1. Introduction
Ditylenchus Filipjev [
1] is the largest genus of the family Anguinidae Nicoll, [
2,
3] that has adapted to a wide range of ecological processes, including phytoparasitism [
4,
5,
6], mycophagy [
7,
8,
9], phoretic association with insects in soil [
10,
11], biocontrol of weeds [
12], and acting as a vector for
Corynebacterium spp. [
13,
14,
15]. The majority of studied
Ditylenchus members are fungal feeders [
3,
16]. However,
D. africanus Wendt, Swart, Vrain and Webster [
17];
D. angustus (Butler) Filipjev [
1,
18];
D. destructor Thorne [
19];
D. dipsaci (Kühn) Filipjev [
1,
20];
D. gallaeformans Oliveira, Santin, Seni, Dietrich, Salazar, Subbotin, Mundo-Ocampo, Goldenberg and Barreto [
12];
D. gigas Vovlas, Troccoli, Palomares-Rius, De Luca, Liebanas, Landa, Subbotin and Castillo [
6]; and
D. myceliophagus Goodey [
7] have attracted attention due to their parasitic potential and quarantine regulations [
4,
6,
21,
22,
23,
24]. Several
Ditylenchus species are polyphagous and display endoparasitic behaviour; as a result, these species can be disseminated through seeds, plant material or contaminated field equipment [
8,
25,
26].
The regulated status of
Ditylenchus spurred widespread research, which led to the discovery of numerous new species without giving adequate consideration to the limits of species variability [
27]. Several taxonomists have reviewed the genus characteristics in attempts to limit the number of valid species by revisions and synonymization [
3,
27,
28]. More recently, Hashemi and Karegar [
16] updated the entire genus
Ditylenchus and documented 60 nominal species; presently, the genus contains 63 species. Of all valid species, the literature lists
D. filimus Anderson [
29] and
D. dryadis Anderson and Mulvey [
30] as Canadian native species, whereas
D. valveus Thorne and Malek [
31] and
D. weischeri Chizhov, Borisov and Subbotin [
32] were listed as new records from Canada. In addition,
D. destructor Thorne [
19] and
D. dipsaci (Kühn, 1857) Filipjev [
1,
20] were reported from isolated locations (British Columbia, Ontario, and Prince Edward Island) and subjected to strict quarantine regulations to prevent spread [
13,
21,
26,
29,
30,
33,
34,
35,
36]. Among the isolated species described in Canada,
D. dipsaci is the only one reported from Alberta where it was found in alfalfa-growing areas [
13]. Therefore, the cultivated areas of southern Alberta are regularly surveyed and examined for the presence of nematode pest species [
37,
38,
39]. These surveys include detecting not only the regulated pest species but also new and unusual nematode species that may pose a challenge to agricultural produce. During our nematode inventory survey, we detected four different populations of
Ditylenchus in no-till fields (in which soil was undisturbed from harvest to planting). Due to its regulatory importance, the recovered unidentified populations were immediately processed for morphological and molecular characterization. Light microscopy indicated the presence of delicate stylets (>10 µm long), six lateral lines and round to pointy tail tips. The populations were identified as members of the
D. triformis group, which includes species with six lateral lines, a rounded tail tip, and a mycophagous life cycle.
Further morphological comparisons revealed their identity as
D. anchilisposomus (Tarjan) Fortuner [
28,
40],
D. clarus Thorne and Malek [
31],
D. tenuidens Gritsenko [
41], and
D. valveus. All of these species are new records in Canada except for the
D. valveus.
