
Biostimulant and Elicitor Responses to Cricket Frass (Acheta domesticus) in Tomato (Solanum lycopersicum L.) under Protected Conditions
 , , ,
, , , 
Abstract
:1. Introduction
2. Results
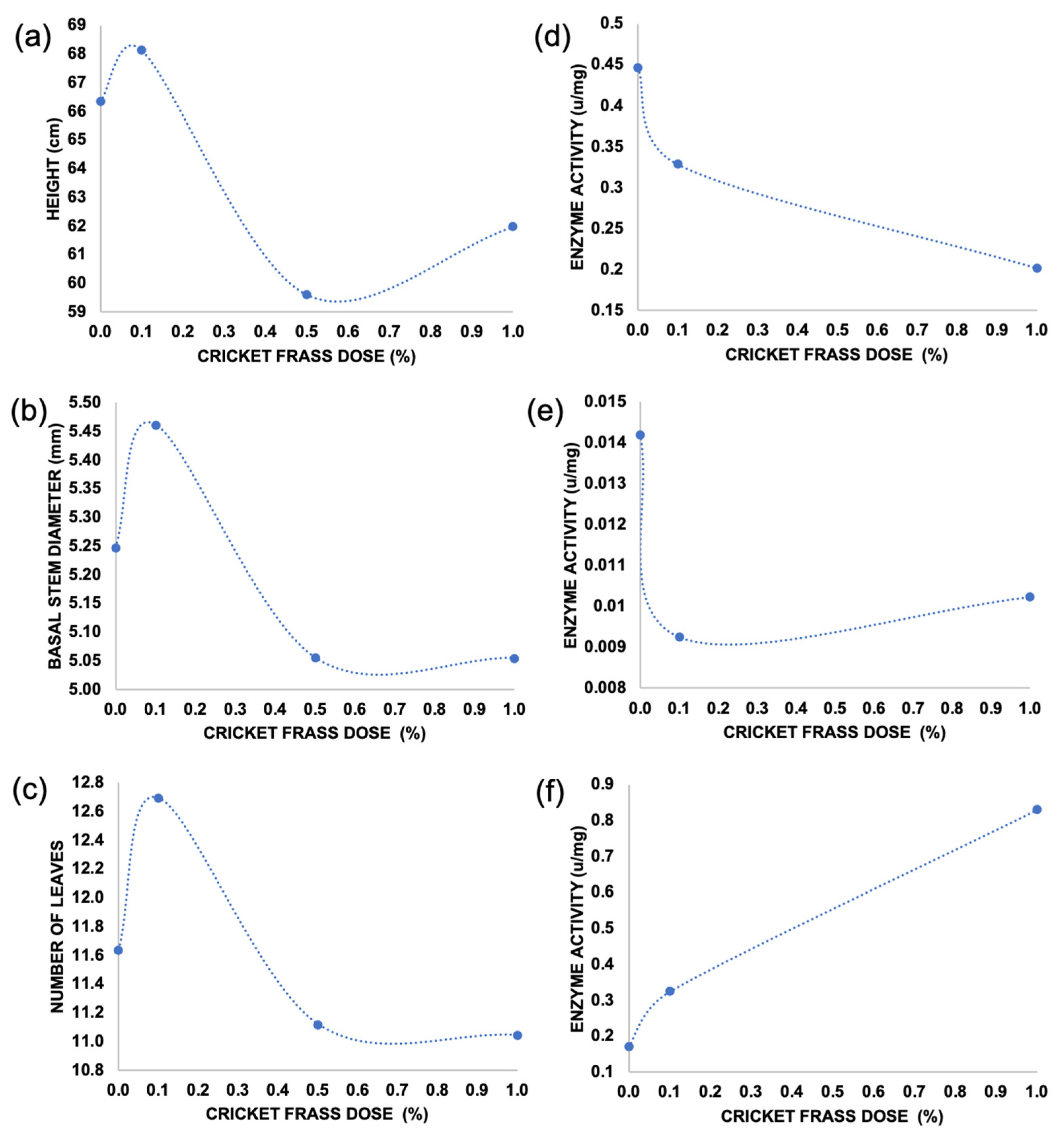
2.1. Effect of Cricket Frass on Plant Morphological Variables in Tomato
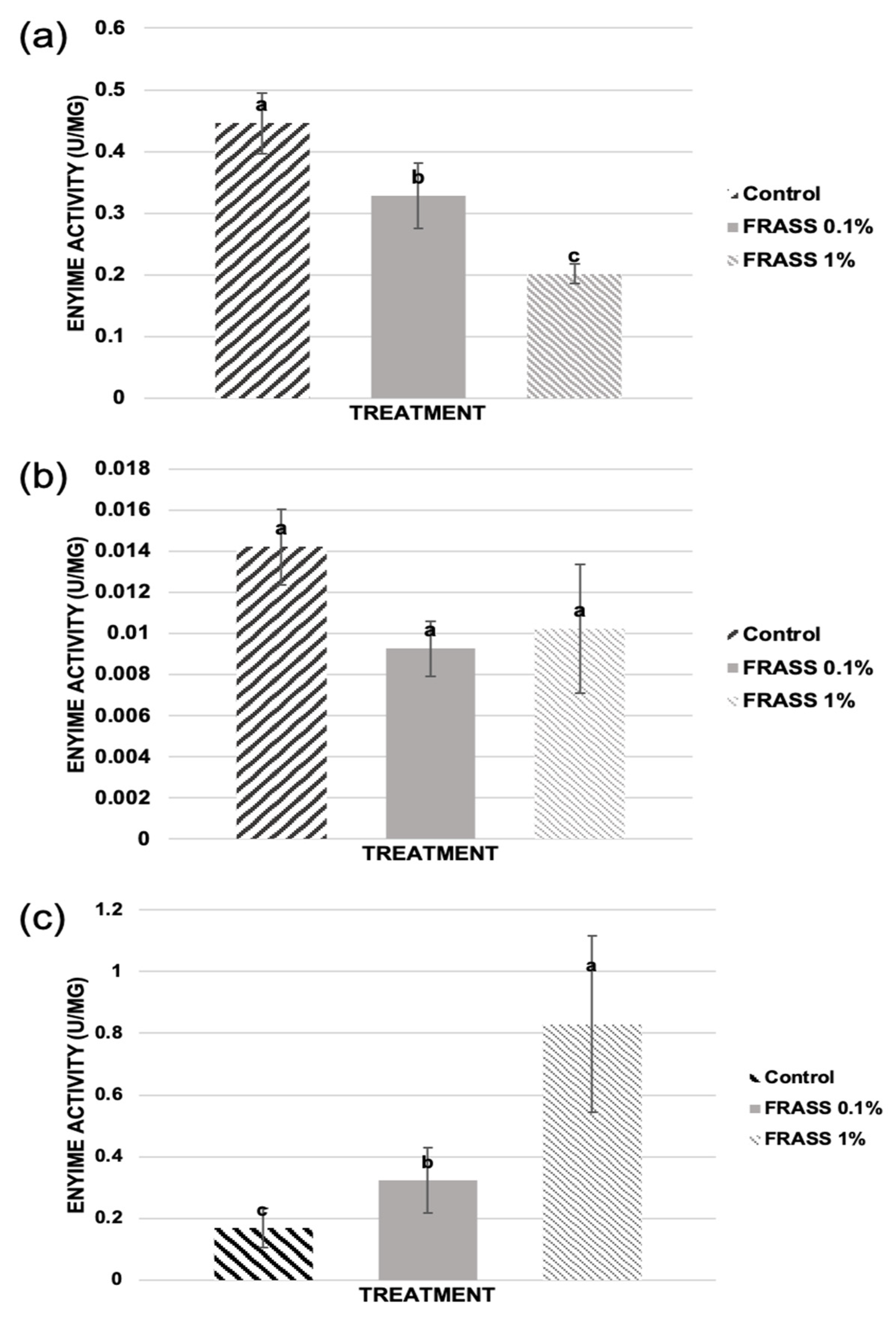
2.2. Effect of Cricket Frass on Plant Antioxidant Enzymatic Immunity Markers
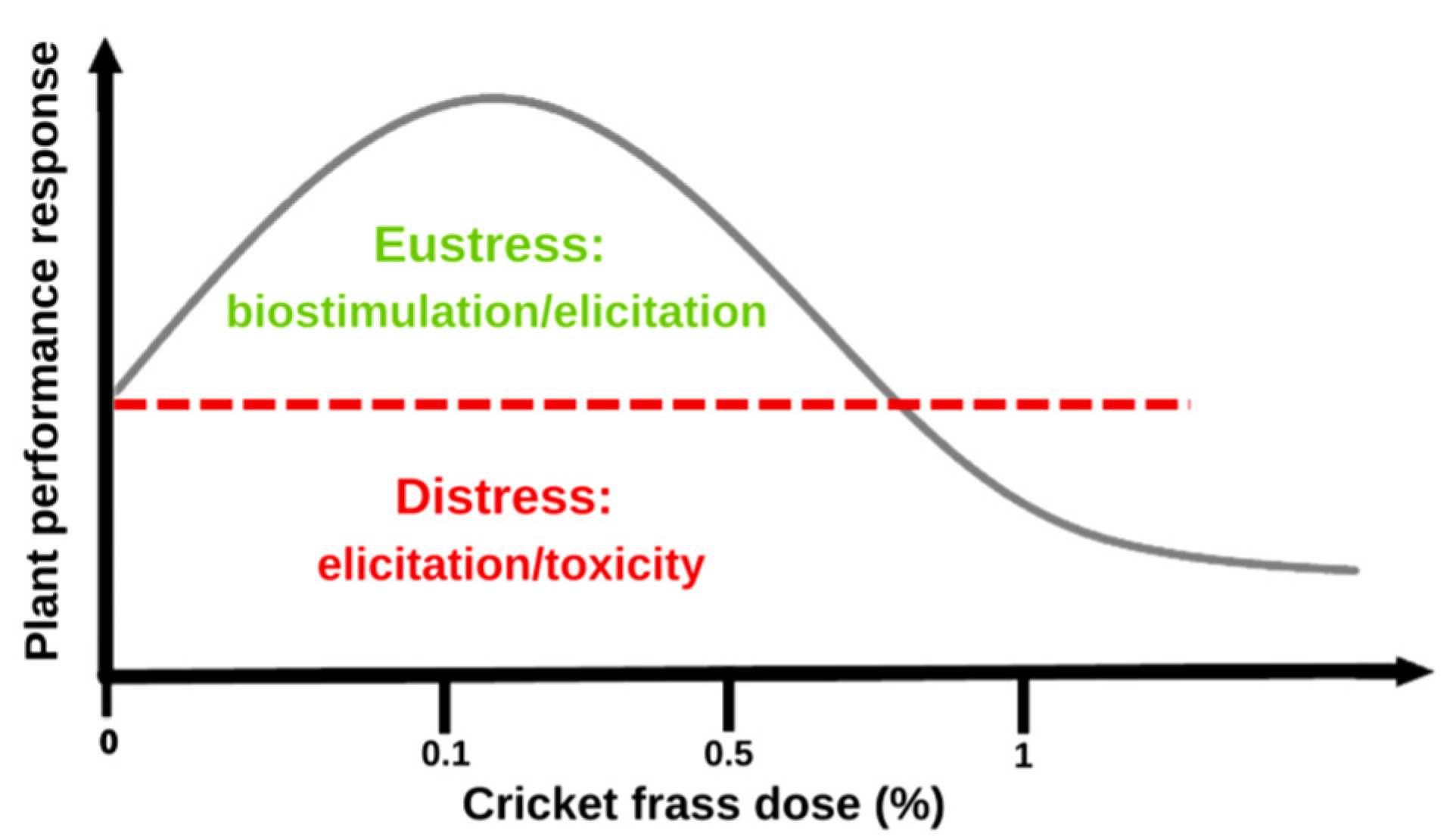
3. Discussion
4. Materials and Methods
4.1. Plant Growth Conditions and Fertigation
4.2. Frass Nutrient Characteristics
4.3. Treatments
4.4. Determination of Plant Morphological Characteristics
4.5. Antioxidant Enzyme Activities
4.5.1. Superoxide Dismutase (SOD) Enzyme Activity
4.5.2. Catalase (CAT) Enzyme Activity
4.5.3. Phenylalanine Ammonium Lyase (PAL) Activity
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vázquez-Hernández, M.C.; Parola-Contreras, I.; Montoya-Gómez, L.M.; Torres-Pacheco, I.; Schwarz, D.; Guevara-González, R.G. Eustressors: Chemical and physical stress factors used to enhance vegetables production. Sci. Hortic. 2019, 250, 223–229. [Google Scholar] [CrossRef]
- Vargas-Hernandez, M.; MAcias-Bobadilla, I.; Guevara-Gonzalez, R.G.; de Romero-Gomez, S.J.; Rico-Garcia, E.; Ocampo-Velazquez, R.V.; de Alvarez-Arquieta, L.L.; Torres-Pacheco, I. Plant hormesis management with biostimulants of biotic origin in agriculture. Front. Plant Sci. 2017, 8, 1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solano-Alvarez, N.; Valencia-Hernández, J.A.; Rico-García, E.; Torres-Pacheco, I.; Ocampo-Velázquez, R.V.; Escamilla-Silva, E.M.; Romero-García, A.L.; Alpuche-Solís, Á.G.; Guevara-González, R.G. A novel isolate of bacillus cereus promotes growth in tomato and inhibits clavibacter michiganensis infection under greenhouse conditions. Plants 2021, 10, 506. [Google Scholar] [CrossRef] [PubMed]
