

Impact of Sodium Nitroprusside on the Photosynthetic Performance of Maize and Sorghum

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Pigment Composition
2.2. Contents of Malondialdehyde and Hydrogen Peroxide
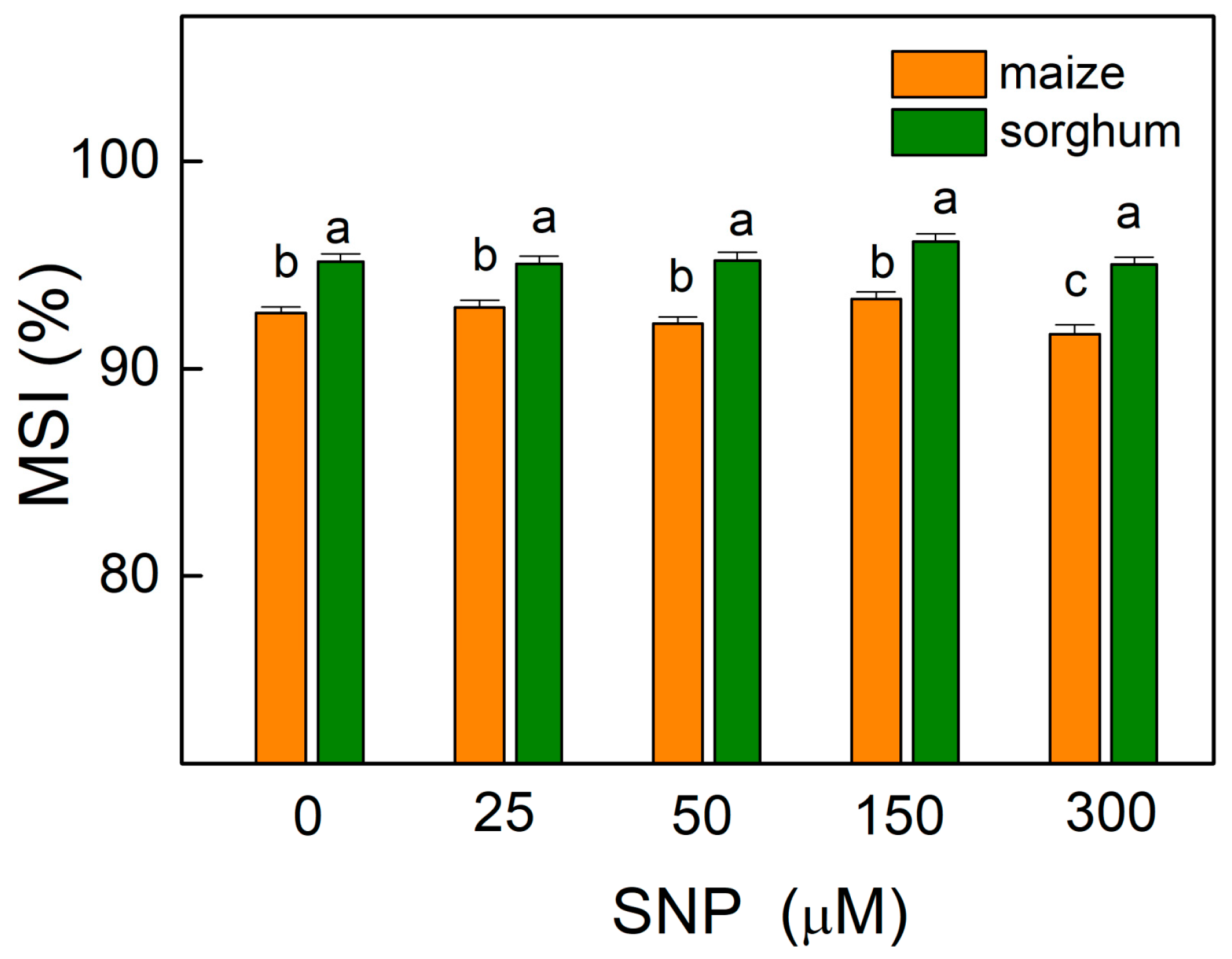
2.3. Membrane Stability Index
2.4. PAM Chlorophyll Fluorescence
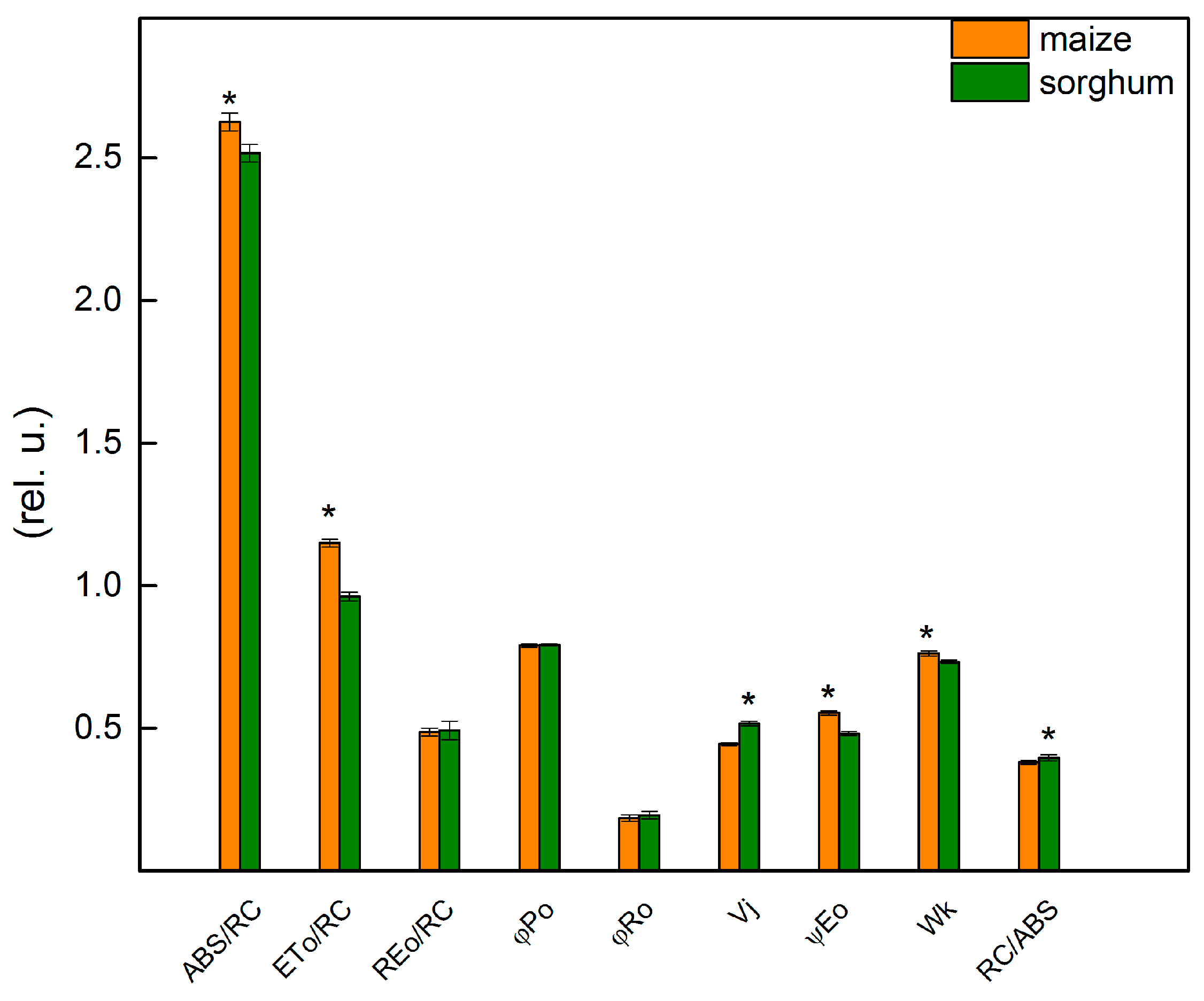
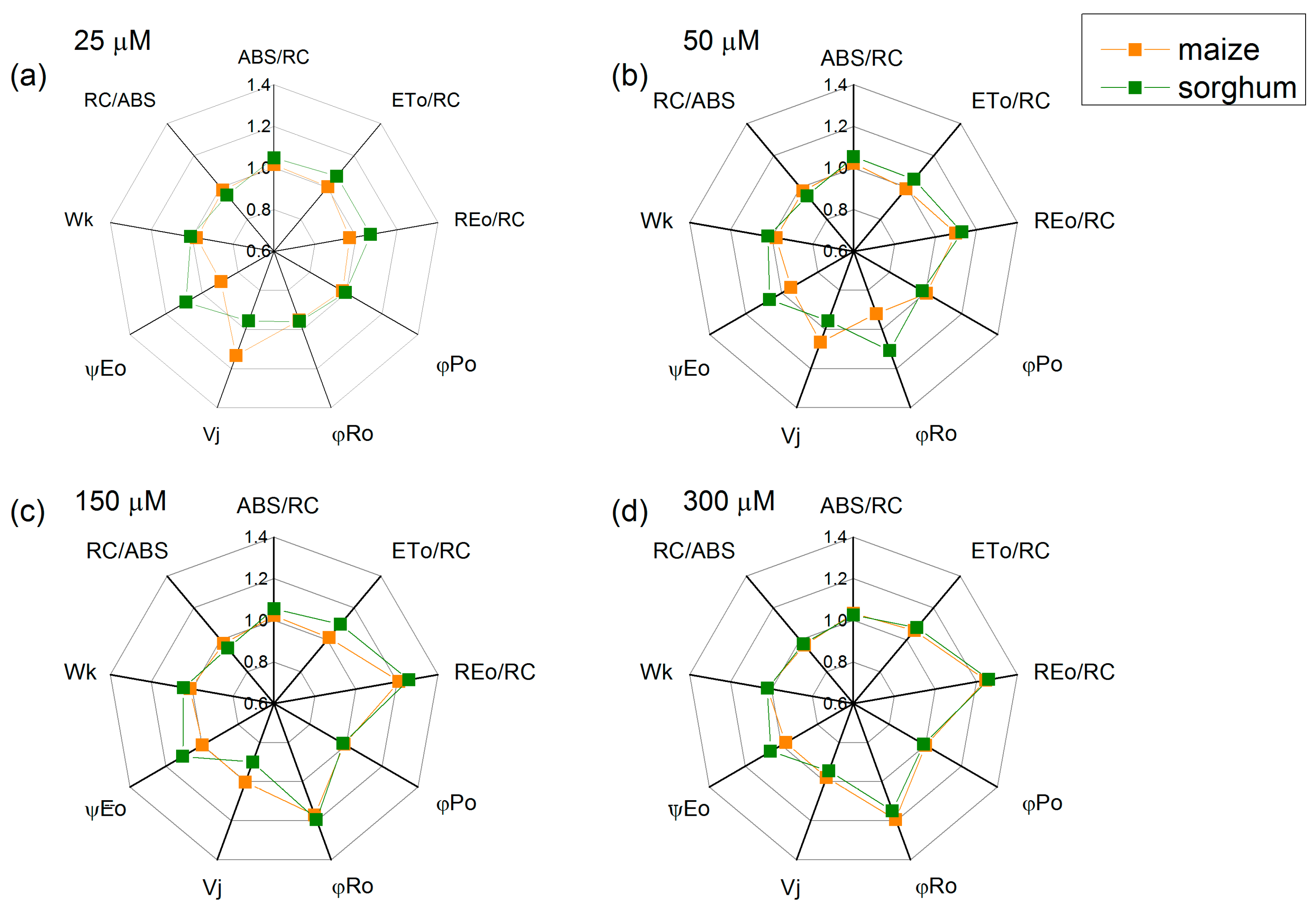
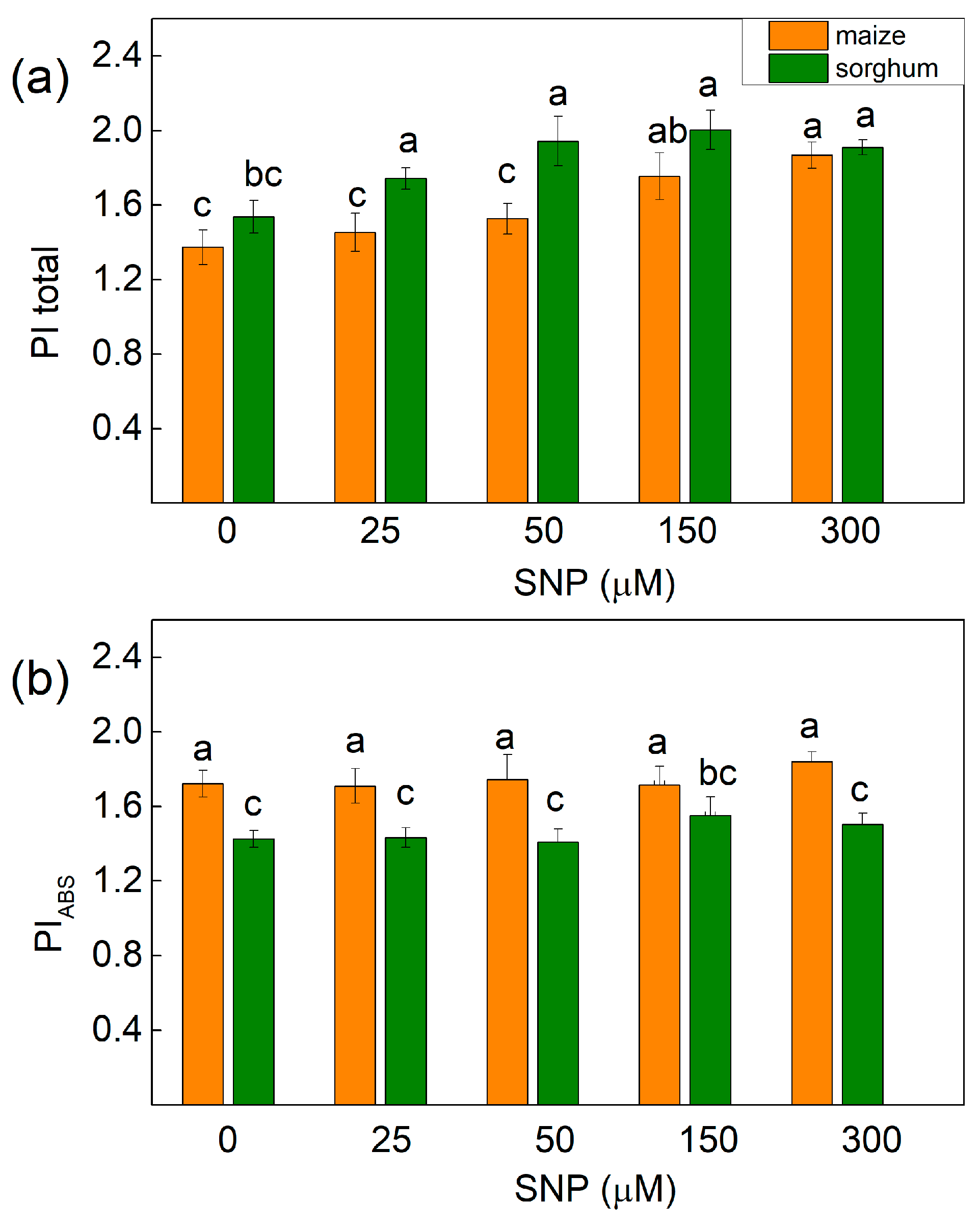
2.5. Chlorophyll Fluorescence Induction
2.6. Oxidation–Reduction Kinetics of P700
3. Discussion
4. Materials and Methods
4.1. Plant Growth Conditions and SNP Treatment
4.2. Photosynthetic Pigments
4.3. Malondialdehyde and Hydrogen Peroxide Contents
4.4. Membrane Stability Index
4.5. Chlorophyll Fluorescence Measurements
4.6. Oxidation-Reduction Kinetics of P700
4.7. Statistics
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Misra, A.N.; Singh, R.; Misra, M.; Vladkova, R.; Dobrikova, A.G.; Apostolova, E.L. Nitric Oxide Mediated Effects on Chloroplasts. In Photosynthesis: Structures, Mechanisms, and Applications; Springer International Publishing: Cham, Switzerland, 2017; pp. 305–320. ISBN 9783319488738. [Google Scholar]
- Gupta, K.J.; Kaladhar, V.C.; Fitzpatrick, T.B.; Fernie, A.R.; Møller, I.M.; Loake, G.J. Nitric oxide regulation of plant metabolism. Mol. Plant 2022, 15, 228–242. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Ali, S.; Al Azzawi, T.N.I.; Yun, B.-W. Nitric oxide acts as a key signaling molecule in plant development under stressful conditions. Int. J. Mol. Sci. 2023, 24, 4782. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.C.; Cataneo, A.C. Nitric oxide in plants: A brief discussion on this multifunctional molecule. Sci. Agric. 2010, 67, 236–243. [Google Scholar] [CrossRef]
- Gao, Z.; Zhang, J.; Zhang, J.; Zhang, W.; Zheng, L.; Borjigin, T.; Wang, Y. Nitric oxide alleviates salt-induced stress damage by regulating the ascorbate–glutathione cycle and Na+/K+ homeostasis in Nitraria tangutorum Bobr. Plant Physiol. Biochem. 2022, 173, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Manai, J.; Kalai, T.; Gouia, H.; Corpas, F.J. Exogenous nitric oxide (NO) ameliorates salinity-induced oxidative stress in tomato (Solanum lycopersicum) plants. J. Soil Sci. Plant Nutr. 2014, 14, 433–446. [Google Scholar] [CrossRef]
