Abstract
Tree peony is a “spring colored-leaf” plant which has red leaves in early spring, and the red color of the leaves usually fades in late spring. Flavonols are one subgroup of flavonoids, and they affect the plant organs’ color as co-pigments of anthocyanins. To investigate the color variation mechanism of leaves in tree peony, PqMYBF1, one flavonol biosynthesis-related MYB gene was isolated from Paeonia qiui and characterized. PqMYBF1 contained the SG7 and SG7-2 motifs which are unique in flavonol-specific MYB regulators. Subcellular localization and transactivation assay showed that PqMYBF1 localized to the nucleus and acted as a transcriptional activator. The ectopic expression of PqMYBF1 in transgenic tobacco caused an observable increase in flavonol level and the anthocyanin accumulation was decreased significantly, resulting in pale pink flowers. Dual-luciferase reporter assays showed that PqMYBF1 could activate the promoters of PqCHS, PqF3H, and PqFLS. These results suggested that PqMYBF1 could promote flavonol biosynthesis by activating PqCHS, PqF3H, and PqFLS expression, which leads metabolic flux from anthocyanin to flavonol pathway, resulting in more flavonol accumulation. These findings provide a new train of thought for the molecular mechanism of leaf color variation in tree peony in spring, which will be helpful for the molecular breeding of tree peony with colored foliage.
1. Introduction
Leaf color is one of the attractive traits for ornamental plants, and plants with colored foliage are often called “colored-leaf plants”, including spring colored-leaf plants, autumn colored-leaf plants, and colored-leaf plants all year round. Among them, spring colored-leaf plants can be used in landscape construction in early spring. The red leaf color formation in early spring and the red color fading of leaf in late spring are two remarkable events in spring colored-leaf plants, such as tree peony and crabapple.
Studies have shown that the color variation from red to green is caused by the change of pigments inside the leaves. It is well known that red color usually arises from anthocyanins (one subgroup of flavonoids). For example, Tang et al. reported that the anthocyanin content gradually decreases during red fading in herbaceous peony leaves [1]. Decreasing anthocyanin content led to fading red color in Malus ‘Radiant’ leaves [2]. Flavonols, another subgroup of flavonoids, can also modulate plant organ color as co-pigments of anthocyanins [3,4].
As a branch of flavonoid biosynthesis, the biosynthesis of flavonol has been studied in Arabidopsis, Vitis vinifera, Malus crabapple, and other species [5,6,7,8,9,10]. First, chalcone synthase (CHS) catalyzes coumaroyl-CoA and malonyl-CoA to form chalcone, which is isomerized to naringenin by chalcone isomerase (CHI). Then, naringenin is converted to dihydroflavonols by flavanone-3-hydrolase (F3H) and flavonoid 3′-hydroxylase (F3′H). Subsequently, dihydroflavonols can be converted to anthocyanins by dihydroflavonol 4-reductase (DFR) and anthocyanidin synthase (ANS). At the same time, flavonols can also be formed from didydroflavonols by flavonol synthase (FLS). The competition relationship between FLS and DFR directly affects the accumulation of flavonols and anthocyanins, which in turn affects plant coloration [4,11,12,13].
In addition, the biosynthesis of flavonol is also controlled by a complex regulatory network, the most important of which is the MYB regulator [14,15,16]. In A. thaliana, R2R3-MYBs are divided into 25 subfamilies according to the differences in C-terminal. MYBs in subgroup SG4, SG5, SG6, SG7, and SG15 are involved in regulating the synthesis of flavonoids [17,18]. MYB members belonging to the SG7 group play a significant role in the synthesis of flavonol [19]. In recent years, R2R3-MYB transcriptional activators for flavonol synthesis have been isolated from Gerbera hybrida, Medicago truncatula, Malus sieversii, Fagopyrum tataricum, and Freesia hybrida [20,21,22,23,24]. For example, the pear PbMYB12b protein positively regulates the accumulation of flavonol by promoting the expression of PbCHSb and PbFLS [25]; the overexpression of Mt134 from M. truncatula in Arabidopsis complements the flavonol deficiency [23]. In addition to transcriptional activators, researchers have also isolated repressors that inhibited flavonol biosynthesis, such as FaMYB1, from strawberry (Fragaria × ananassa) [26]. In Chrysanthemum × morifolium, CmMYB012 inhibits flavonol biosynthesis by inactivating the transcriptional activity of CmCHS [27]. However, most of these studies were conducted in flowers or fruits; the regulation mechanism of flavonol biosynthesis in spring colored-leaf plants is not clear.
Tree peony is one of the top ten Chinese traditional flowers and is known as the “king of flowers” for its gorgeous and magnificent flowers. At the same time, tree peony is also a “spring red leaf” plant. Its young leaves are purplish red in early spring, and the red color of the leaves usually fades in late spring [28]. Among all kinds of tree peony, the leaf coloring trait of Paeonia qiui is the most typical. In P. qiui, the high level of anthocyanins rapidly accumulates in red young leaves, after which anthocyanin level decreases and other flavonoids (including flavonol) increase with leaf development [29]. In our previous studies, some flavonoid biosynthesis-related R2R3-MYBs were found in P. qiui through transcriptome sequencing [30]. Among them, two R2R3-MYBs (PqMYB113 and PqMYB4) were isolated, and their biological functions were verified in anthocyanin biosynthesis [31,32]. In this study, another flavonoid (flavonol) biosynthesis activator, PqMYBF1 from the leaves of P. qiui, was characterized, and the biological function of PqMYBF1 was investigated using genetically modified tobacco, which will enrich our knowledge about tree peony leaf color variation in spring.
2. Results
2.1. PqMYBF1 Is a Potential Regulator of Flavonol Biosynthesis
Phylogenetic analysis showed that PqMYBF1 was placed into the subgroup 7 (SG7) family proteins which are flavonol-specific R2R3-MYB transcription factors (TFs), such as grape VvMYBF1, apple MdMYB22, tomato SlMYB12, Arabidopsis AtMYB11, AtMYB12, and AtMYB111. PqMYBF1 shared 44.29% amino acid sequence identity with grape VvMYBF1, 35.95% with Arabidopsis AtMYB12 and 35.48% with apple MdMYB22 (Figure 1A). Based on these results, we speculated that PqMYBF1 was probably involved in the regulation of flavonol synthesis of leaves in P. qiui.

Figure 1.