- Mejía-Teniente, L.; Durán-Flores, B.A.; Torres-Pacheco, I.; González-Chavira, M.M.; Rivera-Bustamante, R.F.; Feregrino-Perez, A.A.; Pérez-Ramírez, I.; Rocha-Guzmán, N.E.; Reynoso-Camacho, R.; Guevara-González, R.G. Hydrogen peroxide protects pepper (Capsicum annuum L.) against pepper golden mosaic geminivirus (PepGMV) infections. Physiol. Mol. Plant Pathol. 2019, 106, 23–29. [Google Scholar] [CrossRef]
- Díaz, G.; Rodríguez, G.; Montana, L.; Miranda, T.; Basso, C.; Arcia, M. Efecto de la aplicación de bioestimulantes y Trichoderma sobre el crecimiento en plántulas de maracuyá (Passiflora edulis Sims) en vivero. Bioagro 2020, 32, 195–204. [Google Scholar]
- Crouch, I.J.; Beckett, R.P.; van Staden, J. Effect of seaweed concentrate on the growth and mineral nutrition of nutrient-stressed lettuce. J. Appl. Phycol. 1990, 2, 269–272. [Google Scholar] [CrossRef]
- Sadiqi, S.; Hamza, M.; Ali, F.; Alam, S.; Shakeela, Q.; Ahmed, S.; Ayaz, A.; Ali, S.; Saqib, S.; Ullah, F.; et al. Molecular Characterization of Bacterial Isolates from Soil Samples and Evaluation of their Antibacterial Potential against MDRS. Molecules 2022, 27, 6281. [Google Scholar] [CrossRef]
- Pereira, J.F.; Oliveira, A.L.M.; Sartori, D.; Yamashita, F.; Mali, S. Perspectives on the Use of Biopolymeric Matrices as Carriers for Plant-Growth Promoting Bacteria in Agricultural Systems. Microorganisms 2023, 11, 467. [Google Scholar] [CrossRef]
- El Amerany, F.; Meddich, A.; Wahbi, S.; Porzel, A.; Taourirte, M.; Rhazi, M.; Hause, B. Foliar application of chitosan increases tomato growth and influences mycorrhization and expression of endochitinase-encoding genes. Int. J. Mol. Sci. 2020, 21, 535. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.M.N.V. A review of chitin and chitosan applications. React. Funct. Polym. 2000, 46, 1–27. [Google Scholar] [CrossRef]
