

Effect of Regulation of Whole-Plant Corn Silage Inoculated with Lactobacillus buchneri or Bacillus licheniformis Regarding the Dynamics of Bacterial and Fungal Communities on Aerobic Stability
Abstract
:
1. Introduction
2. Results
2.1. Chemical Composition of Fresh Material and Nutrient Composition of Whole-Plant Corn Silage
2.2. Aerobic Stability
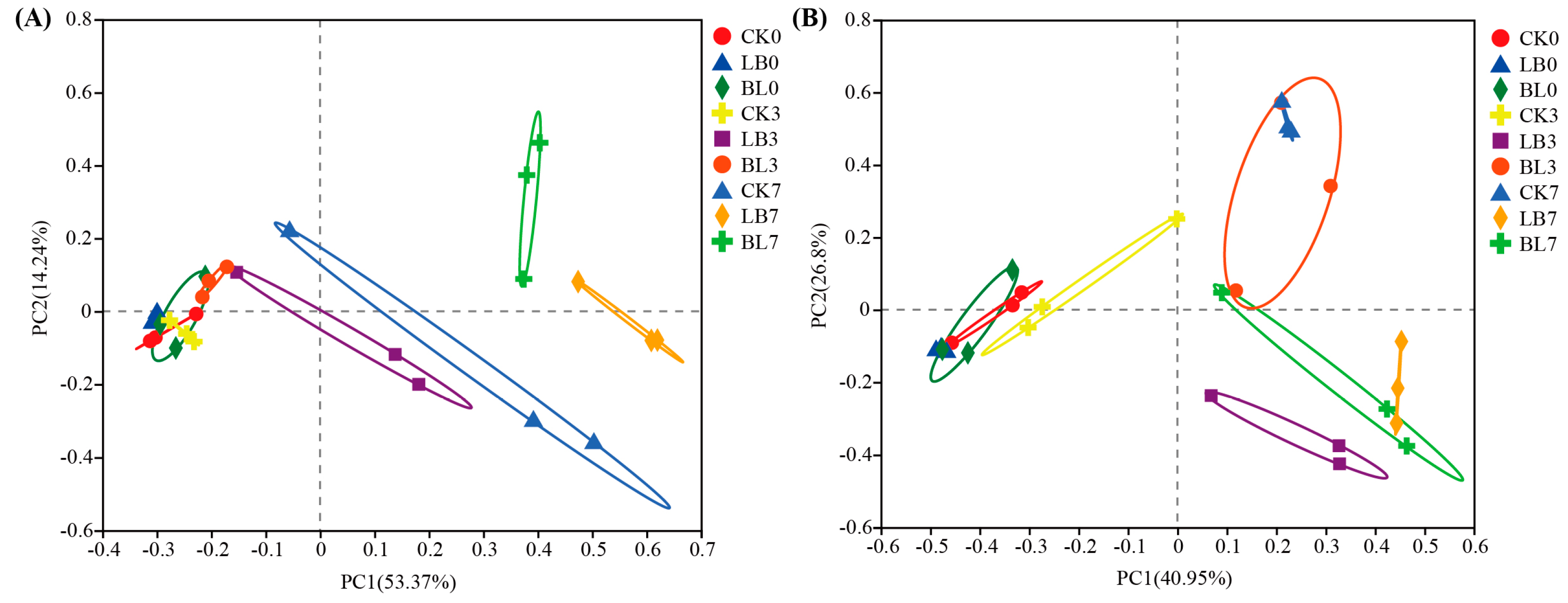
2.3. Microbial Community
3. Discussion
3.1. Characteristics of Fresh Material and Nutrient Composition of Silage
3.2. Aerobic Stability
3.3. Microbial Community
4. Materials and Methods
4.1. Materials and Silage Preparation
4.2. Aerobic Stability
4.3. Fermentation Quality and Nutrient Composition Analyses
4.4. Bacterial and Fungal Community Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hao, Y.; Huang, S.; Liu, G.; Zhang, J.; Liu, G.; Cao, Z.; Wang, Y.; Wang, W.; Li, S. Effects of different parts on the chemical composition, silage fermentation profile, in vitro and in situ digestibility of paper mulberry. Animals 2021, 11, 413. [Google Scholar] [CrossRef]
- Han, H.; Wang, C.; Huang, Z.; Zhang, Y.; Sun, L.; Xue, Y.; Guo, X. Effects of lactic acid bacteria-inoculated corn silage on bacterial communities and metabolites of digestive tract of sheep. Fermentation 2022, 8, 320. [Google Scholar] [CrossRef]
- Danner, H.; Holzer, M.; Mayrhuber, E.; Braun, R. Acetic acid increases stability of silage under aerobic conditions. Appl. Environ. Microbiol. 2003, 69, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.M.; Davies, D.R. The aerobic stability of silage: Key findings and recent developments. Grass Forage Sci. 2013, 68, 1–19. [Google Scholar] [CrossRef]
- Chen, L.; Guo, G.; Yuan, X.; Shimojo, M.; Yu, C.; Shao, T. Effect of applying molasses and propionic acid on fermentation quality and aerobic stability of total mixed ration silage prepared with whole-plant corn in Tibet. Asian-Australas. J. Anim. Sci. 2014, 27, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.P.M.d.; Santos, E.M.; Araújo, G.G.L.d.; Oliveira, J.S.d.; Zanine, A.d.M.; Pinho, R.M.A.; Cruz, G.F.d.L.; Ferreira, D.d.J.; Perazzo, A.F.; Pereira, D.M.; et al. Effect of inoculation with preactivated Lactobacillus buchneri and urea on fermentative profile, aerobic stability and nutritive value in corn silage. Agriculture 2020, 10, 335. [Google Scholar] [CrossRef]
- Weinberg, Z.G.; Khanal, P.; Yildiz, C.; Chen, Y.; Arieli, A. Ensiling fermentation products and aerobic stability of corn and sorghum silages. Grassl. Sci. 2011, 57, 46–50. [Google Scholar] [CrossRef]
- Da Silva, T.C.; Smith, M.L.; Barnard, A.M.; Kung, L. The effect of a chemical additive on the fermentation and aerobic stability of high-moisture corn. J. Dairy Sci. 2015, 98, 8904–8912. [Google Scholar] [CrossRef] [PubMed]
- B Benjamim da Silva, É.; Costa, D.M.; Santos, E.M.; Moyer, K.; Hellings, E.; Kung, L. The effects of Lactobacillus hilgardii 4785 and Lactobacillus buchneri 40788 on the microbiome, fermentation, and aerobic stability of corn silage ensiled for various times. J. Dairy Sci. 2021, 104, 10678–10698. [Google Scholar] [CrossRef]