Phylogenetic analysis and sequence alignment of PqMYBF1 with other MYB proteins (A) Phylogenetic analysis of PqMYBF1 protein and MYB proteins from other species. Presumed functions of these MYB proteins are listed on the right side of the phylogenetic tree. PqMYBF1 is highlighted with a red dot. The neighbor-joining method with MEGA software was used to construct the phylogenic tree. Bootstrap values as a percentage of 1000 replicates are indicated at corresponding branch nodes. Scale bar represents the number of amino acid substitutions per site. (B) Sequence alignment of PqMYBF1 with MYB involved in flavonol synthesis. The black line indicates the positions of the R2 and R3 MYB domains. The red box shows the SG7 and SG7-2 motif.
The ORF of PqMYBF1 was 1140 bp encoding a protein of 379 amino acid residues. Multiple sequence alignment analysis revealed that PqMYBF1 contains the typical SANT domain of the MYB family at the N-terminus (Figure 1B). The SG7 motif ([K/R][R/x][R/K]xGRT[S/x][R/G]xx[M/x]K) and SG7-2 motif ([W/x][L/x]LS) which are the characteristics of flavonol biosynthesis regulators, were also found at the C-terminus of PqMYBF1 (Figure 1B) [7,8,19,33,34]. PqMYBF1 did not contain the motif [D/E]Lx2[R/K]x3Lx6Lx3R that interacts with bHLH proteins, suggesting that PqMYBF1 functions independently without bHLH [35].
2.2. PqMYBF1 Localizes to the Nucleus and Acts as a Transcriptional Activator
To investigate the subcellular localization of PqMYBF1, the recombinant vector pCAMBIA2300-PqMYBF1-GFP was transformed into onion epidermis. Red fluorescence showed the location of the nucleus. Onion cell expressing the PqMYBF1-GFP fusion protein showed a strong fluorescent signal in the nucleus. Therefore, we speculated that PqMYBF1 was localized and functioned in the nucleus (Figure 2A).
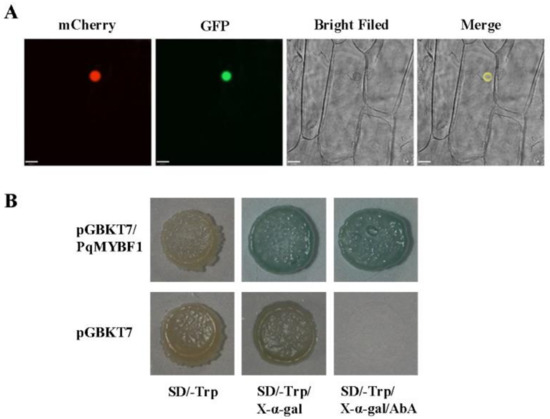
Figure 2.
Subcellular localization and transcriptional activity of PqMYBF1. (A) Subcellular location of GFP fusion of PqMYBF1. The mCherry protein indicates nucleus localization. Bars = 33 µm. (B) Transcriptional activity of PqMYBF1 in yeast. Y2H transformed with pGBKT7-PqMYBF1 or pGBKT7 vector was grown on SD/-Trp, SD/-Trp with 40 µg/mL X-α-gal and SD/-Trp with 40 µg/mL X-α-gal adding 200 ng/mL aureobasidin A (AbA) for three days.
A transcriptional activity test was performed in Y2H yeast using a recombinant vector pGBKT7-PqMYBF1 carrying the GAL DNA-domain to determine whether PqMYBF1 has transcriptional activation activity. Yeast carrying pGBKT7-PqMYBF1 grew normally and appeared blue on SD/-Trp with X-α-gal adding aureobasidin A (AbA). The negative control group showed an inhibition of colony growth (Figure 2B). These indicated that PqMYBF1 has transcriptional activation activity.
2.3. PqMYBF1 Expression Correlates with Flavonol Accumulation and Flavonol Biosynthetic Gene Expression
The expression levels of PqMYBF1 and flavonoid biosynthetic genes in different leaf stages were revealed by qRT-PCR. The content of anthocyanins and flavonols were measured (Figure 3). The total anthocyanin content of P. qiui leaves increased first, peaked at S3, and then decreased mildly in S4 and declined dramatically in S5, which was consistent with the observed phenotype (Figure 3B). For flavonol content, the results showed that there was a slight increased trend from S1 to S3, and then reduced slightly at S4 and S5, which overlapped with that of anthocyanin, but was not the same. The flavonol content was the highest in S3 stage, followed by S4, S5, and S2; the lowest was in S1 (Figure 3C).
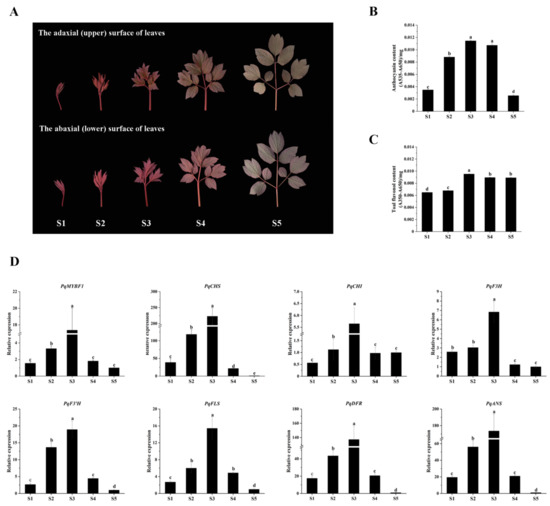
Figure 3.
Leaf phenotypes in different states of Paeonia qiui and expression level of flavonoid structural genes and PqMYBF1 at different stages in P. qiui. (A) The adaxial (upper) and abaxial (lower) surface of P. qiui leaves at different stages. The leaf development was divided into five stages mainly based on leaf status and pigmentation: S1: the leaves curled up; S2: the leaves extended slightly; S3: the leaves basically unfolded; S4: the leaves unfolded completely; S5: the leaves turned green. (B) Anthocyanin content in P. qiui. (C) Total flavonol content in P. qiui. (D) The expression level of PqMYBF1, PqCHS, PqCHI, PqF3H, PqF3′H, PqFLS, PqDFR, and PqANS. a, b, c, and d indicate significant differences at the p ≤ 0.05 level in the Duncan test.
As for the expression levels of PqMYBF1 (Figure 3D), the results showed that it increased first and then decreased. PqMYBF1 had the highest expression level in the S3 stage. In addition, the expression levels of flavonoid biosynthetic genes were also analyzed. The result showed that PqCHS, PqCHI, PqF3H, PqF3’H, PqFLS, PqDFR, and PqANS genes presented a basically consistent trend, which increased first and then decreased. The maximum value of their expression was in S3 stage. The expression levels of PqCHI were the lowest in S1, while the expression levels of PqCHS, PqF3H, PqF3’H, PqFLS, PqDFR, and PqANS were the lowest in the S5 (Figure 3D).