- Antony, R.; Arun, T.; Manickam, S.T.D. A review on applications of chitosan-based Schiff bases. Int. J. Biol. Macromol. 2019, 129, 615–633. [Google Scholar] [CrossRef] [PubMed]
- Saqib, S.; Zaman, W.; Ayaz, A.; Habib, S.; Bahadur, S.; Hussain, S.; Muhammad, S.; Ullah, F. Postharvest disease inhibition in fruit by synthesis and characterization of chitosan iron oxide nanoparticles. Biocatal. Agric. Biotechnol. 2020, 28, 101729. [Google Scholar] [CrossRef]
- Van Huis, A. Insects as food and feed, a new emerging agricultural sector: A review. J. Insects Food Feed 2020, 6, 27–44. [Google Scholar] [CrossRef] [Green Version]
- Cortes Ortiz, J.A.; Ruiz, A.; Morales-Ramos, J.; Thomas, M.; Rojas, M.G.; Tomberlin, J.K.; Yi, L.; Han, R.; Giroud, L.; Jullien, R.L. Chapter 6—Insect Mass Production Technologies. In Insects as Sustainable Food Ingredients: Production, Processing and Food Applications; Dossey, A.T., Morales-Ramos, J.A., Rojas, M.G.B.T., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 153–201. [Google Scholar] [CrossRef]
- Wang, H.; Rehman, K.; Liu, X.; Yang, Q.; Zheng, L.; Li, W.; Cai, M.; Li, Q.; Zhang, J.; Yu, Z. Biotechnology for Biofuels Insect biorefinery: A green approach for conversion of crop residues into biodiesel and protein. Biotechnol. Biofuels 2017, 10, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbajal-Valenzuela, I.A.; Guzmán-Cruz, R.; González-Chavira, M.M.; Medina-Ramos, G.; Serrano-Jamaica, L.M.; Torres-Pacheco, I.; Vázquez, L.; Feregrino-Pérez, A.A.; Rico-García, E.; Guevara-González, R.G. Response of Plant Immunity Markers to Early and Late Application of Extracellular DNA from Different Sources in Tomato (Solanum lycopersicum). Agriculture 2022, 12, 1587. [Google Scholar] [CrossRef]