The objectives of the present study were to: (1) document the presence of Ditylenchus species from cultivated areas in southern Alberta, (2) provide detailed morphological and molecular characterizations of these species, and (3) study the phylogenetic relationship of these species with related ones. The results of this study will aid in the decision of whether these species should be incorporated into nematode management programs since nontarget species can become pests due to changes in cropping patterns or climate.
2. Results
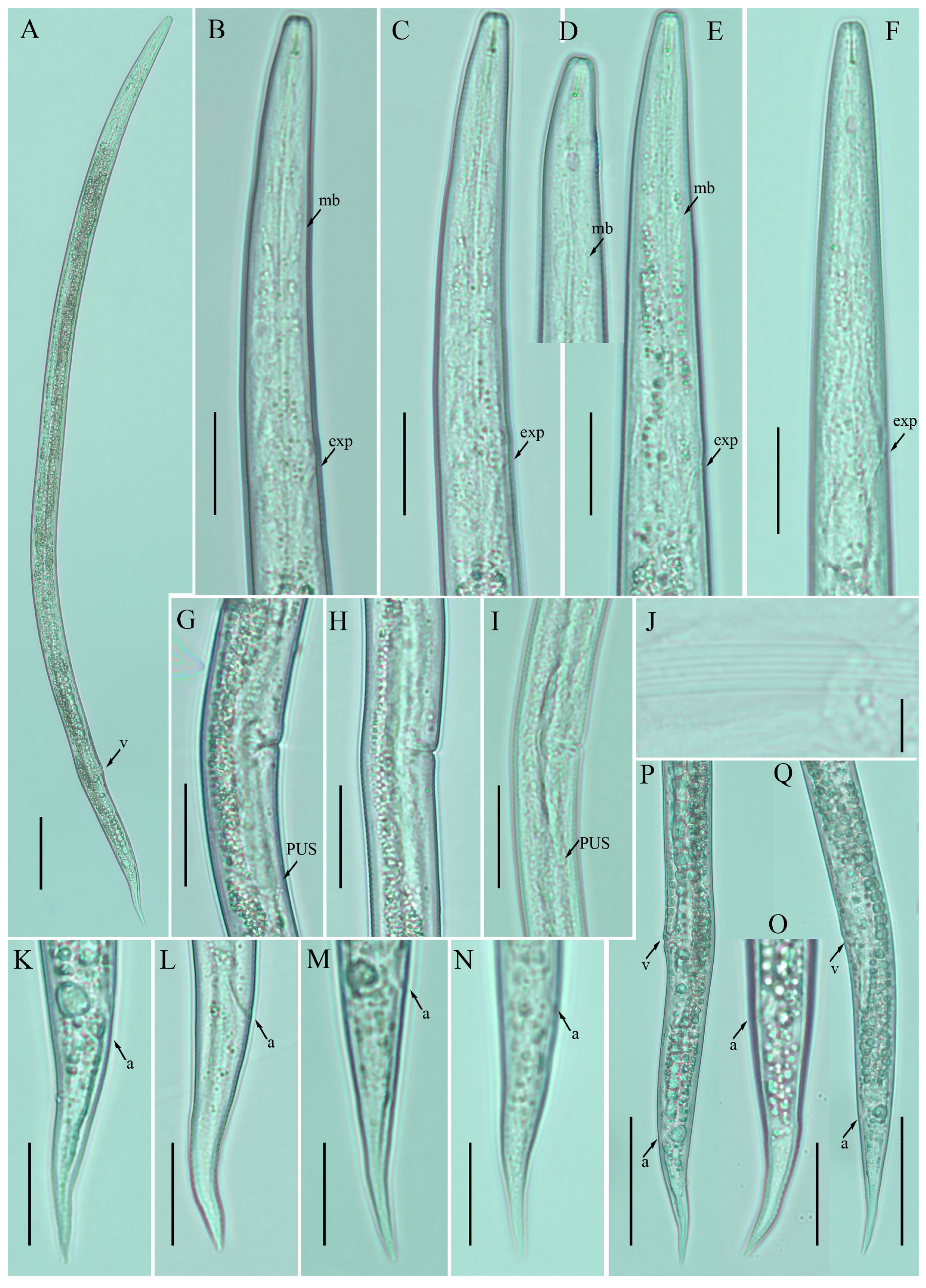
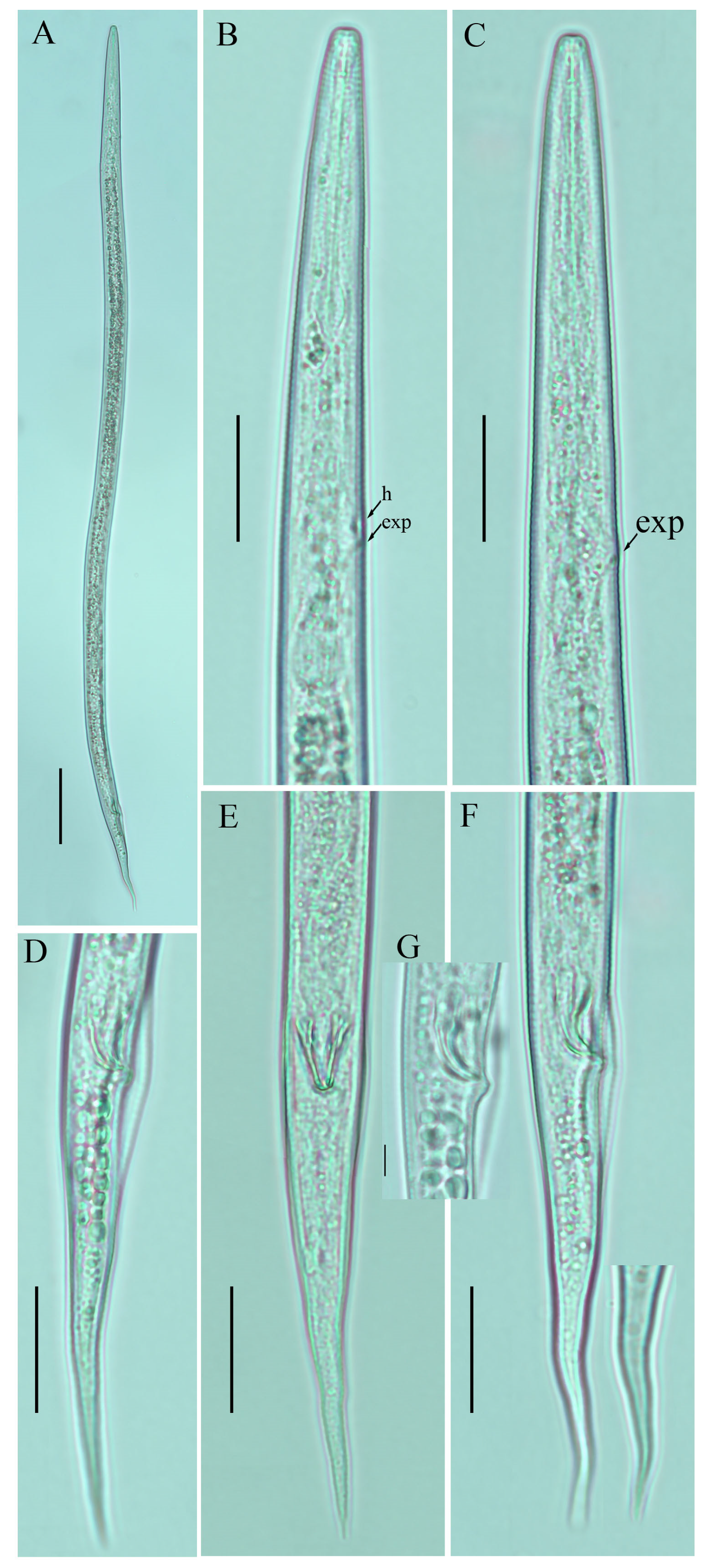
2.1. Description of Ditylenchus anchilisposomus (Tarjan) Fortuner [28,40]
Female: Body cylindrical, slightly ventral arcuate when heat-relaxed. Cuticle finely annulated; lateral field with six incisures. Lip region low, anteriorly truncated, with rounded margins, continuous with body contour. Stylet delicate; conus 30–35% of total stylet length. Stylet knobs small and rounded. Dorsal pharyngeal gland orifice (DGO) situated close to stylet knobs. Median pharyngeal bulb moderately developed, with indistinct elongated valve plates. Isthmus slender, encircled with a nerve ring. Hemizonid streak-like, two to three annuli anterior to the secretory–excretory pore. This pore located slightly in the range of the anterior level of the pharyngeal basal bulb. Basal pharyngeal bulb pyriform to elongated, slightly overlapping intestine. Ovary outstretched, oocytes arranged in a single row, spermatheca tubular devoid of sperm, in line with the genital tract. Vagina straight; vulva a transverse slit occupying less than half of the corresponding body width. Vulval lips simple, not protruding. Post-vulval uterine sac empty; tube-like sac along the ventral body wall ca. more than one vulval body width long. Anus a transverse slit. Tail conical; the posterior half of the tail curved ventrally, giving rise to a rounded to pointed terminus (
Figure 1;
Table 1).
Male: not detected.
Remarks:
Ditylenchus anchilisposomus was originally described from California, USA in the rhizosphere of grass as
Pseudhalenchus anchilisposomus [
40]. This genus was formed to accommodate the species that were closely related to the genera,
Ditylenchus,
Halenchus Cobb [
42] and
Tylenchus Bastian [
43]. Taxonomically,
Pseudhalenchus Tarjan [
40] can be differentiated from
Ditylenchus by having overlapped esophageal glands. Thorne [
44] studied
D. destructor and reported that the position of the pharyngeal lobe is a host-dependent character. Observations of
D. myceliophagus also suggest that the length of the pharyngeal overlap varies significantly during the life cycle of an individual [
28]. Consequently,
P. anchilisposomus was transferred to the genus,
Ditylenchus, as
D. anchilisposomus by Fortuner [
28]. After the formal recognition, the species was reported from Nebraska, USA and Iran without indication of the host association [
16,
31]. In the present study, we found
D. anchilisposomus in a no-till field (previously planted with potatoes) from southern Alberta. The description of the Nebraska population provided the measurements of a single specimen, and, therefore, cannot be used for intraspecies variation comparison. Moreover, the description from Iran is devoid of photo documentation or illustrations. Alternately, our
D. anchilisposomus population provides a complete set of morphological and morphometric characteristics, which are consistent with the original description’s characteristics and can be used for further molecular and taxonomical considerations (
Table 1).