2.4. Overexpression of PqMYBF1 Promoted Flavonol Accumulation and the Expression of Flavonol Pathway Genes in Tobacco
To further investigate the function of PqMYBF1, the gene was cloned into an overexpression vector driven by 35S promoter and overexpressed in tobacco. Thirteen overexpression (OE) transgenic lines were generated (Figure 4A). Three transgenic lines (OE-6, OE-8 and OE-11) were selected for subsequent experiments. The flowers of three transgenic tobacco lines showed pale pink color, whereas the wild type (WT) lines showed red-pink flowers (Figure 4B). Then, the level of total flavonols and anthocyanins in transgenic and the WT petals were quantified. The total flavonol contents of the transgenic lines were significantly higher by 1.68- (OE-6), 1.57- (OE-8) and 3.17-fold (OE-11) than those in the WT. However, the anthocyanin contents of the transgenic plants were decreased by 71.16% (OE-6), 60.17% (OE-8) and 86.97% (OE-11) compared with the WT (Figure 4C). qRT-PCR was performed to evaluate the expression of anthocyanin and flavonol biosynthesis pathway genes in transgenic tobacco flowers. PqMYBF1 was overexpressed in transgenic tobacco flowers and absent in the WT. Compared with the WT, the expression levels of NtCHS, NtF3H, NtF3′H, and NtFLS in transgenic tobacco flowers were higher, especially NtCHS, NtF3H, and NtFLS (Figure 4D). However, the expression levels of NtDFR and NtANS decreased slightly compared with the WT.
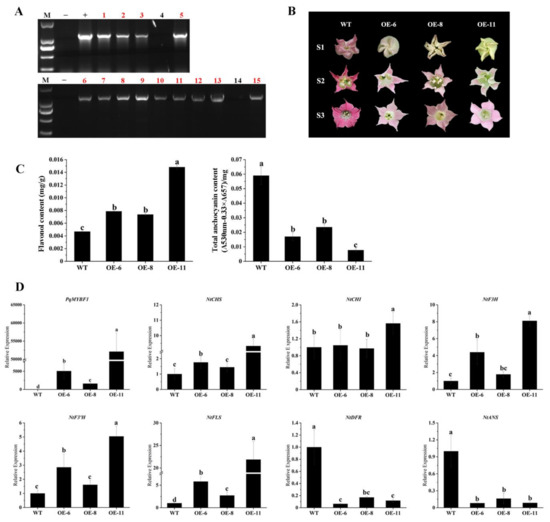
Figure 4.
The phenotype and the effect of PqMYBF1 overexpression in transgenic tobacco. (A) PCR analysis of PqMYBF1 transformed tobaccos. M: Trans2K Plus DNA Marker; −: Negative control; +: Positive control; 1–15: PCR identification of tobacco; the red numbers indicate the transgenic tobaccos. (B) The petal phenotypes of transgenic and wild tobaccos at three flowering stages. S1: closed buds, S2: initially open flowers, S3: open flowers. (C) The anthocyanin and flavonol content in petals. (D) The expression profiles of vital anthocyanin and flavonol biosynthesis pathway genes in PqMYBF1 overexpression transgenic tobacco petals. a, b, c, and d indicate significant differences at the p ≤ 0.05 level in the Duncan test.
We also investigated the influence of PqMYBF1 on flavonol synthesis in tobacco leaves. As shown in Figure 5, the flavonol content of tobacco leaves in OE-6, OE-8, and OE-11 were about 3.4, 2.0, 3.6 times those in wild type leaves, respectively. We quantified the flavonol-related genes expression of transgenic and wild type tobacco leaves. PqMYBF1 gene was highly expressed in transgenic tobacco lines leaves and had little or no expression in the WT. The expression levels of NtCHS, NtCHI, NtF3H, and NtFLS in transgenic tobacco leaves were much higher than those in the WT, especially NtCHS (up-regulated by 8.24-fold in OE-6, 5.83-fold in OE-8, and 27.5-fold in OE-11), NtF3H (up-regulated by 7.68-fold in OE-6, 4.69-fold in OE-8, and 51.71-fold in OE-11), and NtFLS (up-regulated by14.67-fold in OE-6, 12.57-fold in OE-8, and 43.57-fold in OE-11) (Figure 5). All of these suggested that PqMYBF1 could promote flavonol biosynthesis and accumulation in transgenic tobacco.
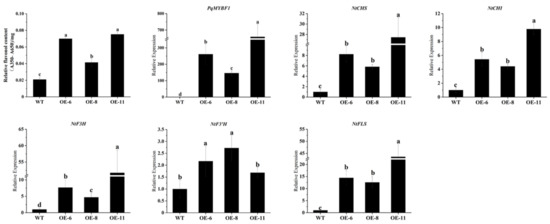
Figure 5.
The expression profiles of vital flavonol biosynthesis pathway genes in PqMYBF1 overexpression transgenic tobacco leaves. a, b, c, and d indicate significant differences at the p ≤ 0.05 level in the Duncan test.
2.5. PqMYBF1 Activated the Promoters of PqCHS, PqF3H, and PqFLS
The expressions of NtCHS, NtF3H, and NtFLS genes were more strongly increased than other genes of the flavonol pathway in PqMYBF1-overexpressing tobacco lines. We speculated that PqCHS, PqF3H, and PqFLS were potential targets of PqMYBF1 transcriptional activation. A dual-luciferase reporter assay was carried out to determine the interaction of PqMYBF1 with PqCHS, PqF3H, and PqFLS. The promoter sequences of PqCHS, PqF3H, and PqFLS from P. qiui were cloned and named as pPqCHS, pPqF3H, and pPqFLS, and the length of them were 2016 bp, 1624 bp, and 1488 bp, respectively.
The key cis-elements of pPqCHS, pPqF3H, and pPqFLS analysis were carried out using the PlantCARE online software and MYB binding sites were found in all of these three promoters (Figure 6A). These results indicated that pPqCHS, pPqF3H, and pPqFLS may be subject to MYB regulation. These three promoters were cloned into pBI121 vector with GUS reporter gene. GUS staining results showed that tobacco leaves treated with the fusion expression vectors and pBI121-pPqCHS-GUS, pBI121-pPqF3H-GUS, and pBI121-pPqFLS-GUS showed different degrees of staining, indicating that the promoters of PqCHS, PqF3H, and PqFLS genes have biological activity (Figure 6B).