- Ferrusquía-Jiménez, N.I.; González-Arias, B.; Rosales, A.; Esquivel, K.; Escamilla-Silva, E.M.; Ortega-Torres, A.E.; Guevara-González, R.G. Elicitation of Bacillus cereus-Amazcala (B.c-A) with SiO2 Nanoparticles Improves Its Role as a Plant Growth-Promoting Bacteria (PGPB) in Chili Pepper Plants. Plants 2022, 11, 3445. [Google Scholar] [CrossRef]
- Magaña-López, E.; Palos-Barba, V.; Zuverza-Mena, N.; Vázquez-Hernández, M.C.; White, J.; Nava-Mendoza, R.; Feregrino-Pérez, A.A.; Torres-Pacheco, I.; Guevara-González, R.G. Nanostructured mesoporous silica materials induce hormesis on chili pepper (Capsicum annuum L.) under greenhouse conditions. Heliyon 2022, 8, e09049. [Google Scholar] [CrossRef]
- Vargas-Hernandez, M.; Ferrusquia-Jimenez, N.I. Introducción a la teoría del estrés e inmunología vegetal. In Manejo del Estrés Vegetal como una Estrategia para una Agricultura Sostenible; Guevara-González, R.G., Torres-Pacheco, I., Eds.; Editorial Universidad de Almería: Almería, Spain, 2022; pp. 53–101. [Google Scholar]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodge, S.; Conway, J. The Effects of Insect Frass Fertilizer and Biochar on the Shoot Growth of Chicory and Plantain, Two Forage Herbs Commonly Used in Multispecies Swards. Agronomy 2022, 12, 2459. [Google Scholar] [CrossRef]
- Poveda, J.; Jiménez-Gómez, A.; Saati-Santamaría, Z.; Usategui-Martín, R.; Rivas, R.; García-Fraile, P. Mealworm frass as a potential biofertilizer and abiotic stress tolerance-inductor in plants. Appl. Soil Ecol. 2019, 142, 110–122. [Google Scholar] [CrossRef]