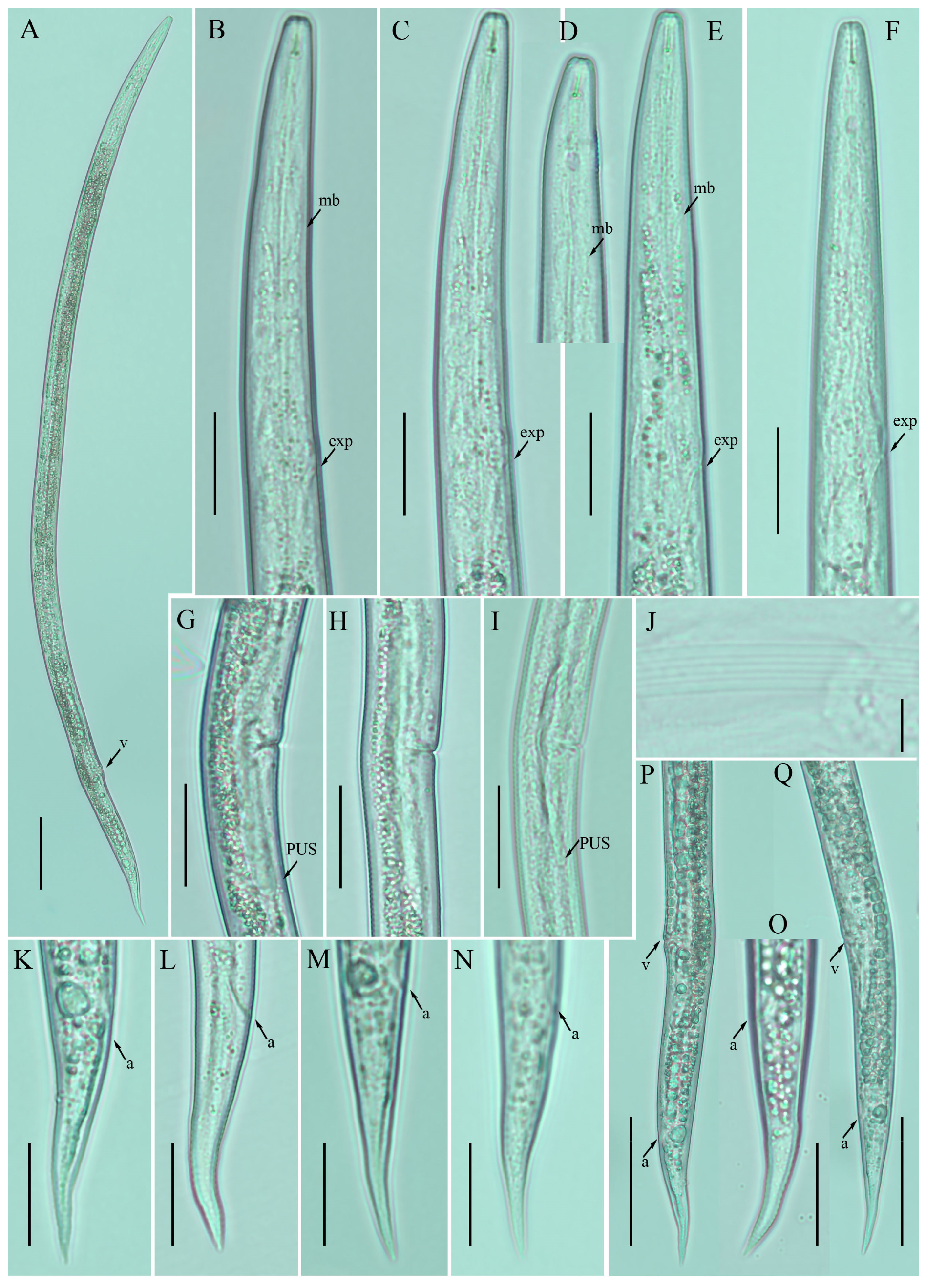
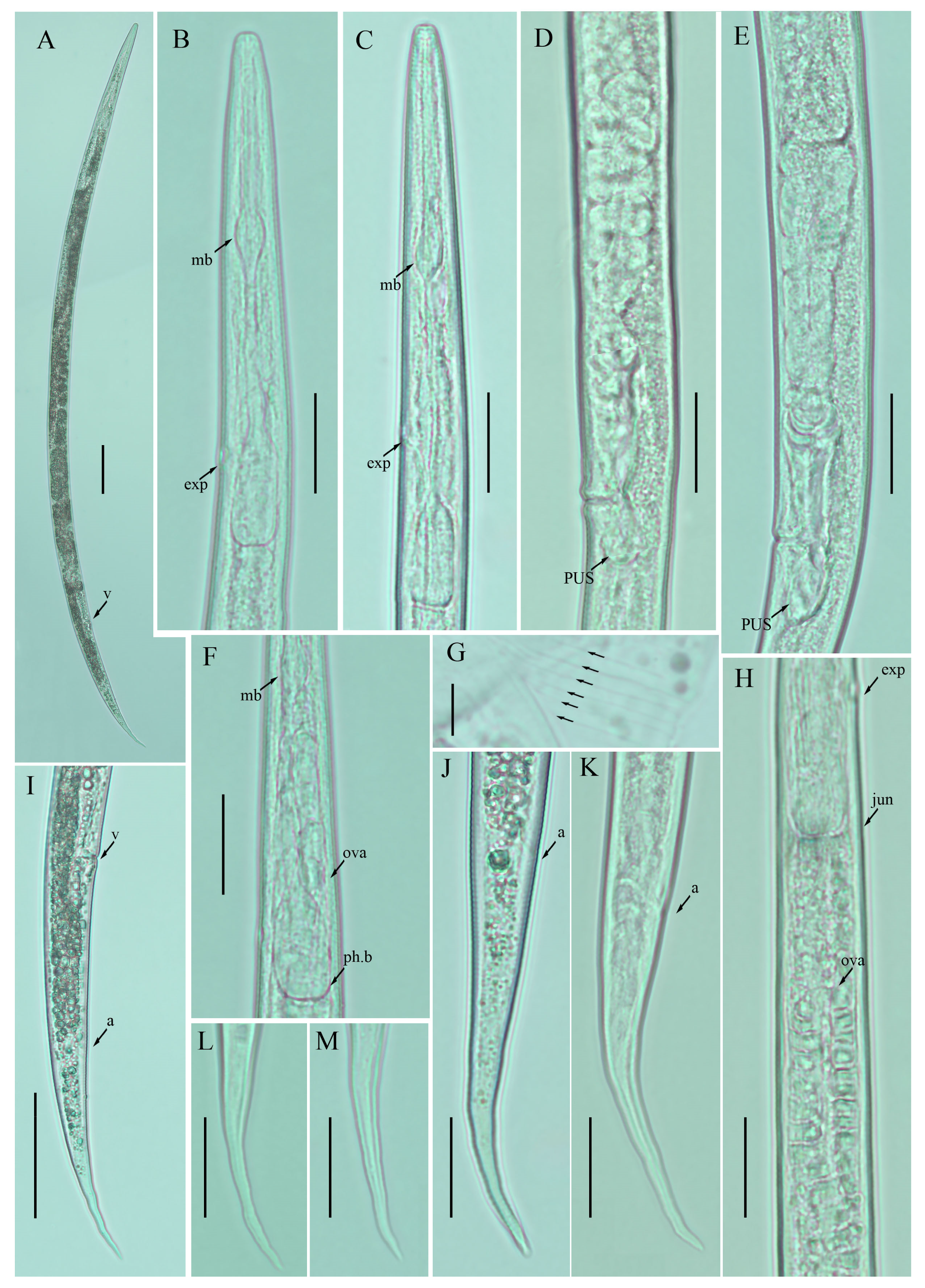
2.2. Description of Ditylenchus clarus Thorne and Malek [31]
Female: Body cylindrical, medium-sized, slightly ventral arcuate when heat-relaxed. Cuticle finely annulated; lateral field with six incisures. Lip region low, anteriorly flattened, the width is twice the height of the lip region. Stylet delicate; conus 30–40% of total stylet length. Stylet knobs small and rounded. Dorsal pharyngeal gland orifice (DGO) situated close to stylet knobs. Median pharyngeal bulb moderately developed with elongated valve plates. Isthmus slender encircled with a nerve ring. Hemizonid streak-like, one to three annuli anterior to the secretory–excretory pore. This pore located in the range of the pharyngeal basal bulb. Basal pharyngeal bulb pyriform to elongated, slightly overlapping intestine. Ovary outstretched in mature females, extending forward to the median bulb. Oocytes in a single row, columella well discernible, spermatheca tubular devoid of sperm, in line with the genital tract. A valvular apparatus present at the entrance to the uterine tract. Vagina straight; vulva a transverse slit occupying less than half of the corresponding body width. Vulval lips prominent slightly protruding in some specimens. Postvulval uterine sac small, empty, broad sac along the ventral body wall, extending ca. halfway to anus in some specimens. Anus a transverse slit. Tail conoid, stout; the posterior half of the tail curved ventrally, giving rise to a rounded terminus (
Figure 2).
Male: not detected.
Remarks: The species was originally described from a thicket in South Dakota, USA, without definitive host association [
31]. In the present study,
D. clarus was isolated from
Amaranthus sp. rhizosphere growing on a headland of a post harvested wheat field. The general appearance and dimensions of the southern Alberta population of
D. clarus were consistent with the original description except for the shorter stylet length (7.0–7.5 vs. 10 µm). After the formal description, the species was not reported again. Moreover, the original description likely provided the measurements of a single specimen and, therefore, cannot be used for intraspecies variation comparison. In contrast, southern Alberta population data can be used for further molecular and taxonomical considerations (
Table 2).