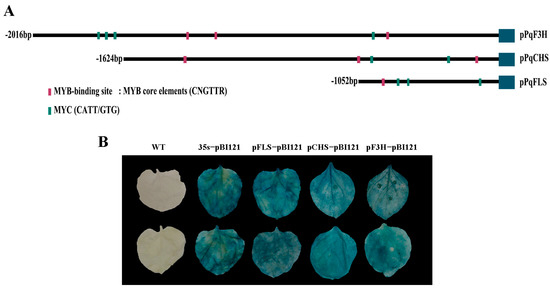
Figure 6.
Promoter analysis of flavonol synthesis-related gene. (A) The distribution of MYB binding elements in the promoter sequences of PqCHS, PqF3H, and PqFLS. (B) GUS activity analysis of promoters from PqCHS, PqF3H, and PqFLS.
Dual-luciferase reporter assay revealed that the promoter luminescence intensities of PqCHS, PqF3H, and PqFLS were all significantly increased by PqMYBF1 compared with the corresponding controls (Figure 7). This indicated that PqMYBF1 could activate the promoters of PqCHS, PqF3H, and PqFLS.
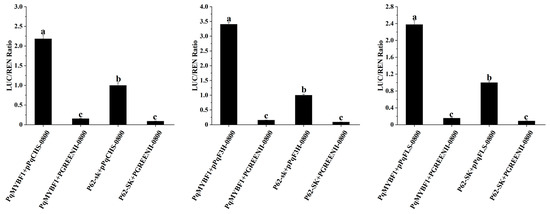
Figure 7.
PqMYBF1 interacts with the flavonol-related synthesis gene in vivo. PqMYBF1 induced the expression of PqCHS, PqF3H, and PqFLS. The activities of promoters were indicated by the ratio of LUC/REN. Three independent experiments were performed for each sample. The data are shown as the means ±SDs. a, b, and c indicate significant differences at the p ≤ 0.05 level in the Duncan test.
3. Discussion
It is well known that the TFs in the same subgroup usually have similar function [36]. In plants, R2R3-MYB TFs can be categorized into at least 25 subgroups, of which subgroup 7 plays a vital role in the synthesis of flavonol, which is characterized by the SG7 motif ([K/R][R/x][R/K]xGRT[S/x][R/G]xx[M/x]K) and the SG7-2 motif ([W/x][L/x]LS) [19]. Phylogenetic analysis indicated that PqMYBF1 was clustered within subgroup 7, with other known flavonol regulators such as AtMYB11, AtMYB12, and AtMYB111 from Arabidopsis, GtMYBP3, and GtMYBP4 from gentian and VvMYBF1 from grape (Figure 1A) [7,8,19,37]. Sequence analysis showed that PqMYF1 not only contains R2 and R3 domain, but also includes the SG7 motif and the SG7-2 motif (Figure 1B). In addition, no [DE]Lx(2)[RK]x(3)Lx(6)Lx(3)R motif for interaction with bHLH proteins was found in PqMYBF1, indicating that PqMYBF1 is functionally independent on bHLH cofactors. So far, no cofactors have been reported as being necessary for plant flavonol-specific MYBs [5]. All of these indicated that PqMYBF1 should be a potential R2R3-MYB protein that has the function of regulating flavonol biosynthesis.
Previous studies have shown that flavonol accumulation was promoted by flavonol biosynthesis MYB regulator [36,38]. In grape, the expression level of VvMYBF1 was closely related to the accumulation of flavonol in grape berries. Overexpression of MdMYB22 in apple callus significantly increased flavonol accumulation [20]. Mutations of the flavonol-specific regulators AtMYB11, AtMYB12, and AtMYB111 resulted in an abolition of flavonol accumulation in Arabidopsis [23]. Functions of MYB regulators are not only conserved in the same species, but also play a similar role in heterologous plants. For example, overexpression of AtMYB12 and AtMYB11 in transgenic tobacco or tomato modulated the flavonoid pathway genes and up-regulated flavonol content [7,39]. Overexpression of GtMYBP3 and GtMYBP4 from gentian in tobacco promoted flavonol synthesis, resulting in a pink-white petal phenotype [37]. In the present study, the expression trends of PqMYBF1, PqCHS, PqCHI, PqF3H, and PqFLS were consistent with the accumulation of flavonol in P.qiui leaves (Figure 3). Up-regulation of flavonol synthesis genes (NtCHS, NtCHI, NtF3H, and NtFLS) was detected in PqMYBF1-overexpressing tobacco and flavonol accumulation was increased in flowers and leaves of transgenic tobacco (Figure 4C and Figure 5). Based on this, we reasoned that a similar function of PqMYBF1 should exist in tree peony.
Flavonol and anthocyanin biosynthesis are two important branches of flavonoid pathway. Flavonol and anthocyanin share the early biosynthetic pathway of flavonoids. CHS uses coumaroyl-CoA and malonyl-CoA as substrates to form naringenin chalcone, which was converted naringenin by CHI. F3H converts naringenin to dihydroflavonols. Then, dihydroflavonols are converted to flavonols by FLS. At the same time, dihydroflavonols can also formed to leucoanthocyanins by DFR, and then leucoanthocyanins are converted to anthocyanins by ANS. Therefore, DFR and FLS target the same substrate (dihydroflavonol). The competition relationship between them directly affects the accumulation of anthocyanins and flavonols [5,40]. In this study, overexpression of PqMYBF1 in tobacco significantly up-regulated the expression levels of NtCHS, NtF3H, and NtFLS and slightly down-regulated the expression levels of NtDFR and NtANS, which affected the direction of metabolic flux from anthocyanin to flavonol pathway and caused the accumulation of flavonols and the reduction of anthocyanins, resulting in a pink-white petal phenotype in transgenic tobacco (Figure 4). Similar results were also reported in AtMYB12, AtMYB11, and FeMYBF1 [41,42].