- Ray, S.; Alves, P.C.M.S.; Ahmad, I.; Gaffoor, I.; Acevedo, F.E.; Peiffer, M.; Jin, S.; Han, Y.; Shakee, S.; Felton, G.W.; et al. Turnabout is fair play: Herbivory-induced plant chitinases excreted in fall armyworm frass suppress herbivore defenses in maize. Plant Physiol. 2016, 171, 694–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo, F.E.; Peiffer, M.; Tan, C.W.; Stanley, B.A.; Stanley, A.; Wang, J.; Jones, A.G.; Hoover, K.; Rosa, C.; Luthe, D.; et al. Fall armyworm-associated gut bacteria modulate plant defense responses. Mol. Plant-Microbe Interact. 2017, 30, 127–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozo, J.; Juan, E.A.; Morales, I.; Requena, J.; La Malfa, T. A New Local Sustainable Inorganic Material for Soilless Culture in Spain: Granulated Volcanic Rock. HortScience 2014, 49, 1537–1541. [Google Scholar] [CrossRef]
- Sonneveld, C.; Straver, N. Nutrient solutions for vegetables and flowers grown in water or substrates. Voedingsoploss. Glastuinb 1994, 10, 45. Available online: http://edepot.wur.nl/237302%0Ahttps://agris.fao.org/agris-search/search.do?recordID=NL8901262 (accessed on 1 April 2022).
- Da Cunha-Chiamolera, T.P.L.; Urrestarazu, M.; Filho, A.B.C.; Morales, I. Agronomic and economic feasibility of tomato and lettuce intercropping in a soilless system as a function of the electrical conductivity of the nutrient solution. HortScience 2017, 52, 1195–1200. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, D.; Reca, J.; Martínez, J.; Lao, M.T.; Urrestarazu, M. Effect of controlling the leaching fraction on the fertigation and production of a tomato crop under soilless culture. Sci. Hortic. 2014, 179, 153–157. [Google Scholar] [CrossRef]
- Urrestarazu, M.; Gallegos, V.; Álvaro, J.E. The Use of Thermography Images in the Description of the Humidification Bulb in Soilless Culture. Commun. Soil Sci. Plant Anal. 2017, 48, 1595–1602. [Google Scholar] [CrossRef] [Green Version]
