, M. Reconsidering Hydrosols as Main Products of Aromatic Plants Manufactory: The Lavandin (Lavandula × Intermedia) Case Study in Tuscany. Molecules 2020, 25, 2225. [Google Scholar] [CrossRef]
- Sayed, S.; Soliman, M.M.; Al-Otaibi, S.; Hassan, M.M.; Elarrnaouty, S.-A.; Abozeid, S.M.; El-Shehawi, A.M. Toxicity, Deterrent and Repellent Activities of Four Essential Oils on Aphis Punicae (Hemiptera: Aphididae). Plants 2022, 11, 463. [Google Scholar] [CrossRef] [PubMed]
- Andrade, L.H.d.; Oliveira, J.V.d.; Lima, I.M.d.M.; Santana, M.F.d.; Breda, M.O. Efeito Repelente de Azadiractina e Óleos Essenciais Sobre Aphis Gossypii Glover (Hemiptera: Aphididae) Em Algodoeiro. Rev. Ciência Agronômica 2013, 44, 628–634. [Google Scholar] [CrossRef] [Green Version]
- Benddine, H.; Zaid, R.; Babaali, D.; Daoudi-Hacini, S. Biological Activity of Essential Oils of Myrtus Communis (Myrtaceae, Family) and Foeniculum Vulgare (Apiaceae, Family) on Open Fields Conditions against Corn Aphids Rhopalosiphum Maidis (Fitch, 1856) in Western Algeria. J. Saudi Soc. Agric. Sci. 2022. [Google Scholar] [CrossRef]
- Digilio, M.C.; Mancini, E.; Voto, E.; de Feo, V. Insecticide Activity of Mediterranean Essential Oils. J. Plant Interact. 2008, 3, 17–23. [Google Scholar] [CrossRef]
- Garzoli, S.; Pirolli, A.; Vavala, E.; di Sotto, A.; Sartorelli, G.; Božović, M.; Angiolella, L.; Mazzanti, G.; Pepi, F.; Ragno, R. Multidisciplinary Approach to Determine the Optimal Time and Period for Extracting the Essential Oil from Mentha Suaveolens Ehrh. Molecules 2015, 20, 9640–9655. [Google Scholar] [CrossRef] [Green Version]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-Independent MS/MS Deconvolution for Comprehensive Metabolome Analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Misra, B. Steps for Building an Open Source EI-MS Mass Spectral Library for GC-MS -Based Metabolomics. Available online: https://www.protocols.io/view/steps-for-building-an-open-source-ei-ms-mass-spect-eq2ly33rqgx9/v1 (accessed on 20 December 2022).
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manglli, A.; Bertin, S.; Tomassoli, L. Preliminary Analysis of ZYMV and WMV Interaction in Mixed Infection by ΔΔCt Rt-QPCR. In Proceedings of the International Advances in Plant Virology, Rome, Italy, 29 October 2019. [Google Scholar]
- Zhang, S.; Liu, J.; Xu, B.; Zhou, J. Differential Responses of Cucurbita Pepo to Podosphaera Xanthii Reveal the Mechanism of Powdery Mildew Disease Resistance in Pumpkin. Front. Plant Sci. 2021, 12, 633221. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Obrero, Á.; Die, J.v.; Román, B.; Gómez, P.; Nadal, S.; González-Verdejo, C.I. Selection of Reference Genes for Gene Expression Studies in Zucchini (Cucurbita Pepo) Using QPCR. J. Agric. Food Chem. 2011, 59, 5402–5411. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Class of Components | MS EO | MS HS | FV EO | FV HS |
|---|---|---|---|---|
| Monoterpene hydrocarbons | 21.13 | - | 26.91 | 6.02 |
| Oxygenated monoterpenes | 17.93 | 16.09 | 33.19 | 76.36 |
| Sesquiterpene hydrocarbons | 2.51 | - | 0.12 | - |