AtMYB11, AtMYB12, and AtMYB11 identified in Arabidopsis exerted their regulatory function on the biosynthesis of flavonol in the root and cotyledons by transactivation of CHS, CHI, F3H, and FLS [7,19,36]. AtMYB21 in Arabidopsis stamen regulated flavonol accumulation by increasing AtFLS1 promoter activity [43]. In M. domestica, MYB22 directly binded to FLS promoter and affect flavonol biosynthesis [44,45]. In gerbera, GhMY1a significantly activated the promoter of NtCHS and NtFLS, causing an increase in flavonol accumulation in tobacco [24]. A study of F.esculentum reported trans-activation function of the flavonol modulator FeMYBF1 on FeCHS, FeFLS and FeFLS1 [41]. In M. truncatula, MtMYB134 regulate flavonol biosynthesis by interacting with the promoters of MtCHS and MtFLS [23]. In our study, PqMYBF1 positively regulated flavonol biosynthesis by activating the activity of PqCHS, PqF3H, and PqFLS promoters (Figure 7). All these results revealed that the regulation mechanism of flavonol biosynthesis is species-specific. In addition, some MYB TFs are flavonol-specific regulators, and they only control flavonol accumulation [5,7,9,19,21,23,25], while other MYB TFs not only regulate flavonol biosynthesis, but also anthocyanin or proanthocyanidin biosynthesis [24,40,46,47]. Differences of promoter sequence in structural genes in different species may be responsible for these different results [7,38].
4. Materials and Methods
4.1. Plant Materials
The samples of P. qiui were collected from the tree peony germplasm garden of Northwest A&F University at 9:00–11:00 am during the March and April March 2021 (10–18 °C in the day and 5–8 °C in the night), including the leaves, at five different leaf color stages (S1, S2, S3, S4, and S5) (Figure 3A). The collected leaves were immediately frozen with liquid nitrogen and stored at −80 °C for subsequent use. Nicotiana tabacum and Nicotiana benthamiana were cultivated in the climate chamber at an ambient temperature of 22–25 °C, a humidity of 60–70%, and a cycle of 16 h of light/8 h of darkness.
4.2. Cloning and Bioinformatics Analysis of PqMYBF1
Previous studies have shown that tree peony and V. vinifera are closer in terms of genetic relationship [30]. To identify flavonol-related transcription factor in P. qiui, we downloaded a protein sequence named VvMYBF1 (ACT88298) of grape (V. vinifera) which positively regulates flavonol biosynthesis from the National Center for Biotechnology Information (NCBI) GenBank database [8]. Then, VvMYBF1 was used to query against the transcriptome data of P. qiui leaves using the BLAST program. One query had the best hit, and was designated as PqMYBF1. The full length of PqMYBF1 was cloned from the cDNA of P.qiui leaves using specific primers (primer sequences were designed using Oligo7.0 software and listed in Table S1). The complete open reading frame (ORF) of PqMYBF1 was integrated into the pCAMBIA1300 vector, and the isolation and sequencing of the recombinant plasmid was completed at Tsingke Biotech Beijing China. MYB proteins known to be associated with flavonoid synthesis were retrieved from NCBI and constructed the phylogenetic tree of PqMYBF1 and other known flavonoid synthesis-related MYB proteins using the neighbor-joining method of MEGA 7.0 software. Motif Scan was used to predict the conserved domain. Multiple sequence alignment was performed using DNAMAN 8.0.
4.3. RNA Isolation and qRT-PCR
Total RNA were extracted from P.qiui using the RNA prep Pure Plant kit (Tiangen Biotech Co. Ltd., Beijing, China). After testing the quality and concentration of total RNA, genomic DNA were removed using a PrimeScript® RT reagent Kit with gDNA Eraser (DRR047A, Takara, Japan). Then, cDNA was synthesized from 1 μg RNA sample. After diluting cDNA to 200 ng/µg with RNA-free H2O, qRT-PCR experiments were performed using SYBR Premix Ex Taq II (DRR041A, Takara, Japan) on StepOnePlus Real-Time PCR system (Applied Biosystems, Foster City, CA, USA). To normalize the expression data, Ubiquitin was used as a reference gene. The relative expression levels of genes were calculated using 2−△△Ct comparative threshold cycle (Ct) method. The gene-specific primers used for qRT-PCR analysis were listed in Table S2. To ensure the accuracy of the data, three biological replicates were performed for each gene.
4.4. Tobacco Transformation
An 1140bp fragment containing PqMYBF1 (open reading frame) ORF is inserted into the overexpression vector pcAMBIA1300 using Xbal and Ncol restriction sites to form a recombinant plasmid pCAMBIA1300-PqMYBF1. The fragment containing a 35S-CaMV promoter, the PqMYBF1 ORF, and a hygromycin resistance gene in the recombinant plasmid were integrated into the tobacco genome using an Agrobacterium-mediated leaf disc transformation method. Transgenic tobacco plants were screened in MS medium using 20 mg/L hygromycin, and the positive transformants were further identified by PCR method. Transgenic tobaccos and control lines were grown in the same environment and photographed under the same light conditions.
4.5. Subcellular Localization
The coding sequence of PqMYBF1 without the termination codon was constructed to the 5′ end of the GFP gene using a homologous recombination kit (Novoprotein, Shanghai, China) in the pCAMBIA2300-GFP (Kana Resistance) vector (Primers were listed in Table S3). The onion epidermis was disinfected with 70% ethanol, then the inner epidermis was torn and spread flat on MS medium in a sterile environment and cultured for one week. The onion epidermis samples were immersed in the Agrobacterium carrying the 35S: GFP-PqMYBF1 vector and the mCherry protein directed to the nucleus localization for at least 12 h dark at 26 °C. The GFP signal and mCherry fluorescent signal were observed using a laser scanning confocal microscope.
4.6. Transcriptional Activity Test
The target vector pGBKT7-PqMYBF1 was constructed using a seamless cloning and assembly kit (Novoprotein, Shanghai, China), and the primer sequences are shown in Table S4. The PGBKT7-PqMYBF1 vector and a negative control of pGBKT7 vector only containing the BD domain of GAL protein were introduced into yeast Y2H competent cells, respectively. Monoclonal yeast plaques of pGBKT7-PqMYBF1 and pGBKT7 plasmids were picked and dissolved in 200 µL 0.09% Nacl solution and adjusted to be OD600 = 1. The absorbed dilutions were incubated on SD/-Trp, SD/-Trp + 40 µg/mL X-α-Gal (Coolaber, Beijing, China), SD/-Trp + 40µg/mL X-α-Gal + 200 ng/mL AbA (Clontech, Mountain View, CA, USA) medium at 29 °C for about 72 h to observe the formation of plaque and coloration.