- Khan, E.A.; Misra, M.; Sharma, P.; Misra, A.N. Effects of exogenous nitric oxide on protein, proline, MDA contents and antioxidative system in Pea seedling (Pisum sativum L.). Res. J. Pharm. Technol. 2017, 10, 3137. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.H.M.B.; Mahmud, J.A.; Baluska, F.; Fujita, M. Nitric oxide-induced salt stress tolerance in plants: ROS metabolism, signaling, and molecular interactions. Plant Biotechnol. Rep. 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Santisree, P.; Sanivarapu, H.; Gundavarapu, S.; Sharma, K.K.; Bhatnagar-Mathur, P. Nitric oxide as a signal in inducing secondary metabolites during plant stress. In Reference Series in Phytochemistry; Springer: Cham, Switzerland, 2020; pp. 593–621. [Google Scholar] [CrossRef]
- Wodala, B.; Deák, Z.; Vass, I.; Erdei, L.; Altorjay, I.; Horváth, F. In vivo target sites of Nitric oxide in photosynthetic electron transport as studied by chlorophyll fluorescence in pea leaves. Plant Physiol. 2008, 146, 1920–1927. [Google Scholar] [CrossRef]
- Misra, A.N.; Vladkova, R.; Singh, R.; Misra, M.; Dobrikova, A.G.; Apostolova, E.L. Action and target sites of nitric oxide in chloroplasts. Nitric Oxide-Biol. Chem. 2014, 39, 35–45. [Google Scholar] [CrossRef]
- Stefanov, M.A.; Rashkov, G.D.; Yotsova, E.K.; Borisova, P.B.; Dobrikova, A.G.; Apostolova, E.L. Protective effects of sodium nitroprusside on photosynthetic performance of Sorghum bicolor L. under salt stress. Plants 2023, 12, 832. [Google Scholar] [CrossRef]
- Wang, L.; Lin, R.; Xu, J.; Song, J.; Shao, S.; Yu, J.; Zhou, Y. High nitric oxide concentration inhibits photosynthetic pigment biosynthesis by promoting the degradation of transcription factor HY5 in tomato. Int. J. Mol. Sci. 2022, 23, 6027. [Google Scholar] [CrossRef] [PubMed]
- Procházková, D.; Haisel, D.; Wilhelmová, N.; Pavlíková, D.; Száková, J. Effects of exogenous nitric oxide on photosynthesis. Photosynthetica 2013, 51, 483–489. [Google Scholar] [CrossRef]
- Fan, J.; Chen, K.; Amombo, E.; Hu, Z.; Chen, L.; Fu, J. Physiological and molecular mechanism of nitric oxide (NO) involved in bermudagrass response to cold stress. PLoS ONE 2015, 10, e0132991. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, Y.; Zhao, L.; Shi, S.; Zhang, L. Involvement of nitric oxide in light-mediated greening of barley seedlings. J. Plant Physiol. 