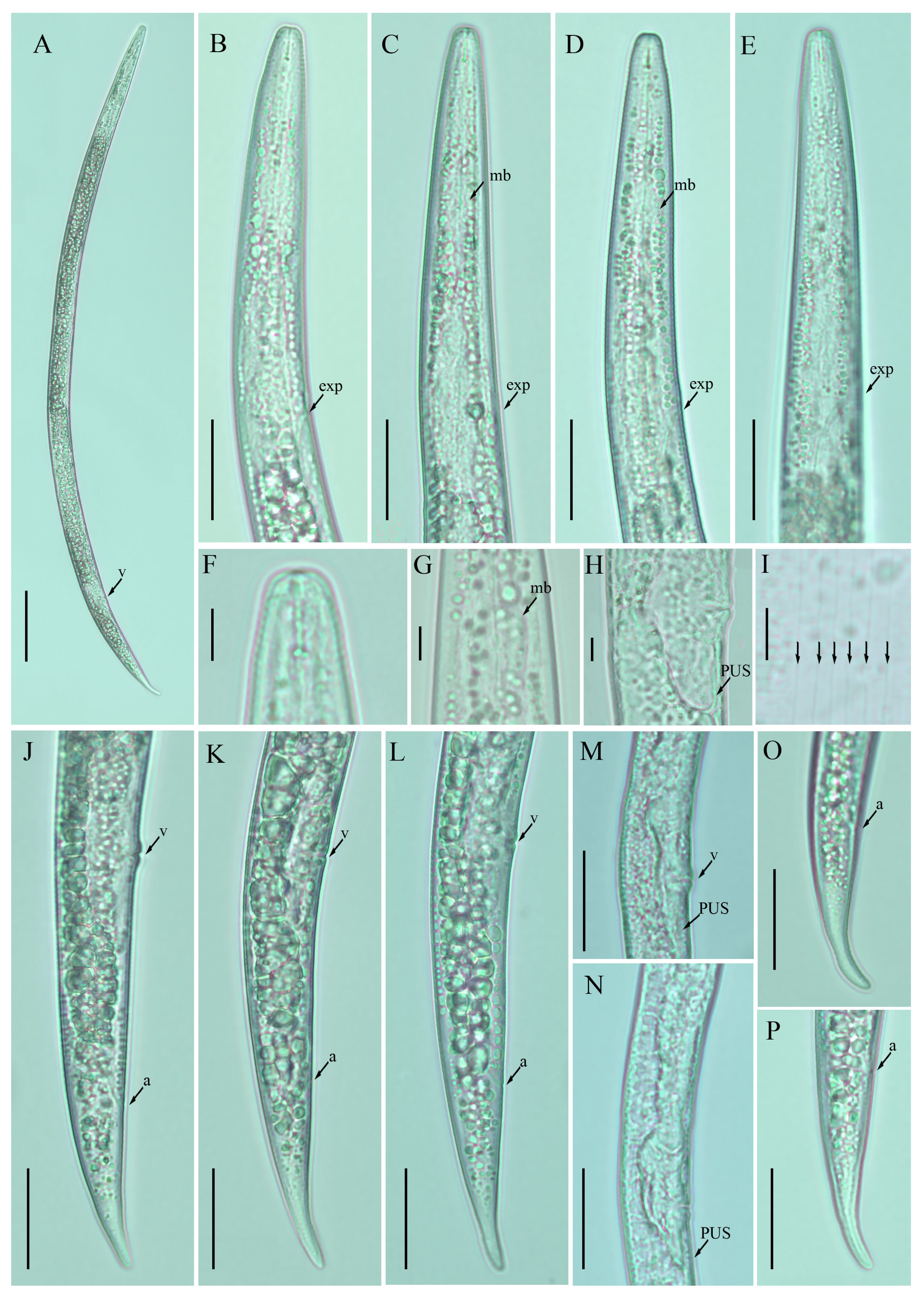
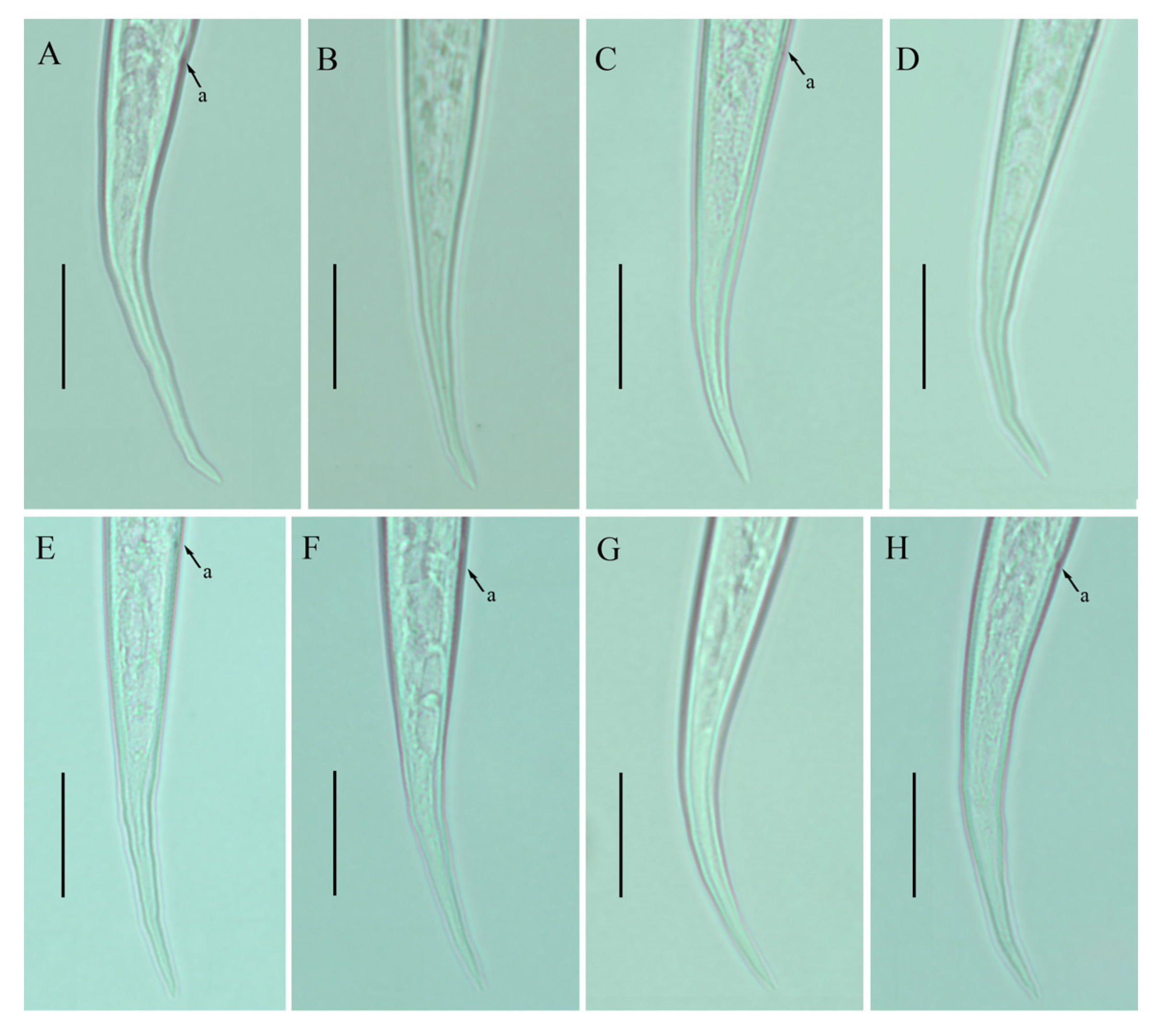
2.3. Description of Ditylenchus tenuidens Gritsenko [41]
Female: Body cylindrical, moderately long, slightly ventral arcuate when heat-relaxed. Cuticle finely annulated, lateral field with six incisures. Lip region narrow anteriorly flattened; width is twice the height of the lip region. Stylet delicate; conus 35–40% of total stylet length. Stylet knobs small, rounded, anteriorly sloping. Dorsal pharyngeal gland orifice (DGO) situated close to stylet knobs. Median pharyngeal bulb moderately developed with elongated valve plates. Isthmus slender encircled with a nerve ring. Hemizonid streak-like, one to three annuli anterior to the secretory–excretory pore. This pore located slightly anterior to or in the range of the anterior level of the pharyngeal basal bulb. Basal pharyngeal bulb pyriform to slightly elongated, abutting intestine. Ovary outstretched, oocytes in a single row, spermatheca and columella well discernible. Spermatheca rounded to elongate; in a few specimens, scarcely filled with rounded sperm generally in line with the genital tract. A valvular structure surrounds the entrance to the uterine tract. Vagina straight; vulva a transverse slit occupying less than half of the corresponding body width. Vulval lips simple, not protruding. Postvulval uterine sac empty tube-like, along the ventral body wall, one or more vulval body width long. Anus a transverse slit. Tail cylindrical elongated; the posterior half of the tail slender and tapers to form a pointed terminus.