- Serva, L.; Magrin, L.; Marchesini, G.; Andrighetto, I. Short communication: Prognostic values of a multiparametric risk score in maize silage undergoing different ensiling conditions. Agronomy 2022, 12, 774. [Google Scholar] [CrossRef]
- Xia, G.-h.; Huang, Y.; Wu, C.-r.; Zhang, M.-z.; Yin, H.-y.; Yang, F.; Chen, C.; Hao, J. Characterization of mycotoxins and microbial community in whole-plant corn ensiled in different silo types during aerobic exposure. Front. Microbiol. 2023, 14, 1136022. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Schmidt, R.J.; McDonell, E.E.; Klingerman, C.M.; Kung, L. The effect of Lactobacillus buchneri 40788 or Lactobacillus plantarum MTD-1 on the fermentation and aerobic stability of corn silages ensiled at two dry matter contents. J. Dairy Sci. 2009, 92, 3907–3914. [Google Scholar] [CrossRef] [PubMed]
- Reich, L.J.; Kung, L. Effects of combining Lactobacillus buchneri 40788 with various lactic acid bacteria on the fermentation and aerobic stability of corn silage. Anim. Feed Sci. Technol. 2010, 159, 105–109. [Google Scholar] [CrossRef]
- Ranjit, N.K.; Kung, L. The effect of Lactobacillus buchneri, Lactobacillus plantarum, or a chemical preservative on the fermentation and aerobic stability of corn silage. J. Dairy Sci. 2000, 83, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Gandra, J.R.; Oliveira, E.R.; de Sena Gandra, E.R.; Takiya, C.S.; Tonissi Buschineli de Goes, R.H.; Pires Oliveira, K.M.; Silveira, K.A.; Cariolano Araki, H.M.; Orbach, N.D.; Vasquez, D.N. Inoculation of Lactobacillus buchneri alone or with Bacillus subtilis and total losses, aerobic stability, and microbiological quality of sunflower silage. J. Appl. Anim. Res. 2017, 45, 609–614. [Google Scholar] [CrossRef]
- Bonaldi, D.S.; Carvalho, B.F.; Ávila, C.L.d.S.; Silva, C.F. Effects of Bacillus subtilis and its metabolites on corn silage quality. Appl. Microbiol. Biotechnol. 2021, 73, 46–53. [Google Scholar] [CrossRef]
- Bampidis, V.; Azimonti, G.; Bastos, M.D.L.; Christensen, H.; Dusemund, B.; Kouba, M.; Kos, D.M.; Lópezalonso, M.; López, P.S.; Marcon, F.; et al. Safety and efficacy of Bacillus licheniformis DSM 32457 as a silage additive for all animal species. EFSA J. 2019, 17, 5787. [Google Scholar]
- Afordoanyi, D.; Diabankana, R.; Krupin, E.; Bikchantaev, I.; Taylan, A.; Validov, S. Inhibitory effects of Bacillus licheniformis WJ53A and homofermentative lactic acid bacteria on clostridial growth in corn silage. Pak. J. Agric. Sci. 2023, 60, 53–63. [Google Scholar]
- Zhu, Y.; Xiong, H.; Wen, Z.; Tian, H.; Chen, Y.; Wu, L.; Guo, Y.; Sun, B. Effects of different concentrations of Lactobacillus plantarum and Bacillus licheniformis on silage quality, in vitro fermentation and microbial community of Hybrid Pennisetum. Animals 2022, 12, 1752. [Google Scholar] [CrossRef]
- Liu, B.; Yang, Z.; Huan, H.; Gu, H.; Xu, N.; Ding, C. Impact of molasses and microbial inoculants on fermentation quality, aerobic stability, and bacterial and fungal microbiomes of barley silage. Sci. Rep. 2020, 10, 5342. [Google Scholar] [CrossRef]