- Sáenz-de la, O.D.; Morales, L.O.; Strid, Å.; Torres-Pacheco, I.; Guevara-González, R.G. Ultraviolet-B exposure and exogenous hydrogen peroxide application lead to cross-tolerance toward drought in Nicotiana tabacum L. Physiol. Plants 2021, 173, 666–679. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Ahmad, H.; Ali, M.; Ren, K.; Cheng, Z. Aqueous garlic extract stimulates growth and antioxidant enzymes activity of tomato (Solanum lycopersicum). Sci. Hortic. 2018, 240, 139–146. [Google Scholar] [CrossRef]
- Afiyanti, M.; Chen, H.J. Catalase activity is modulated by calcium and calmodulin in detached mature leaves of sweet potato. J. Plant Physiol. 2014, 171, 35–47. [Google Scholar] [CrossRef]
- Toscano, S.; Ferrante, A.; Leonardi, C.; Romano, D. PAL activities in asparagus spears during storage after ammonium sulfate treatments. Postharvest. Biol. Technol. 2018, 140, 34–41. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphologic Variables | Week | Control | 0.1% Frass | 0.5% Frass | 1% Frass |
|---|---|---|---|---|---|
| Height | 1 | 14 a | 15 a | 14 a | 15 a |
| 2 | 19 a | 20 a | 19 a | 20 a | |
| 3 | 26 a | 29 a | 24 a | 26 a | |
| 4 | 34 a,b | 41 a | 32 b | 32 b | |
| 5 | 40 a,b | 46 a | 38 a,b | 37 b | |
| 6 | 44 a,b | 51 a | 42 b | 42 b | |
| 7 | 58 a | 60 a | 51 a | 53 a | |
| 8 | 66 a | 68 a | 60 a | 62 a | |
| Stem diameter | 1 | 3.24 a | 3.64 a | 3.33 a | 3.45 a |
| 2 | 3.29 a | 3.69 a | 3.38 a | 3.50 a | |
| 3 | 3.33 a | 3.73 a | 3.42 a | 3.54 a | |
| 4 | 4.16 a,b | 4.58 a | 4.02 b | 4.11 b | |
| 5 | 4.22 a,b | 4.64 a | 4.07 b | 4.18 ab | |
| 6 | 5.02 a | 5.23 a | 4.82 a | 4.82 a | |
| 7 | 5.12 a | 5.33 a | 4.92 a | 4.92 a | |
| 8 | 5.25 a | 5.46 a | 5.05 a | 5.05 a | |
| Number of leaves | 1 | 3 a | 4 a | 3 a | 3 a |
| 2 | 3 a | 4 a | 3 a | 3 a | |
| 3 | 4 a | 5 a | 4 a | 4 a | |
| 4 | 7 a,b | 8 a | 6 b | 6 b | |
| 5 | 9 a,b | 10 a | 8 b | 8 b | |
| 6 | 7 a | 8 a | 6 a | 6 a | |
| 7 | 10 a | 11 a | 9 a | 9 a | |