4.7. GUS Reporter Assay
The promoters of PqCHS, PqF3H, and PqFLS were inserted into pBI121-GUS to activate the GUS reporter gene. Primers are listed in Table S5. The recombinant vectors were introduced into Agrobacterium tumefaciens strain GV3101. Agrobacterium cultures carrying the empty vector pBI121 served as a positive control. Bacterial liquids carried positive controls. Agrobacterium cultures carrying the recombinant vector were injected into tobacco leaves, respectively. GUS staining analysis was as previously described [48].
4.8. Dual-Luciferase Reporter Assay
The promoter sequences of the PqCHS, PqF3H, and PqFLS genes were cloned and inserted into the pGreenII-0800-LUC vector to construct the reporter vector, and the effector vector was constructed by inserting the PqMYBF1 ORF into PGreen-62-SK vector. Primers used above are listed in Table S6. The effector vector and reporter vector were, respectively, combined and introduced into A. tumefaciens strain, then the bacterial liquids were injected into tobacco leaves (N. benthamiana) for transient expression analysis. The control group consisted of the pGREENII-62-SK vector combined with a reporter vector containing promoter sequence. The fluorescence expression status of firefly luciferase and renilla luciferase in tobacco leaves was detected under a promega luminometer (Promega, Madison, WI, USA), and the LUC/REN fluorescence ratio was calculated. Three biological replicates were performed for each sample group.
4.9. Measurement of Anthocyanin and Flavonol Content
Fully open flowers were collected for anthocyanin extraction from wild type and transgenic tobacco lines. The petals were cooled immediately using liquid nitrogen to prevent browning. Then, 25 mg samples were ground into powder in liquid nitrogen, to which were then added 250 µL of methanol and 1% HCL (v/v), and were then extracted at 4 °C for 24 h. Mixtures were centrifuged at 10,000 rpm for 10 min at 4 °C, and the supernatants were collected. Absorbance of the supernatant was measured at 530 and 657 nm by a spectrophotometer. The relative anthocyanin levels were calculated by a formula of (A530 nm–0.33 A657 nm)/mg. Flavonol was extracted in the same way using a methanol solution, and the supernatants were filtered through 0.22-μm filter membrane. The relative flavonol level was calculated by the formula (A350–A650)/mg. HPLC analysis were carried out to measure flavonol content in tobacco according to the method described previously [49]. Three samples were collected from each plant, and each sample was measured in triplicate.
5. Conclusions
PqMYBF1 is an R2R3-MYB transcription factor that has the SG7 and SG7-2 motifs, which are characteristics of flavonol biosynthesis activators. PqMYBF1 could promote flavonol biosynthesis and accumulation by activating the promoter activity of PqCHS, PqF3H, and PqFLS, and promoting their expression. Our results shows that PqMYBF1 is a functional flavonol-specific TF. These findings will deepen our understanding about tree peony leaf color transformation in spring.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12071427/s1, Figure S1: The sequence information of PqMYBF1; Figure S2: Original Images of gel for transgenic plant identification; Table S1: The primers used for PqMYBF1 gene cloning; Table S2: The gene specific primers used for qRT-PCR analysis; Table S3: The primers used for constructing vectors of subcellular localization assay; Table S4: The primers used for constructing vectors of transcriptional activity test; Table S5: The primers used for constructing vectors of GUS staining analysis; Table S6: The primers used for constructing vectors of dual-luciferase reporter assay.
Author Contributions
Y.Z. (Yue Zhang), J.D. and J.L. initiated and designed the research; Y.Z. (Yue Zhang), J.L. and Y.Z. (Yanlong Zhang) wrote, reviewed, and edited the manuscript; Y.Z. (Yue Zhang), J.D., Q.W., M.Z. and H.Z. conducted the experiments; Y.Z. (Yue Zhang), J.D. and Z.B. analyzed the data; J.L. was the author and responsible person of this study and directed the experimental design, data analysis, writing and revision of the paper; all authors approved the final article. All authors have read and agreed to the published version of the manuscript.
Funding
This research work has been supported by the National Natural Science Foundation of China (Grant No. 31971709, 32271950) and Shaanxi Science and Technology Plan Project (Grant No. 2022NY-146).
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no potential conflict of interest.
References
- Tang, Y.; Fang, Z.; Liu, M.; Zhao, D.; Tao, J. Color characteristics, pigment accumulation and biosynthetic analyses of leaf color variation in herbaceous peony (Paeonia lactiflora Pall.). 3 Biotech 2020, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.-X.; Wei, J.; Chi, R.-F.; Qiao, Y.-H.; Zhou, J.; Wang, Y.-L.; Wang, H.; Li, H.-H. MrMYB44-Like Negatively Regulates Anthocyanin Biosynthesis and Causes Spring Leaf Color of Malus ‘Radiant’ to Fade from Red to Green. Front. Plant Sci. 2022, 13, 42. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Ning, G.; Wang, Z.; Shen, Y.; Jin, H.; Li, P.; Huang, S.; Zhao, J.; Bao, M. Disequilibrium of Flavonol Synthase and Dihydroflavonol-4-Reductase Expression Associated Tightly to White vs. Red Color Flower Formation in Plants. Front. Plant Sci. 2015, 6, 1257. [Google Scholar] [CrossRef]