2006, 163, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Galatro, A.; Puntarulo, S.; Guiamet, J.J.; Simontacchi, M. Chloroplast functionality has a positive effect on nitric oxide level in soybean cotyledons. Plant Physiol. Biochem. 2013, 66, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Wodala, B.; Deák, Z.; Vass, I.; Erdei, L.; Horváth, F. Nitric oxide modifies photosynthetic electron transport in pea leaves. Acta Biol. Szeged. 2005, 49, 7–8. [Google Scholar]
- Vladkova, R.; Dobrikova, A.G.; Singh, R.; Misra, A.N.; Apostolova, E. Photoelectron transport ability of chloroplast thylakoid membranes treated with NO donor SNP: Changes in flash oxygen evolution and chlorophyll fluorescence. Nitric Oxide 2011, 24, 84–90. [Google Scholar] [CrossRef]
- Ioannidis, N.; Schansker, G.; Barynin, V.V.; Petrouleas, V. Interaction of nitric oxide with the oxygen evolving complex of photosystem II and manganese catalase: A comparative study. J. Biol. Inorg. Chem. 2000, 5, 354–363. [Google Scholar] [CrossRef]
- Shams, M.; Ekinci, M.; Ors, S.; Turan, M.; Agar, G.; Kul, R.; Yildirim, E. Nitric oxide mitigates salt stress effects of pepper seedlings by altering nutrient uptake, enzyme activity and osmolyte accumulation. Physiol. Mol. Biol. Plants 2019, 25, 1149–1161. [Google Scholar] [CrossRef]
- Kaya, C.; Akram, N.A.; Ashraf, M. Influence of exogenously applied nitric oxide on strawberry (Fragaria × ananassa) plants grown under iron deficiency and/or saline stress. Physiol. Plant. 2019, 165, 247–263. [Google Scholar] [CrossRef]
- Antoniou, C.; Filippou, P.; Mylona, P.; Fasoula, D.; Ioannides, I.; Polidoros, A.; Fotopoulos, V. Developmental stage-and concentration- specific sodium nitroprusside application results in nitrate reductase regulation and the modification of nitrate metabolism in leaves of Medicago truncatula plants. Plant Signal. Behav. 2013, 8, e25479. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Guo, F.-Q. Nitric oxide deficiency accelerates chlorophyll breakdown and stability loss of thylakoid membranes during dark-induced leaf senescence in Arabidopsis. PLoS ONE 2013, 8, e56345. [Google Scholar] [CrossRef] [PubMed]