| 8 | 12 a | 13 a | 11 a | 11 a |
| Nutrients | Element | Units | Results | Method |
|---|---|---|---|---|
| Macronutrients | Total nitrogen | % | 4.035 | MicroKjeldahl |
| Total Phosphorus (P2O5) | % | 1.560 | Spectrophotometric | |
| Potassium (K2O) | % | 1.820 | Atomic absorption | |
| Secondary elements | Calcium | % | 1.340 | Atomic absorption |
| Magnesium | % | 0.510 | Atomic absorption | |
| Sulfur | % | 0.699 | Turbidimetric | |
| Sodium | % | 0.620 | Atomic absorption | |
| Micronutrients | Iron | ppm | 334.040 | Atomic absorption |
| Cooper | ppm | 47.770 | Atomic absorption | |
| Manganese | ppm | 154.350 | Atomic absorption | |
| Zinc | ppm | 195.680 | Atomic absorption |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferruzca-Campos, E.A.; Rico-Chavez, A.K.; Guevara-González, R.G.; Urrestarazu, M.; Cunha-Chiamolera, T.P.L.; Reynoso-Camacho, R.; Guzmán-Cruz, R. Biostimulant and Elicitor Responses to Cricket Frass (Acheta domesticus) in Tomato (Solanum lycopersicum L.) under Protected Conditions. Plants 2023, 12, 1327. https://doi.org/10.3390/plants12061327
Ferruzca-Campos EA, Rico-Chavez AK, Guevara-González RG, Urrestarazu M, Cunha-Chiamolera TPL, Reynoso-Camacho R, Guzmán-Cruz R. Biostimulant and Elicitor Responses to Cricket Frass (Acheta domesticus) in Tomato (Solanum lycopersicum L.) under Protected Conditions. Plants. 2023; 12(6):1327. https://doi.org/10.3390/plants12061327
Chicago/Turabian StyleFerruzca-Campos, Ema Alejandra, Amanda Kim Rico-Chavez, Ramón Gerardo Guevara-González, Miguel Urrestarazu, Tatiana Pagan Loeiro Cunha-Chiamolera, Rosalía Reynoso-Camacho, and Rosario Guzmán-Cruz. 2023. "Biostimulant and Elicitor Responses to Cricket Frass (Acheta domesticus) in Tomato (Solanum lycopersicum L.) under Protected Conditions" Plants 12, no. 6: 1327. https://doi.org/10.3390/plants12061327