- Wang, T.; Teng, K.; Cao, Y.; Shi, W.; Xuan, Z.; Zhou, J.; Zhang, J.; Zhong, J. Effects of Lactobacillus hilgardii 60TS-2, with or without homofermentative Lactobacillus plantarum B90, on the aerobic stability, fermentation quality and microbial community dynamics in sugarcane top silage. Bioresour. Technol. 2020, 312, 123600. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Lu, Q.; Sun, L.; Du, S.; Liu, T.; Hou, M.; Ge, G.; Wang, Z.; Jia, Y. Effects of lactic acid bacteria additives on the quality, volatile chemicals and microbial community of leymus chinensis silage during aerobic exposure. Front. Microbiol. 2022, 13, 938153. [Google Scholar] [CrossRef]
- Yin, H.; Zhao, M.; Pan, G.; Zhang, H.; Yang, R.; Sun, J.; Yu, Z.; Bai, C.; Xue, Y. Effects of Bacillus subtilis or Lentilactobacillus buchneri on aerobic stability, and the microbial community in aerobic exposure of whole plant corn silage. Front. Microbiol. 2023, 14, 1177031. [Google Scholar] [CrossRef] [PubMed]
- Guyader, J.; Baron, V.S.; Beauchemin, K.A. Effect of harvesting corn after frost in Alberta (Canada) on whole-plant yield, nutritive value, and kernel properties. Agronomy 2021, 11, 459. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, G.; Ke, W.; Pan, Y.; Hou, M.; Chang, C.; Sa, D.; Lv, M.; Liu, Y.; Lu, Q. Effect of endogenous sodium and potassium ions in plants on the quality of alfalfa silage and bacterial community stability during fermentation. Front. Plant Sci. 2023, 14, 1295114. [Google Scholar] [CrossRef] [PubMed]
- Kung, L.; Taylor, C.C.; Lynch, M.P.; Neylon, J.M. The effect of treating alfalfa with Lactobacillus buchneri 40788 on silage fermentation, aerobic stability, and nutritive value for lactating dairy cows. J. Dairy Sci. 2003, 86, 336–343. [Google Scholar] [CrossRef]
- Guo, X.; Guo, W.; Yang, M.; Sun, Y.; Wang, Y.; Yan, Y.; Zhu, B. Effect of Bacillus additives on fermentation quality and bacterial community during the ensiling process of whole-plant corn silage. Processes 2022, 10, 978. [Google Scholar] [CrossRef]
- ZieliDska, K.; Fabiszewska, A.; StefaDska, I. Different aspects of Lactobacillus inoculants on the improvement of quality and safety of alfalfa silage. Chil. J. Agric. Res. 2015, 75, 298–306. [Google Scholar] [CrossRef]
- Bai, J.; Xu, D.; Xie, D.; Wang, M.; Li, Z.; Guo, X. Effects of antibacterial peptide-producing Bacillus subtilis and Lactobacillus buchneri on fermentation, aerobic stability, and microbial community of alfalfa silage. Bioresour. Technol. 2020, 315, 123881. [Google Scholar] [CrossRef]
- Wu, B.; Ai, J.; Li, T.; Qin, W.; Hu, Z.; Siqin, T.; Wu, T.; Wang, C.; Niu, H. Fermentation quality, aerobic stability, and microbiome structure and function of Caragana korshinskii silage inoculated with/without Lactobacillus rhamnosus or Lactobacillus buchneri. Front. Sustain. Food Syst. 2023, 7, 1255936. [Google Scholar] [CrossRef]