- Gautam, H.; Sehar, Z.; Rehman, M.T.; Hussain, A.; AlAjmi, M.F.; Khan, N.A. Nitric oxide enhances photosynthetic nitrogen and sulfur-use efficiency and activity of ascorbate-glutathione cycle to reduce high temperature stress-induced oxidative stress in rice (Oryza sativa L.) plants. Biomolecules 2021, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Elkelish, A.; Ibrahim, M.F.M.; Ashour, H.; Bondok, A.; Mukherjee, S.; Aftab, T.; Hikal, M.; El-Yazied, A.A.; Azab, E.; Gobouri, A.A.; et al. Exogenous application of nitric oxide mitigates water stress and reduces natural viral disease incidence of tomato plants subjected to deficit irrigation. Agronomy 2021, 11, 87. [Google Scholar] [CrossRef]
- Adamu, T.; Mun, B.-G.; Lee, S.-U.; Hussain, A.; Yun, B.-W. Exogenously applied nitric oxide enhances salt tolerance in rice (Oryza sativa L.) at seedling stage. Agronomy 2018, 8, 276. [Google Scholar] [CrossRef]
- Safian, N.; Naderi, M.R.; Torabi, M.; Soleymani, A.; Salemi, H.R. Corn (Zea mays L.) and sorghum (Sorghum bicolor (L.) Moench) yield and nutritional quality affected by drought stress. Biocatal. Agric. Biotechnol. 2022, 45, 102486. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C.; Knapp, M. How to correctly determine the different chlorophyll fluorescence parameters and the chlorophyll fluorescence decrease ratio RFd of leaves with the PAM fluorometer. Photosynthetica 2005, 43, 379–393. [Google Scholar] [CrossRef]
- Bukhov, N.G.; Samson, G.; Carpentier, R. Nonphotosynthetic reduction of the intersystem electron transport chain of chloroplasts following heat stress. The pool size of stromal reductants. Photochem. Photobiol. 2001, 74, 438. [Google Scholar] [CrossRef]
- Shirao, M.; Kuroki, S.; Kaneko, K.; Kinjo, Y.; Tsuyama, M.; Förster, B.; Takahashi, S.; Badger, M.R. Gymnosperms have increased capacity for electron leakage to oxygen (Mehler and PTOX reactions) in photosynthesis compared with angiosperms. Plant Cell Physiol. 2013, 54, 1152–1163. [Google Scholar] [CrossRef]
- Deák, Z.; Sass, L.; Kiss, É.; Vass, I. Characterization of wave phenomena in the relaxation of flash-induced chlorophyll fluorescence yield in cyanobacteria. Biochim. Biophys. Acta-Bioenerg. 2014, 1837, 1522–1532. [Google Scholar] [CrossRef]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Kulkarni, K.P.; Imran, Q.M.; Mun, B.-G.; Yun, B.-W. Nitric oxide regulates plant responses to drought, salinity, and heavy metal stress. Environ. Exp. Bot. 2019, 161, 120–133. [Google Scholar] [CrossRef]
- Young, A.J. The photoprotective role of carotenoids in higher plants. Physiol. Plant. 1991, 83, 702–708. [Google Scholar] [CrossRef]