- Cao, Y.; Xie, L.; Ma, Y.; Ren, C.; Xing, M.; Fu, Z.; Wu, X.; Yin, X.; Xu, C.; Li, X. PpMYB15 and PpMYBF1 Transcription Factors Are Involved in Regulating Flavonol Biosynthesis in Peach Fruit. J. Agric. Food Chem. 2019, 67, 644–652. [Google Scholar] [CrossRef]
- Falcone Ferreyra, M.L.; Casas, M.I.; Questa, J.I.; Herrera, A.L.; Deblasio, S.; Wang, J.; Jackson, D.; Grotewold, E.; Casati, P. Evolution and expression of tandem duplicated maize flavonol synthase genes. Front. Plant Sci. 2012, 3, 101. [Google Scholar] [CrossRef]
- Mehrtens, F.; Kranz, H.; Bednarek, P.; Weisshaar, B. The Arabidopsis transcription factor MYB12 is a flavonol-specific regulator of phenylpropanoid biosynthesis. Plant Physiol. 2005, 138, 1083–1096. [Google Scholar] [CrossRef]
- Czemmel, S.; Stracke, R.; Weisshaar, B.; Cordon, N.; Harris, N.N.; Walker, A.R.; Robinson, S.P.; Bogs, J. The Grapevine R2R3-MYB Transcription Factor VvMYBF1 Regulates Flavonol Synthesis in Developing Grape Berries. Plant Physiol. 2009, 151, 1513–1530. [Google Scholar] [CrossRef]
- Li, H.; Li, Y.; Yu, J.; Wu, T.; Zhang, J.; Tian, J.; Yao, Y. MdMYB8 is associated with flavonol biosynthesis via the activation of the MdFLS promoter in the fruits of Malus crabapple. Hortic. Res. 2020, 7, 19. [Google Scholar] [CrossRef]
- Park, N.I.; Li, X.; Thwe, A.A.; Lee, S.Y.; Kim, S.G.; Wu, Q.; Park, S.U. Enhancement of rutin in Fagopyrum esculentum hairy root cultures by the Arabidopsis transcription factor AtMYB12. Biotechnol. Lett. 2012, 34, 577–583. [Google Scholar] [CrossRef]
- Yuan, Y.W.; Rebocho, A.B.; Sagawa, J.M.; Stanley, L.E.; Bradshaw, H.D., Jr. Competition between anthocyanin and flavonol biosynthesis produces spatial pattern variation of floral pigments between Mimulus species. Proc. Natl. Acad. Sci. USA 2016, 113, 2448–2453. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Higashi, Y.; Nakabayashi, R. The Origin and Evolution of Plant Flavonoid Metabolism. Front. Plant Sci. 2019, 10, 943. [Google Scholar] [CrossRef]
- Zhao, D.; Tao, J. Recent advances on the development and regulation of flower color in ornamental plants. Front. Plant Sci. 2015, 6, 261. [Google Scholar] [CrossRef]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef]
- Xu, W.; Dubos, C.; Lepiniec, L. Transcriptional control of flavonoid biosynthesis by MYB-bHLH-WDR complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef]
- Naing, A.H.; Kim, C.K. Roles of R2R3-MYB transcription factors in transcriptional regulation of anthocyanin biosynthesis in horticultural plants. Plant Mol. Biol. 2018, 98, 1–18. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Hichri, I.; Barrieu, F.; Bogs, J.; Kappel, C.; Delrot, S.; Lauvergeat, V. Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. J. Exp. Bot. 2011, 62, 2465–2483. [Google Scholar] [CrossRef]
- Stracke, R.; Ishihara, H.; Huep, G.; Barsch, A.; Mehrtens, F.; Niehaus, K.; Weisshaar, B. Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling. Plant J. 2007, 50, 660–677. [Google Scholar] [CrossRef]
- Wang, N.; Xu, H.; Jiang, S.; Zhang, Z.; Lu, N.; Qiu, H.; Qu, C.; Wang, Y.; Wu, S.; Chen, X. MYB12 and MYB22 play essential roles in proanthocyanidin and flavonol synthesis in red-fleshed apple (Malus sieversii f. niedzwetzkyana). Plant J. 2017, 90, 276–292. [Google Scholar] [CrossRef]
- Yao, P.; Huang, Y.; Dong, Q.; Wan, M.; Wang, A.; Chen, Y.; Li, C.; Wu, Q.; Chen, H.; Zhao, H. FtMYB6, a Light-Induced SG7 R2R3-MYB Transcription Factor, Promotes Flavonol Biosynthesis in Tartary Buckwheat (Fagopyrum tataricum). J. Agric. Food Chem. 2020, 68, 13685–13696. [Google Scholar] [CrossRef]
- Shan, X.; Li, Y.; Yang, S.; Yang, Z.; Qiu, M.; Gao, R.; Han, T.; Meng, X.; Xu, Z.; Wang, L.; et al. The spatio-temporal biosynthesis of floral flavonols is controlled by differential phylogenetic MYB regulators in Freesia hybrida. New Phytol. 2020, 228, 1864–1879. [Google Scholar] [CrossRef] [PubMed]
- Naik, J.; Rajput, R.; Pucker, B.; Stracke, R.; Pandey, A. The R2R3-MYB transcription factor MtMYB134 orchestrates flavonol biosynthesis in Medicago truncatula. Plant Mol. Biol. 2021, 106, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Tang, Y.; Pang, B.; Li, X.; Yang, Y.; Deng, J.; Feng, C.; Li, L.; Ren, G.; Wang, Y.; et al. The R2R3-MYB transcription factor GhMYB1a regulates flavonol and anthocyanin accumulation in Gerbera hybrida. Hortic. Res. 2020, 7, 78. [Google Scholar] [CrossRef] [PubMed]
- Zhai, R.; Zhao, Y.; Wu, M.; Yang, J.; Li, X.; Liu, H.; Wu, T.; Liang, F.; Yang, C.; Wang, Z.; et al. The MYB transcription factor PbMYB12b positively regulates flavonol biosynthesis in pear fruit. BMC Plant Biol. 2019, 19, 85. [Google Scholar] [CrossRef]
- Aharoni, A.; De Vos, C.H.R.; Wein, M.; Sun, Z.K.; Greco, R.; Kroon, A.; Mol, J.N.M.; O’Connell, A.P. The strawberry FaMYB1 transcription factor suppresses anthocyanin and flavonol accumulation in transgenic tobacco. Plant J. 2001, 28, 319–332. [Google Scholar] [CrossRef]
- Li, C.; Wu, J.; Hu, K.-D.; Wei, S.-W.; Sun, H.-Y.; Hu, L.-Y.; Han, Z.; Yao, G.-F.; Zhang, H. PyWRKY26 and PybHLH3 cotargeted the PyMYB114 promoter to regulate anthocyanin biosynthesis and transport in red-skinned pears. Hortic. Res. 2020, 7, 37. [Google Scholar] [CrossRef]
- Duan, J.; Luo, J.; Li, X.; Zhang, Q.; Zhang, Y. Analysis of Pigment Changes and Related Gene Expression during the Red Faded of Tree Peony Leaves in Spring. Acta Bot. Bras. 2018, 38, 1886–1894. (In Chinese) [Google Scholar] [CrossRef]