Male: Body slightly shorter and more cylindroid than females. The anterior region is similar to females. Bursa leptoderan, starting at the same level as the anterior end of the spicules and extends less than half of the tail length. Spicules arcuate moderately long; gubernaculum simple embedded in cloacal sac. Cloacal opening smooth; tail shape is similar to that of females.
Remarks:
Ditylenchus tenuidens was originally described from Kyrgyzstan in the rhizospheres of potato and wheat [
41]. After the formal description, the species was reported from Poland and Sweden in leaf litter, forest organic soils, and moss samples by Brzeski [
27]. Two decades later, Mirshekari and Abdollahi [
45] found the same species in wheat and vegetable fields in Iran; however, the authors did not mention if the samples were collected before planting or after harvest. In the present study, we found
D. tenuidens in a no-till field (previously planted with grains). The morphological and morphometric characteristics of the southern Alberta
D. tenuidens population matched well with the original and subsequent descriptions except for a slightly longer body length and tail in the former population. Studies on the food source of
D. destructor and
D. myceliophagous found that the body length depended on the food supply, in particular the fungal species on which they fed [
46,
47]. Moreover, the body lengths correlated with the tail lengths [
28]. Considering this, the slightly longer bodies and tail lengths of the southern Alberta population can be considered intraspecies variation. Bionomically,
D. tenuidens have been reported from different habitats, which suggests that the host preference of
D. tenuidens is quite diverse (agronomic crops, leaf litter and moss). The description provided by Brzeski [
27] is taxonomically adequate, however, lacks photo documentation; the Iranian description [
45] is based on illustrations and is in the author’s native language (Farsi). Here, we described the southern Alberta
D. tenuidens population using the integrative taxonomical approach, making these data suitable for further molecular and taxonomical considerations (
Figure 3 and
Figure 4;
Table 3 and
Table 4).
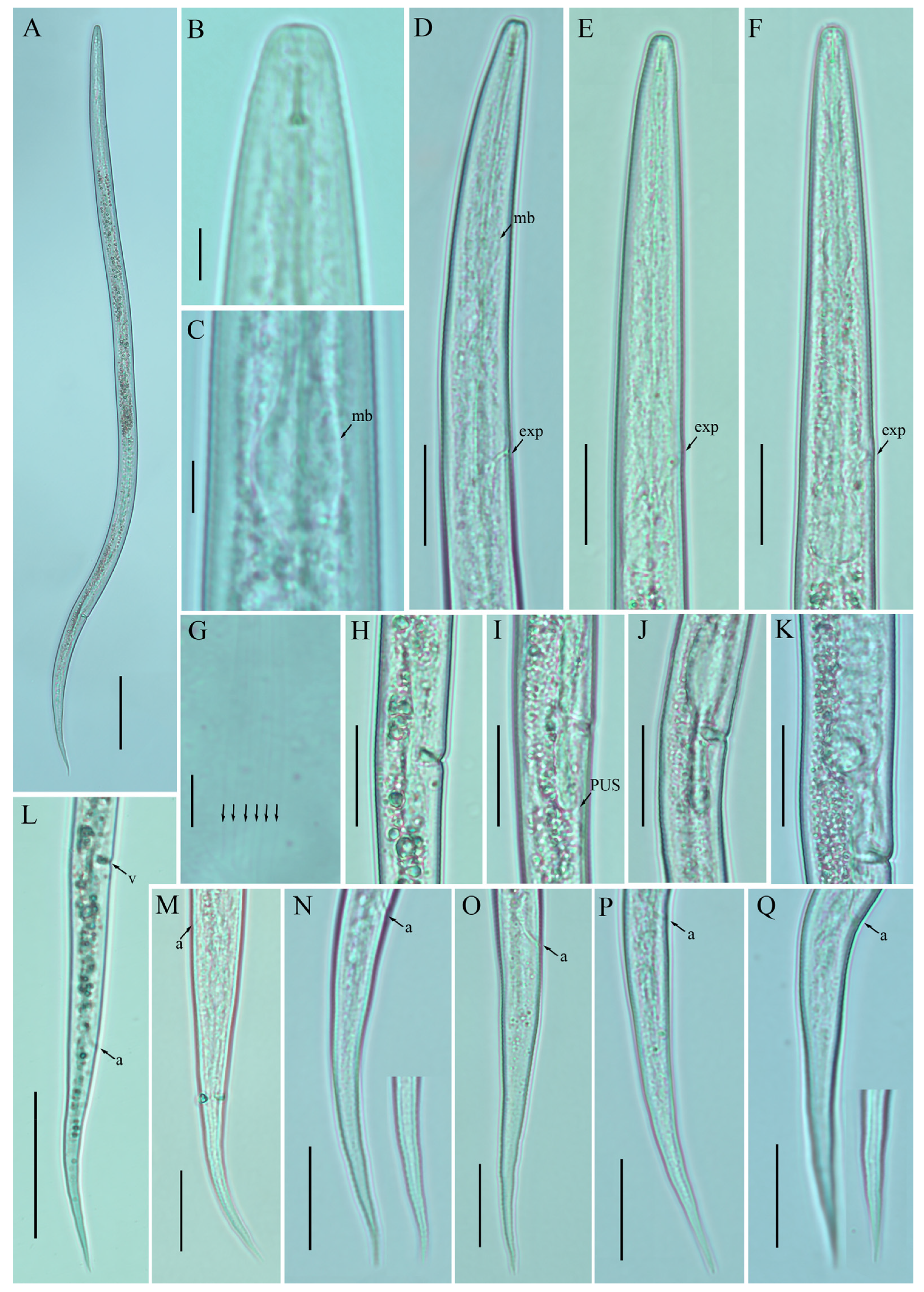
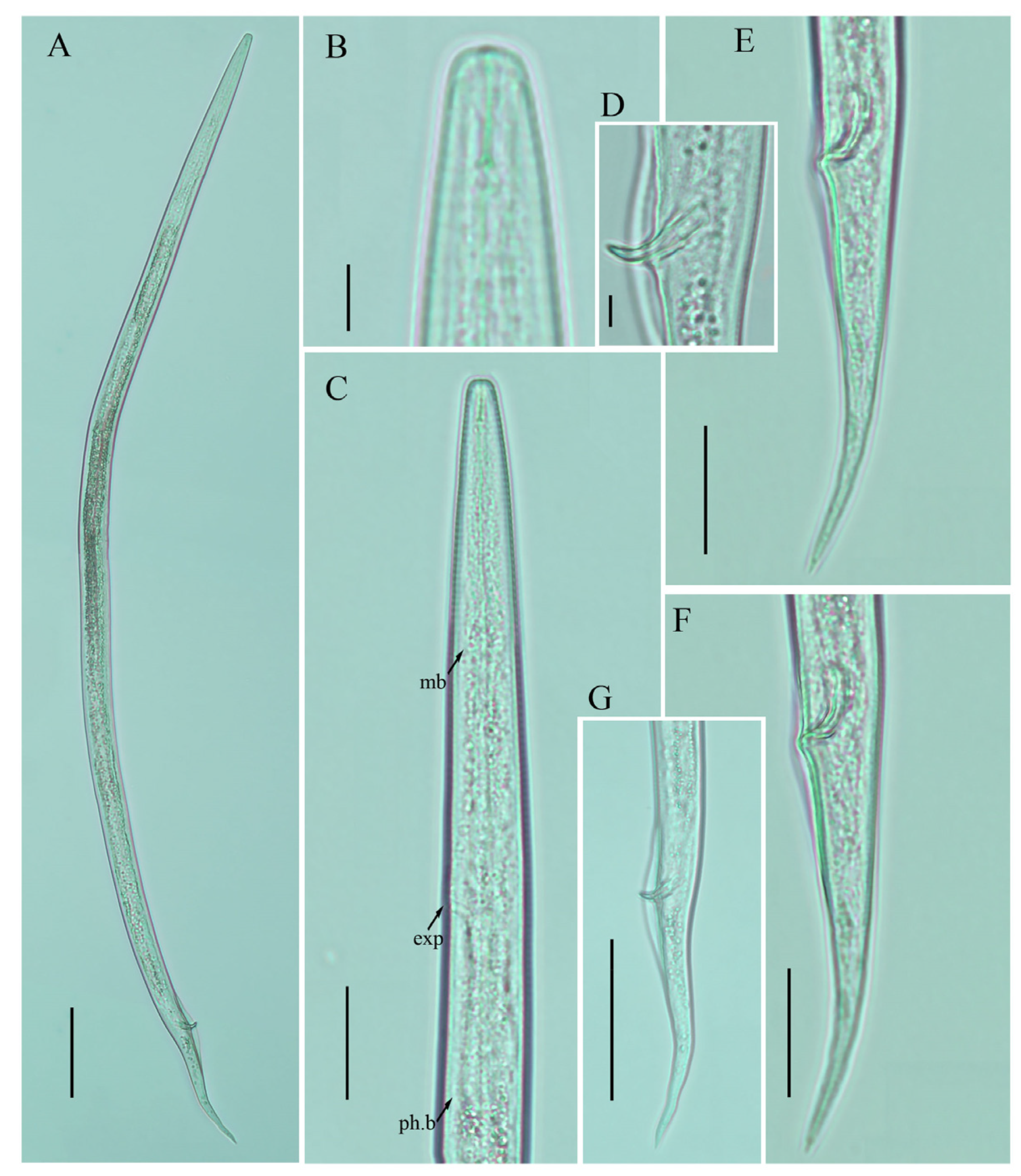
2.4. Description of Ditylenchus valveus Thorne and Malek [31]
Female: Body cylindrical, moderately long, slightly ventral arcuate when heat-relaxed. Cuticle finely annulated, lateral field with six incisures. Lip region low anteriorly flattened; width is twice the height of the lip region. Stylet delicate; conus 40–45% of total stylet length. Stylet knobs small, rounded, anteriorly sloping. Dorsal pharyngeal gland orifice (DGO) situated close to stylet knobs. Median pharyngeal bulb moderately developed with elongated valve plates. Isthmus slender encircled with a nerve ring. Hemizonid streak-like, one to three annuli anterior to the secretory–excretory pore. This pore located slightly anterior to or in the range of the anterior level of the pharyngeal basal bulb. Basal pharyngeal bulb pyriform to slightly elongated, abutting intestine. Ovary outstretched, reflexed in old females, extending forward to the median bulb. Oocytes in a single row, spermatheca and columella well discernible. Spermatheca, scarcely filled with rounded sperm, generally in line with the genital tract
A conspicuous valvular apparatus surrounds the entrance to the uterine tract. Vagina straight; t vulva a transverse slit occupying less than half of the corresponding body width. Vulval lips prominent and slightly protruding in some specimens. Post-vulval uterine sac appears as an empty broad sac along the ventral body wall ca. one vulval body width long. Anus a transverse slit. Tail cylindrical and elongated; the posterior half of the tail curved ventrally, giving rise to variable tail tips, i.e., rounded, slightly pointed, and pointed.