- Swapnil, P.; Meena, M.; Singh, S.K.; Dhuldhaj, U.P.; Harish; Marwal, A. Vital roles of carotenoids in plants and humans to deteriorate stress with its structure, biosynthesis, metabolic engineering and functional aspects. Curr. Plant Biol. 2021, 26, 100203. [Google Scholar] [CrossRef]
- Formaggio, E.; Cinque, G.; Bassi, R. Functional architecture of the major light-harvesting complex from higher plants. J. Mol. Biol. 2001, 314, 1157–1166. [Google Scholar] [CrossRef]
- Trebst, A.; Depka, B. Role of carotene in the rapid turnover and assembly of photosystem II in Chlamydomonas reinhardtii. FEBS Lett. 1997, 400, 359–362. [Google Scholar] [CrossRef]
- Trebst, A. Function of β-carotene and tocopherol in photosystem II. Z. Für Naturforsch. C 2003, 58, 609–620. [Google Scholar] [CrossRef]
- Stoitchkova, K.; Busheva, M.; Apostolova, E.L.; Andreeva, A. Changes in the energy distribution in mutant thylakoid membranes of pea with modified pigment content. II. Changes due to magnesium ions concentration. J. Photochem. Photobiol. B Biol. 2006, 83, 11–20. [Google Scholar] [CrossRef]
- Apostolova, E.L.; Misra, A.N. Alterations in Structural Organization Affect the Functional Ability of Photosynthetic Apparatus. In Handbook of Plant and Crop Physiology, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 103–120. [Google Scholar] [CrossRef]
- Khoshbakht, D.; Asghari, M.R.; Haghighi, M. Effects of foliar applications of nitric oxide and spermidine on chlorophyll fluorescence, photosynthesis and antioxidant enzyme activities of citrus seedlings under salinity stress. Photosynthetica 2018, 56, 1313–1325. [Google Scholar] [CrossRef]
- Huang, W.; Yang, Y.-J.; Zhang, S.-B. Specific roles of cyclic electron flow around photosystem I in photosynthetic regulation in immature and mature leaves. J. Plant Physiol. 2017, 209, 76–83. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Rastogi, A.; Živčák, M.; Brestic, M.; Daszkowska-Golec, A.; Sitko, K.; Alsharafa, K.Y.; Lotfi, R.; Stypiński, P.; Samborska, I.A.; et al. Prompt chlorophyll fluorescence as a tool for crop phenotyping: An example of barley landraces exposed to various abiotic stress factors. Photosynthetica 2018, 56, 953–961. [Google Scholar] [CrossRef]