| Phenolic compounds | 4.11 | 3.94 | 0.10 | - |
| Alcohols | 7.55 | 1.52 | - | - |
| Esters | 9.85 | 4.74 | 18.47 | - |
| Aromatic compounds | 1.21 | - | 11.93 | - |
| Nitrogen compounds | - | 68.46 | 1.14 | - |
| Oxygenated heterobicyclic | 0.87 | - | - | - |
| Total identified | 83.00 | 94.75 | 92.45 | 82.38 |
| Time | MS EO | MS HS | FV EO | FV HS | ||||
|---|---|---|---|---|---|---|---|---|
| Mortality% | Chi-Square | Mortality% | Chi-Square | Mortality% | Chi-Square | Mortality% | Chi-Square | |
| 24 h | 4.0 | 1.72 | 58.6 **** | 80.11 | 8.0 | 0.01 | 25.0 *** | 13.36 |
| 48 h | 5.2 ** | 10.76 | 62.1 **** | 34.57 | 15.2 | 1.08 | 26.7 | 1.10 |
| 72 h | 11.0 ** | 7.93 | 54.5 ** | 7.64 | 7.7 *** | 10.94 | 50.6 ** | 9.74 |
| 96 h | 16.0 | 0.05 | 40.0 | 3.61 | 27.8 | 2.02 | 48.1 *** | 12.66 |
| Time | MS EO | MS HS | FV EO | FV HS | ||||
|---|---|---|---|---|---|---|---|---|
| Progeny | Chi-Square | Progeny | Chi-Square | Progeny | Chi-Square | Progeny | Chi-Square | |
| 24 h | 0.82 | 0.93 | 0.13 **** | 66.10 | 0.55 ** | 7.45 | 1.03 | 0.02 |
| 48 h | 1.09 | 0.07 | 0.32 **** | 23.56 | 0.88 | 1.62 | 1.48 | 1.31 |
| 72 h | 1.30 | 0.18 | 0.48 ** | 10.19 | 1.24 | 0.35 | 1.27 | 0.20 |
| 96 h | 1.19 | 0.08 | 0.47 | 3.19 | 1.50 | 1.58 | 1.38 | 0.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taglienti, A.; Donati, L.; Dragone, I.; Ferretti, L.; Gentili, A.; Araniti, F.; Sapienza, F.; Astolfi, R.; Fiorentino, S.; Vecchiarelli, V.; et al. In Vivo Antiphytoviral and Aphid Repellency Activity of Essential Oils and Hydrosols from Mentha suaveolens and Foeniculum vulgare to Control Zucchini Yellow Mosaic Virus and Its Vector Aphis gossypii. Plants 2023, 12, 1078. https://doi.org/10.3390/plants12051078
Taglienti A, Donati L, Dragone I, Ferretti L, Gentili A, Araniti F, Sapienza F, Astolfi R, Fiorentino S, Vecchiarelli V, et al. In Vivo Antiphytoviral and Aphid Repellency Activity of Essential Oils and Hydrosols from Mentha suaveolens and Foeniculum vulgare to Control Zucchini Yellow Mosaic Virus and Its Vector Aphis gossypii. Plants. 2023; 12(5):1078. https://doi.org/10.3390/plants12051078
Chicago/Turabian StyleTaglienti, Anna, Livia Donati, Immacolata Dragone, Luca Ferretti, Andrea Gentili, Fabrizio Araniti, Filippo Sapienza, Roberta Astolfi, Simona Fiorentino, Valerio Vecchiarelli, and et al. 2023. "In Vivo Antiphytoviral and Aphid Repellency Activity of Essential Oils and Hydrosols from Mentha suaveolens and Foeniculum vulgare to Control Zucchini Yellow Mosaic Virus and Its Vector Aphis gossypii" Plants 12, no. 5: 1078. https://doi.org/10.3390/plants12051078
APA StyleTaglienti, A., Donati, L., Dragone, I., Ferretti, L., Gentili, A., Araniti, F., Sapienza, F., Astolfi, R., Fiorentino, S., Vecchiarelli, V., Papalini, C., Ragno, R., & Bertin, S. (2023). In Vivo Antiphytoviral and Aphid Repellency Activity of Essential Oils and Hydrosols from Mentha suaveolens and Foeniculum vulgare to Control Zucchini Yellow Mosaic Virus and Its Vector Aphis gossypii. Plants, 12(5), 1078. https://doi.org/10.3390/plants12051078