Male: Body slightly shorter and more cylindroid than females. The anterior region is similar to females. Bursa leptoderan, starting at the same level as the anterior end of the spicules, and extends about half of the tail length. Spicules arcuate, moderately long; gubernaculum simple embedded in cloacal sac. Cloacal opening smooth; tail shape is similar to that of females.
Remarks:
Ditylenchus valveus was originally reported from a ploughed field in South Dakota, USA [
31]. After the formal description, the species was reported from the rhizospheres of unknown bushes (Poland, Bulgaria [
27]) and olive trees (Iran [
16,
48]). In Canada, the species was previously reported from Manitoba in mushroom compost [
29], whereas in this study we found
D. valveus in a no-till field (previously planted with barley). The original description mentioned the presence of a conspicuous valvular structure at the entrance to the uterus, longer and reflexed ovaries, reaching past the basal pharyngeal bulb. These reproductive system characteristics were not observed in any of the subsequent descriptions except for our southernAlbertan population (see the female description section
Figure 5,
Figure 6 and
Figure 7;
Table 5 and
Table 6). Morphometrically, the Bulgarian population of
D. valveus is the smallest population as compared with the original and all other descriptions [
27]. The population from Iran [
48] has an unusually long postvulval uterine sac (PUS). Consequently, the morphology and morphometric characteristics of the
D. valveus population from southern Alberta fit more precisely with the characteristics presented in the original description.
2.5. Molecular Characterisation and Phylogeny of the Recovered Ditylenchus Species
We molecularly characterized the Ditylenchus species recovered in our survey using the 18S, D2–D3 of 28S and ITS rDNA diagnostic genes. We deposited the obtained sequences into the NCBI database under the following accession numbers: D. anchilisposomus (partial 18S, OP854643–OP854644; D2–D3 of 28S, OP854653–OP854654; and ITS, OQ058851–OQ058852); D. clarus (partial 18S, OP854645–OP854646; D2–D3 of 28S, OP854655–OP854656; and ITS, OQ058853–OQ058854); D. tenuidens (D2–D3 of 28S, OP854657–OP854658; and ITS, OQ058855–OQ058856); and D. valveus (partial 18S, OP854647–OP854652; D2–D3 of 28S, OP854659–OP854665; and ITS, OQ058857–OQ058861).
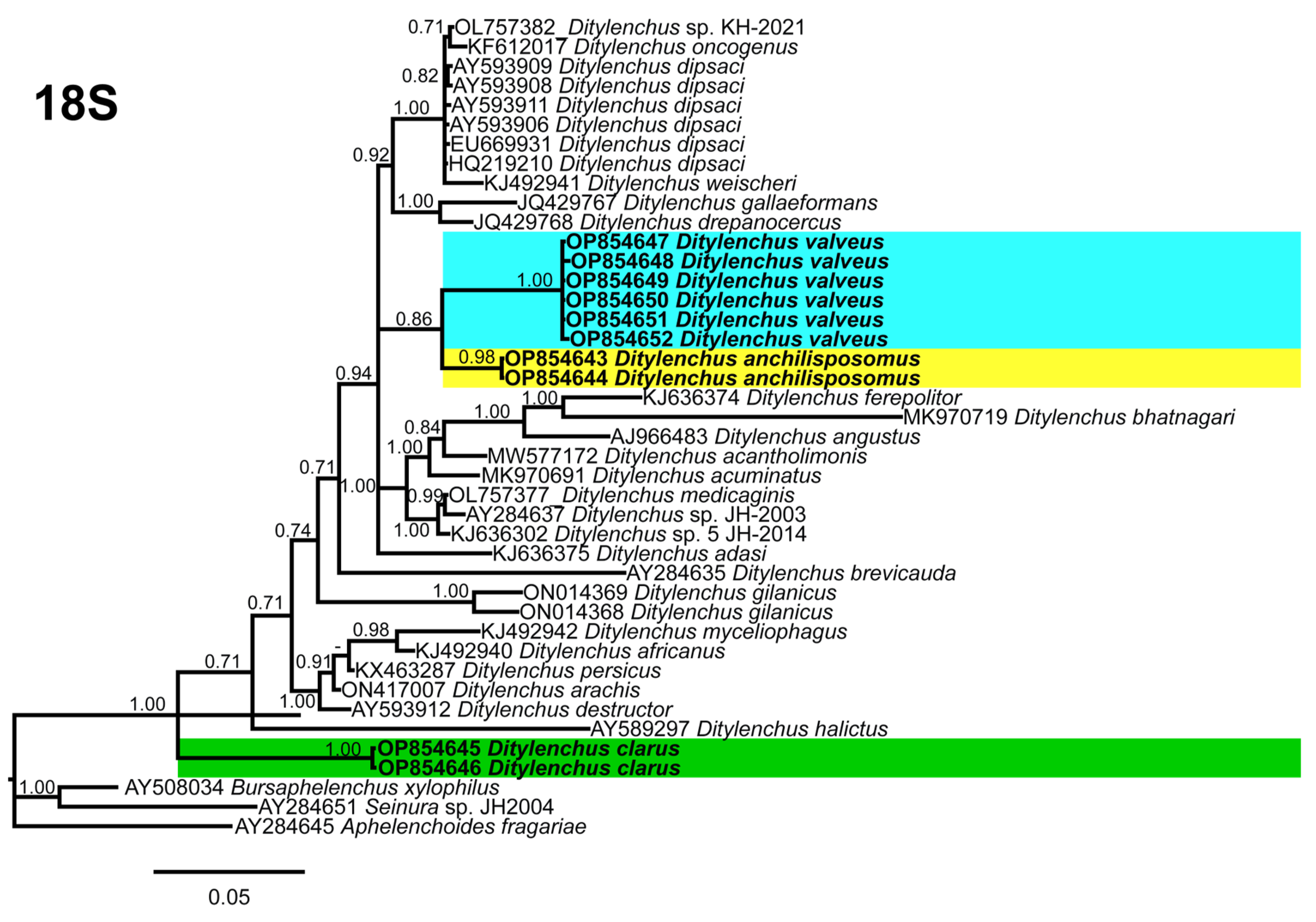
Phylogenetic trees based on the 18S, D2–D3 of 28S and ITS rDNA genes were constructed to study the phylogenetic relationships of the recovered species with the available
Ditylenchus species. The 18S tree (alignment 1694 bp;
Figure 8) was constructed with 39
Ditylenchus sequences and
Bursaphelenchus xylophilus Steiner and Buhrer (Nickle [
49,
50]) (AY508034),
Seinura sp. (AY284251) and
Aphelenchoides fragariae (Ritzema Bos) Christie, [
51,
52] (AY284545) as outgroup taxa. In this tree,
D. anchilisposomus and
D. valveus form a separate, moderately supporting clade (PP = 0.86) among other
Ditylenchus species, and
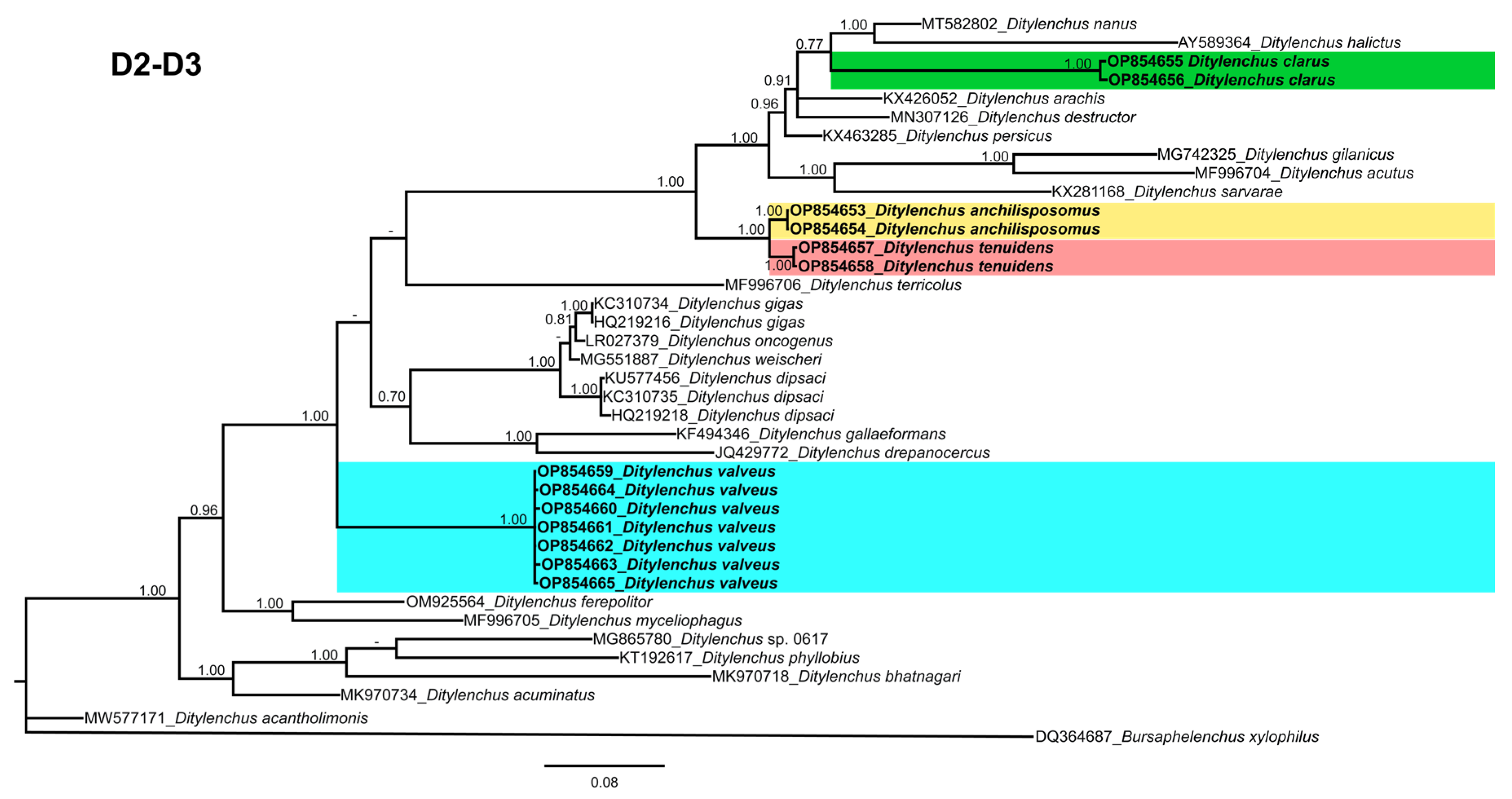
D. clarus occupies a basal position as a separate, well-supported clade (PP = 1.00). The 28S tree (alignment 769 bp;
Figure 9) was constructed with 38
Ditylenchus sequences with
B. xylophilus (DQ364687) as an outgroup taxon. In this tree,
D. clarus shares a branch with
D. halictus Giblin-Davis, Erteld, Kanzaki, Ye, Zeng and Center [
10], (AY589364) and
D. nanus Siddiqi [
53](MT582802) in a low support (PP = 77%), whereas
D. anchilisposomus,
D. tenuidens, and
D. valveus grouped independently among other
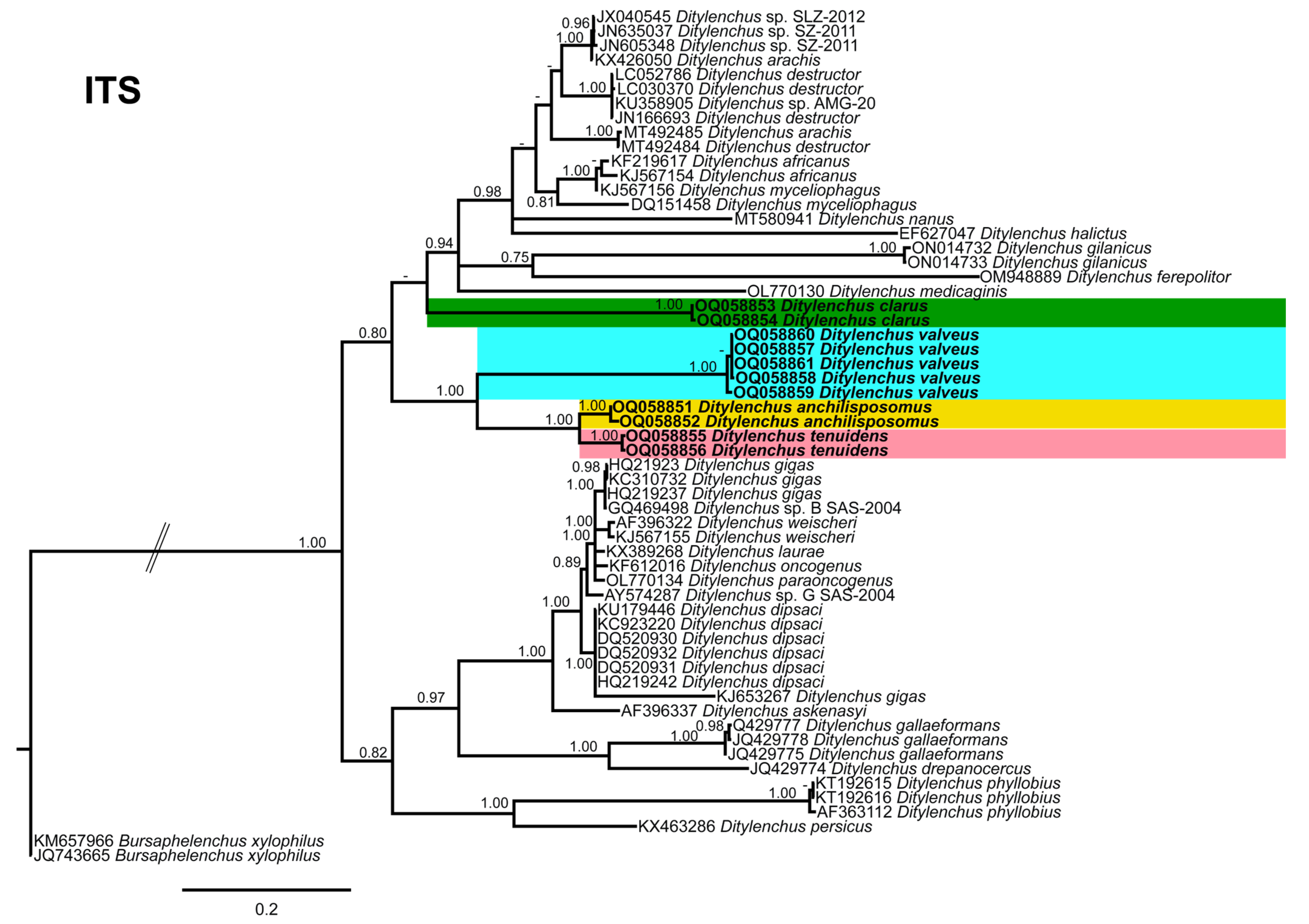
Ditylenchus species in well-supported clades (PP = 1.00). The ITS tree (alignment 786 bp:
Figure 10) was constructed from 57
Ditylenchus sequences and
B. xylophilus (KM657966, JQ743665) as an outgroup taxon. In the ITS tree,
D. anchilisposomus,
D. tenuidens and
D. clarus clustered together in a well-supported clade (PP = 1.00), whereas
D. valveus clustered in a separate low-supported clade (PP = 0.68), maintaining a middle position in the tree; however, all these species grouped independently among other
Ditylenchus species.
Since the recovered species are distinctly different from each other, and do not share striking phylogenetic affinity with other known species, we excluded the sequence similarity comparison for this dataset. The species recovered in this study all belong to the
D.
triformis group; however, this grouping is not evident in the phylogenetic analysis, which supports the notion that morphological grouping aids in prompt identification but does not reflect distinct lineages [
54]. Species in the genus
Ditylenchus have been reported from diverse hosts and different geographic locations; for most species, the biology and their association with the hosts are unknown. We speculate that the species bionomics has some role in the phylogenetic status, and the availability of genus-wide sequencing data will unequivocally elucidate the relationships of these species with each other.
3. Discussion
Based on morphological characteristics and bionomics, Siddiqi [
3] divided
Ditylenchus species into two main groups, namely, a
D.
dipsaci group (having four lateral lines, sharply pointed tails, and obligate parasitic behaviour) and a
D.
triformis group (having six lateral lines, rounded tail tips, and mainly fungal feeders). The
Ditylenchus species recovered in this study belong to the
D.
triformis group; however, at present, we are unable to ascertain any details relating to these species’ host association, dispersal, and survival. We found these species in no-till fields; therefore, it is unknown if these nematodes were feeding on soil fungal propagules or surviving on crop residues or weeds. The native geographic range for
D. anchilisposomus,
D. clarus, and
D. valveus of Western USA [
31,
40] and Kyrgyzstan for
D. tenuidens [
41] is now increased with the present records. The long-distance dispersal of these species is only possible through either phoretic association with insect vectors or transportation by humans and animals. There are reports of bark beetles and sweat bees acting as vectors for
Ditylenchus species [
10,
11] and a presumption that many soil-inhabiting nematodes were introduced to different locations by migratory bison and birds in mud adhered to the animals [
44,
55]. For the moment, we do not have evidence to support the dispersal theories for these
Ditylenchus species; we believe it is an area that warrants future research. Since the recovered species were found in moderate numbers (29–47 individuals in 250 g of soil), we do not consider them as pest species. However, it is crucial to monitor the quantities of
D. anchilisposomus,
D. clarus,
D. tenuidens, and
D. valveus under different crop rotation regimes, as Hajihassani et al. [
26] demonstrated that the severity of disease development in peas and beans is closely related to the number of
D. dipsaci infesting the plant.
Root lesion nematodes are dominating species in southern Alberta and have been reported from several cultivated areas [
56]. In the present study, we found
Ditylenchus species coexisting with root lesion, pin, stunt, and several other soil-inhabiting nematodes (members of Tylenchidae Orley [
57], rhabditids, and dorylaimids). Root lesion nematodes are the third, and stem nematodes are the fifth most damaging PPNs worldwide [