- Stefanov, M.; Rashkov, G.; Borisova, P.; Apostolova, E. Sensitivity of the photosynthetic apparatus in maize and sorghum under different drought levels. Plants 2023, 12, 1863. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Yotsova, E.K.; Dobrikova, A.G.; Stefanov, M.A.; Kouzmanova, M.; Apostolova, E.L. Improvement of the rice photosynthetic apparatus defence under cadmium stress modulated by salicylic acid supply to roots. Theor. Exp. Plant Physiol. 2018, 30, 57–70. [Google Scholar] [CrossRef]
- Stefanov, M.A.; Rashkov, G.D.; Yotsova, E.K.; Borisova, P.B.; Dobrikova, A.G.; Apostolova, E.L. Different sensitivity levels of the photosynthetic apparatus in Zea mays L. and Sorghum bicolor L. under salt stress. Plants 2021, 10, 1469. [Google Scholar] [CrossRef] [PubMed]
- Dobrikova, A.; Apostolova, E.; Adamakis, I.-D.S.; Hanć, A.; Sperdouli, I.; Moustakas, M. Combined impact of excess zinc and cadmium on elemental uptake, leaf anatomy and pigments, antioxidant capacity, and function of photosynthetic apparatus in clary sage (Salvia sclarea L.). Plants 2022, 11, 2407. [Google Scholar] [CrossRef] [PubMed]
- Roháček, K. Chlorophyll fluorescence parameters: The definitions, photosynthetic meaning, and mutual relationships. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Guadagno, C.R.; Virzo De Santo, A.; D’Ambrosio, N. A revised energy partitioning approach to assess the yields of non-photochemical quenching components. Biochim. Biophys. Acta-Bioenerg. 2010, 1797, 525–530. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Stefanov, M.; Yotsova, E.; Gesheva, E.; Dimitrova, V.; Markovska, Y.; Doncheva, S.; Apostolova, E.L. Role of flavonoids and proline in the protection of photosynthetic apparatus in Paulownia under salt stress. S. Afr. J. Bot. 2021, 139, 246–253. [Google Scholar] [CrossRef]
- Stefanov, M.A.; Rashkov, G.D.; Apostolova, E.L. Assessment of the photosynthetic apparatus functions by chlorophyll fluorescence and P700 absorbance in C3 and C4 plants under physiological conditions and under salt stress. Int. J. Mol. Sci. 2022, 23, 3768. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence; Advances in Photosynthesis and Respiration Series; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Tsimilli-Michael, M. Special issue in honour of Prof. Reto J. Strasser—Revisiting JIP-test: An educative review on concepts, assumptions, approximations, definitions and terminology. Photosynthetica 2020, 58, 275–292. [Google Scholar] [CrossRef]