- Liu, Q.; Lindow, S.E.; Zhang, J. Lactobacillus parafarraginis ZH1 producing anti-yeast substances to improve the aerobic stability of silage. Anim. Sci. J. 2018, 89, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, F.; Tabacco, E.; Borreani, G. Lentilactobacillus hilgardii inoculum, dry matter contents at harvest and length of conservation affect fermentation characteristics and aerobic stability of corn silage. Front. Microbiol. 2021, 12, 675563. [Google Scholar] [CrossRef] [PubMed]
- Tabacco, E.; Piano, S.; Revello-Chion, A.; Borreani, G. Effect of Lactobacillus buchneri LN4637 and Lactobacillus buchneri LN40177 on the aerobic stability, fermentation products, and microbial populations of corn silage under farm conditions. J. Dairy Sci. 2011, 94, 5589–5598. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Wang, M.; Ke, W.; Guo, X. Screening of high 1,2-propanediol production by Lactobacillus buchneri strains and their effects on fermentation characteristics and aerobic stability of whole-plant corn silage. Agriculture 2021, 11, 590. [Google Scholar] [CrossRef]
- Filya, I. The effect of Lactobacillus buchneri, with or without homofermentative lactic acid bacteria, on the fermentation, aerobic stability and ruminal degradability of wheat, sorghum and maize silages. J. Appl. Microbiol. 2003, 95, 1080–1086. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-C.; Li, D.-X.; Wang, X.-K.; Lin, Y.-L.; Zhang, Q.; Chen, X.-Y.; Yang, F.-Y. Fermentation quality and aerobic stability of mulberry silage prepared with lactic acid bacteria and propionic acid. Anim. Sci. J. 2019, 90, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Méndez-García, C.; Peláez, A.I.; Mesa, V.; Sánchez, J.; Golyshina, O.V.; Ferrer, M. Microbial diversity and metabolic networks in acid mine drainage habitats. Front. Microbiol. 2015, 6, 475. [Google Scholar]
- Wang, Z.; Tan, Z.; Wu, G.; Wang, L.; Qin, G.; Wang, Y.; Pang, H. Investigation on fermentation characteristics and microbial communities of wheat straw silage with different proportion artemisia argyi. Toxins 2023, 15, 330. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xue, Y.; Xiao, Y.; Te, R.; Wu, X.; Na, N.; Wu, N.; Qili, M.; Zhao, Y.; Cai, Y. Community synergy of lactic acid bacteria and cleaner fermentation of oat silage prepared with a multispecies microbial inoculant. Microbiol. Spectr. 2023, 11, e00705-23. [Google Scholar] [CrossRef]
- Liu, Q.H.; Shao, T.; Zhang, J.G. Determination of aerobic deterioration of corn stalk silage caused by aerobic bacteria. Anim. Feed Sci. Technol. 2013, 183, 124–131. [Google Scholar] [CrossRef]
- Liu, M.; Sun, L.; Wang, Z.; Ge, G.; Jia, Y.; Du, S. Effects of alfalfa hay to oat hay ratios on chemical composition, fermentation characteristics, and fungal communities during aerobic exposure of fermented total mixed ration. Fermentation 2023, 9, 480. [Google Scholar] [CrossRef]