58]. Both these nematodes are migratory endoparasites; if left unchecked, they can affect plant growth and yields under elevated stress. In addition, the co-occurrence of several PPN groups can lay the foundation for a novel encounter between different nematode species and host plants, thus creating new disease complexes that may challenge host defenses [
59].
Ditylenchus penetration and movement within a plant tissue have been shown to cause mechanical injuries that interfere with the uptake of water and nutrients [
15]. Therefore, we emphasize the need for increased surveys and optimum agronomic practices to evaluate the PPN pre-planting numbers to prevent the subsequent spread of different PPN groups.
Recent phylogenetic studies do not concur on the paraphyletic status and presence of different lineages in
Ditylenchus [
10,
11,
60]. In our study, we noted that phylogenetic inference is effective for separating species, but with limited sequence-based information, these inferences are inconclusive in assigning species groups (
D.
dipsaci or
D.
triformis) or predicting the relationship among these species. Due to the unavailability of these important data, the lineage species concept cannot be applied sufficiently to
Ditylenchus phylogeny. Consequently, the typological species concept may continue to provide morphological insights, until new molecular data become available.
The major sources of soil contamination with PPNs in cultivated lands are from previous crops, weeds, or accidental introduction with infested or infected planting material from other locations [
3,
26,
37]. Therefore, accurate species identification is crucial, because false-positive identification can result in quarantining an entire production area, while a false negative can lead to the spread of the regulated species to other cultivated regions [
8]. In this context, our study provided detailed photo documentation and description of
D. anchilisposomus,
D. clarus,
D. tenuidens, and
D. valveus along with their first molecular characterization. The recovered
Ditylenchus species are new records for Canadian nematode fauna; they are non-regulated pests and have never been reported to cause any damage to plants. However, the economically unimportant nematode species that have remained in balance with plant hosts may emerge as parasites when species habitat, agronomic practices, choice of cultivar, or rotation cycles change. Hence, it is important to identify the often-overlooked nematode infestation issues, along with the continuous application of preventive crop protection and pest management measures.
5. Conclusions
There is a constant threat of the emergence of new nematode pest species. Crop rotation is considered an effective part of a management plan to mitigate some of these risks. Therefore, it is important to have good knowledge of the dominating nematode species inhabiting soil, particularly when it comes to migratory endoparasitic nematodes, such as
Ditylenchus, which spend most of their life cycle inside plant tissues. These nematodes can be spread unknowingly and escape timely detection. In the present study, we detected four
Ditylenchus species in no-till fields. This is the first record of any
D.
triformis-group species from southern Alberta, as well as their first molecular characterization. Our light micrographs and sequence-based information will enable the accurate and prompt identification of these species. In Canada, the research on the
Ditylenchus genus is centered on
D. dipsaci and
D. weischeri [
21,
26,
35]. The present work emphasizes the need for studying and documenting other
Ditylenchus species, since many nematodes that we currently consider as benign or nonparasitic may become pests due to changes in agricultural or environmental conditions.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}