- Salim Akhter, M.; Noreen, S.; Mahmood, S.; Athar, H.u.R.; Ashraf, M.; Abdullah Alsahli, A.; Ahmad, P. Influence of salinity stress on PSII in barley (Hordeum vulgare L.) genotypes, probed by chlorophyll-a fluorescence. J. King Saud Univ.-Sci. 2021, 33, 101239. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variants | Chl (µg/g DW) | Car (µg/g DW) | Chl a/b | Car/Chl |
|---|---|---|---|---|
| Zea mays L. | ||||
| 0 µM SNP | 28.03 ± 0.52 a | 7.91 ± 0.04 c | 4.25 ± 0.01 b | 0.21 ± 0.01 a |
| 25 µM SNP | 28.66 ± 0.65 a | 8.30 ± 0.03 b | 4.26 ± 0.03 b | 0.22 ± 0.02 a |
| 50 µM SNP | 28.00 ± 0.47 a | 8.56 ± 0.03 a | 4.25 ± 0.01 b | 0.24 ± 0.03 a |
| 150 µM SNP | 27.93 ± 0.52 a | 8.16 ± 0.05 b | 4.44 ± 0.02 b | 0.22 ± 0.01 a |
| 300 µM SNP | 26.74 ± 0.25 b | 7.86 ± 0.04 c | 4.36 ± 0.01 b | 0.22 ± 0.03 a |
| Sorghum bicolor L. | ||||
| 0 µM SNP | 20.66 ± 0.12 e | 4.89 ± 0.03 g | 5.41 ± 0.02 a | 0.24 ± 0.02 a |
| 25 µM SNP | 21.48 ± 0.06 d | 4.91 ± 0.02 g | 5.44 ± 0.02 a | 0.23 ± 0.03 a |
| 50 µM SNP | 21.49 ± 0.30 d | 4.99 ± 0.03 f | 5.43 ± 0.03 a | 0.23 ± 0.02 a |
| 150 µM SNP | 23.69 ± 0.57 c | 5.40 ± 0.02 d | 5.39 ± 0.01 a | 0.22 ± 0.01 a |
| 300 µM SNP | 23.43 ± 0.67 c | 5.18 ± 0.04 e | 5.46 ± 0.01 a | 0.22 ± 0.01 a |
| Variants | k1 (s−1) | k2 (s−1) |
|---|---|---|
| Zea mays L. | ||
| 0 µM SNP | 1.53 ± 0.14 b | 0.087 ± 0.003 b |
| 25 µM SNP | 1.81 ± 0.15 a | 0.085 ± 0.015 b |
| 50 µM SNP | 1.86 ± 0.07 a | 0.087 ± 0.006 b |
| 150 µM SNP | 1.85 ± 0.09 a | 0.088 ± 0.003 b |
| 300 µM SNP | 1.45 ± 0.08 b | 0.109 ± 0.012 a |
| Sorghum bicolor L. | ||
| 0 µM SNP | 1.87 ± 0.16 a | 0.092 ± 0.004 b |
| 25 µM SNP | 1.84 ± 0.17 a | 0.080 ± 0.006 c |
| 50 µM SNP | 1.94 ± 0.13 a | 0.069 ± 0.007 c |
| 150 µM SNP | 1.85 ± 0.12 a | 0.055 ± 0.003 c |
| 300 µM SNP | 1.76 ± 0.04 a | 0.120 ± 0.001 a |
| Variants | γRC2/(1 − γRC2) | φPo/(1 − φPo) | ψEo/(1 − ψEo) | δREo/(1 − δREo) |
|---|---|---|---|---|
| Zea mays L. | ||||
| 0 μM SNP | 0.397 ± 0.004 a | 3.798 ± 0.057 a | 1.251 ± 0.032 a | 0.759 ± 0.044 c |
| 25 μM SNP | 0.375 ± 0.007 b | 3.772 ± 0.089 a | 1.132 ± 0.051 b | 0.746 ± 0.059 c |
| 50 μM SNP | 0.391 ± 0.007 a | 3.919 ± 0.089 a | 1.118 ± 0.051 b | 0.704 ± 0.059 c |
| 150 μM SNP | 0.372 ± 0.009 b | 3.657 ± 0.095 a | 1.252 ± 0.056 a | 1.032 ± 0.066 b |
| 300 μM SNP | 0.368 ± 0.007 b | 3.843 ± 0.097 a | 1.301 ± 0.058 a | 1.011 ± 0.078 b |