- Duan, J. Screening, Cloning and Functional Verification of MYB Related to Leaf Anthocyanin Siynthesis in Paeonia qiui; Northwest A&F University: Yangling, China, 2019. [Google Scholar]
- Luo, J.; Duan, J.; Huo, D.; Shi, Q.; Niu, L.; Zhang, Y. Transcriptomic Analysis Reveals Transcription Factors Related to Leaf Anthocyanin Biosynthesis in Paeonia qiui. Molecules 2017, 22, 2186. [Google Scholar] [CrossRef]
- Liu, X.; Duan, J.; Huo, D.; Li, Q.; Wang, Q.; Zhang, Y.; Niu, L.; Luo, J. The Paeonia qiui R2R3-MYB Transcription Factor PqMYB113 Positively Regulates Anthocyanin Accumulation in Arabidopsis thaliana and Tobacco. Front. Plant Sci. 2022, 12, 128. [Google Scholar] [CrossRef]
- Huo, D.; Liu, X.; Zhang, Y.; Duan, J.; Zhang, Y.; Luo, J. A Novel R2R3-MYB Transcription Factor PqMYB4 Inhibited Anthocyanin Biosynthesis in Paeonia qiui. Int. J. Mol. Sci. 2020, 21, 5878. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, Z.; Yu, X.; Zhao, L.; Zhao, M.; Han, X.; Qi, S. Identification of Two Novel R2R3-MYB Transcription factors, PsMYB114L and PsMYB12L, Related to Anthocyanin Biosynthesis in Paeonia suffruticosa. Int. J. Mol. Sci. 2019, 20, 1055. [Google Scholar] [CrossRef]
- Wang, H.; Liu, J.-X.; Feng, K.; Li, T.; Duan, A.-Q.; Liu, Y.-H.; Liu, H.; Xiong, A.-S. AgMYB12, a novel R2R3-MYB transcription factor, regulates apigenin biosynthesis by interacting with the AgFNS gene in celery. Plant Cell Rep. 2022, 41, 139–151. [Google Scholar] [CrossRef]
- Zimmermann, I.M.; Heim, M.A.; Weisshaar, B.; Uhrig, J.F. Comprehensive identification of Arabidopsis thaliana MYB transcription factors interacting with R/B-like BHLH proteins. Plant J. 2004, 40, 22–34. [Google Scholar] [CrossRef]
- Pandey, A.; Misra, P.; Trivedi, P.K. Constitutive expression of Arabidopsis MYB transcription factor, AtMYB11, in tobacco modulates flavonoid biosynthesis in favor of flavonol accumulation. Plant Cell Rep. 2015, 34, 1515–1528. [Google Scholar] [CrossRef]
- Nakatsuka, T.; Saito, M.; Yamada, E.; Fujita, K.; Kakizaki, Y.; Nishihara, M. Isolation and characterization of GtMYBP3 and GtMYBP4, orthologues of R2R3-MYB transcription factors that regulate early flavonoid biosynthesis, in gentian flowers. J. Exp. Bot. 2012, 63, 6505–6517. [Google Scholar] [CrossRef]
- Luo, J.; Butelli, E.; Hill, L.; Parr, A.; Niggeweg, R.; Bailey, P.; Weisshaar, B.; Martin, C. AtMYB12 regulates caffeoyl quinic acid and flavonol synthesis in tomato: Expression in fruit results in very high levels of both types of polyphenol. Plant J. 2008, 56, 316–326. [Google Scholar] [CrossRef]
- Pandey, A.; Misra, P.; Choudhary, D.; Yadav, R.; Goel, R.; Bhambhani, S.; Sanyal, I.; Trivedi, R.; Trivedi, P.K. AtMYB12 expression in tomato leads to large scale differential modulation in transcriptome and flavonoid content in leaf and fruit tissues. Sci. Rep. 2015, 5, 12412. [Google Scholar] [CrossRef]
- Li, J.; Luan, Q.; Han, J.; Zhang, C.; Liu, M.; Ren, Z. CsMYB60 directly and indirectly activates structural genes to promote the biosynthesis of flavonols and proanthocyanidins in cucumber. Hortic. Res. 2020, 7, 103. [Google Scholar] [CrossRef]
- Matsui, K.; Oshima, Y.; Mitsuda, N.; Sakamoto, S.; Nishiba, Y.; Walker, A.R.; Ohme-Takagi, M.; Robinson, S.P.; Yasui, Y.; Mori, M.; et al. Buckwheat R2R3 MYB transcription factor FeMYBF1 regulates flavonol biosynthesis. Plant Sci. 2018, 274, 466–475. [Google Scholar] [CrossRef]
- Li, Y.; Chen, M.; Wang, S.; Ning, J.; Ding, X.; Chu, Z. AtMYB11 regulates caffeoylquinic acid and flavonol synthesis in tomato and tobacco. Plant Cell Tissue Organ Cult. 2015, 122, 309–319. [Google Scholar] [CrossRef]
- Zhang, X.; He, Y.; Li, L.; Liu, H.; Hong, G. Involvement of the R2R3-MYB transcription factor MYB21 and its homologs in regulating flavonol accumulation in Arabidopsis stamen. J. Exp. Bot 2021, 72, 4319–4332. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tao, H.; Yang, H.; Zhang, L.; Feng, G.; An, Y.; Wang, L. MdSCL8 as a Negative Regulator Participates in ALA-Induced FLS1 to Promote Flavonol Accumulation in Apples. Int. J. Mol. Sci. 2022, 23, 2033. [Google Scholar] [CrossRef] [PubMed]
- Henry-Kirk, R.A.; Plunkett, B.; Hall, M.; McGhie, T.; Allan, A.C.; Wargent, J.J.; Espley, R.V. Solar UV light regulates flavonoid metabolism in apple (Malus × domestica). Plant Cell Environ. 2018, 41, 675–688. [Google Scholar] [CrossRef]
- Zhou, L.-J.; Geng, Z.; Wang, Y.; Wang, Y.; Liu, S.; Chen, C.; Song, A.; Jiang, J.; Chen, S.; Chen, F. A novel transcription factor CmMYB012 inhibits flavone and anthocyanin biosynthesis in response to high temperatures in chrysanthemum. Hortic. Res. 2021, 8, 248. [Google Scholar] [CrossRef]
- Anwar, M.; Yu, W.; Yao, H.; Zhou, P.; Allan, A.C.; Zeng, L. NtMYB3, an R2R3-MYB from Narcissus, Regulates Flavonoid Biosynthesis. Int. J. Mol. Sci. 2019, 20, 5456. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, W.; Zhang, C.-G.; Zhao, J.-F.; Chen, Y.-L. GUS Staining of Guard Cells to Identify Localised Guard Cell Gene Expression. Bio-Protocol 2017, 7, e2446. [Google Scholar] [CrossRef]
- Chen, K.; Liu, H.; Lou, Q.; Liu, Y. Ectopic Expression of the Grape Hyacinth (Muscari armeniacum) R2R3-MYB Transcription Factor Gene, MaAN2, Induces Anthocyanin Accumulation in Tobacco. Front. Plant Sci. 2017, 8, 965. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).