| Sorghum bicolor L. | ||||
| 0 μM SNP | 0.397 ± 0.004 a | 3.844 ± 0.078 a | 0.933 ± 0.031c | 1.073 ± 0.118 b |
| 25 μM SNP | 0.379 ± 0.006 c | 3.564 ± 0.071b | 1.027 ± 0.055b | 1.307 ± 0.092 a |
| 50 μM SNP | 0.388 ± 0.008 b | 3.549 ± 0.073 b | 1.019 ± 0.029b | 1.234 ± 0.061 a |
| 150 μM SNP | 0.376 ± 0.003 c | 3.562 ± 0.046 b | 1.154 ± 0.063b | 1.281 ± 0.081 a |
| 300 μM SNP | 0.388 ± 0.008 b | 3.615 ± 0.053 b | 1.046 ± 0.021b | 1.269 ± 0.074 a |
| Variants | kF (s−1) | kS (s−1) | ∆A/A |
|---|---|---|---|
| Zea mays L. | |||
| 0 μM SNP | 0.321 ± 0.058 a | 0.031 ± 0.002 a | 10.70 ± 0.26 b |
| 25 μM SNP | 0.363 ± 0.076 a | 0.030 ± 0.001 a | 10.55 ± 0.24 b |
| 50 μM SNP | 0.384 ± 0.063 a | 0.035 ± 0.004 a | 10.70 ± 0.95 b |
| 150 μM SNP | 0.341 ± 0.043 a | 0.030 ± 0.002 a | 10.91 ± 0.64 b |
| 300 μM SNP | 0.180 ± 0.005 b | 0.022 ± 0.001 b | 11.79 ± 0.88 a |
| Sorghum bicolor L. | |||
| 0 μM SNP | 0.304 ± 0.047 a | 0.039 ± 0.003 a | 11.99 ± 0.55 a |
| 25 μM SNP | 0.278 ± 0.070 a | 0.039 ± 0.004 a | 11.39 ± 0.34 a |
| 50 μM SNP | 0.282 ± 0.034 a | 0.034 ± 0.004 a | 11.18 ± 0.43 a |
| 150 μM SNP | 0.242 ± 0.047 ab | 0.030 ± 0.002 a | 10.85 ± 0.42 ab |
| 300 μM SNP | 0.223 ± 0.025 b | 0.030 ± 0.003 a | 10.06 ± 0.23 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rashkov, G.D.; Stefanov, M.A.; Yotsova, E.K.; Borisova, P.B.; Dobrikova, A.G.; Apostolova, E.L. Impact of Sodium Nitroprusside on the Photosynthetic Performance of Maize and Sorghum. Plants 2024, 13, 118. https://doi.org/10.3390/plants13010118
Rashkov GD, Stefanov MA, Yotsova EK, Borisova PB, Dobrikova AG, Apostolova EL. Impact of Sodium Nitroprusside on the Photosynthetic Performance of Maize and Sorghum. Plants. 2024; 13(1):118. https://doi.org/10.3390/plants13010118
Chicago/Turabian StyleRashkov, Georgi D., Martin A. Stefanov, Ekaterina K. Yotsova, Preslava B. Borisova, Anelia G. Dobrikova, and Emilia L. Apostolova. 2024. "Impact of Sodium Nitroprusside on the Photosynthetic Performance of Maize and Sorghum" Plants 13, no. 1: 118. https://doi.org/10.3390/plants13010118
APA StyleRashkov, G. D., Stefanov, M. A., Yotsova, E. K., Borisova, P. B., Dobrikova, A. G., & Apostolova, E. L. (2024). Impact of Sodium Nitroprusside on the Photosynthetic Performance of Maize and Sorghum. Plants, 13(1), 118. https://doi.org/10.3390/plants13010118