- Liu, B.; Huan, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Li, B.; Zheng, M.; Niu, D.; Zuo, S.; Xu, C. Effects of Pediococcus pentosaceus on fermentation, aerobic stability and microbial communities during ensiling and aerobic spoilage of total mixed ration silage containing alfalfa (Medicago sativa L.). Grassl. Sci. 2020, 66, 215–224. [Google Scholar] [CrossRef]
- Su, R.; Li, F.; Liang, Y.; Sheoran, N.; Bai, J.; Hao, L.; Ke, W.; Hu, C.; Jia, M.; Usman, S.; et al. Responses of microbial community dynamics, co-occurrences, functional shifts, and natural fermentation profiles of Elymus nutans silage to altitudinal gradients. Microbiol. Spectr. 2024, 12, e02516-23. [Google Scholar] [CrossRef]
- Kitamoto, H.K.; Ohmomo, S.; Nakahara, T. Selection of killer yeasts (Kluyveromyces lactis) to prevent aerobic deterioration in silage making. J. Dairy Sci. 1993, 76, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Ciani, M. The zymocidial activity of Tetrapisispora phaffii in the control of Hanseniaspora uvarum during the early stages of winemaking. Lett. Appl. Microbiol. 2010, 50, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Wang, S.; Zhao, J.; Dong, Z.; Li, J.; Liu, Q.; Sun, F.; Shao, T. Effect of ensiling alfalfa with citric acid residue on fermentation quality and aerobic stability. Anim. Feed Sci. Technol. 2020, 269, 114622. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.; Wang, Y.; Rinne, M.; de Oliveira Franco, M.; Li, F.; Lin, Y.; Zhang, Q.; Cai, Y.; Zhang, G. Effects of lactic acid bacteria and molasses on the fermentation quality, in vitro dry matter digestibility, and microbial community of Korshinsk peashrub (Caragana korshinskii Kom.) silages harvested at two growth stages. Grass Forage Sci. 2023, 66, 215–224. [Google Scholar] [CrossRef]
- Arthur Thomas, T. An automated procedure for the determination of soluble carbohydrates in herbage. J. Sci. Food Agric. 1977, 28, 639–642. [Google Scholar] [CrossRef]
- Wang, C.; Han, H.; Sun, L.; Na, N.; Xu, H.; Chang, S.; Jiang, Y.; Xue, Y. Bacterial succession pattern during the fermentation process in whole-plant corn silage processed in different geographical areas of northern China. Processes 2021, 9, 900. [Google Scholar] [CrossRef]
- Feng, Q.; Zhang, J.; Ling, W.; Degen, A.A.; Zhou, Y.; Ge, C.; Yang, F.; Zhou, J. Ensiling hybrid Pennisetum with lactic acid bacteria or organic acids improved the fermentation quality and bacterial community. Front. Microbiol. 2023, 14, 1216722. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.; Pan, G.; Leng, R.; Ni, W.; Yang, J.; Sun, J.; Yu, Z.; Liu, Z.; Xue, Y. Effect of ensiling density and storage temperature on fermentation quality, bacterial community, and nitrate concentration of sorghum-sudangrass silage. Front. Microbiol. 2022, 13, 828320. [Google Scholar] [CrossRef]
- Wang, M.; Xu, S.; Wang, T.; Jia, T.; Xu, Z.; Wang, X.; Yu, Z. Effect of inoculants and storage temperature on the microbial, chemical and mycotoxin composition of corn silage. Asian-Australas. J. Anim. Sci. 2018, 31, 1903–1912. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A tool for visualizing high-throughput microbial community data. GigaScience 2013, 2, 16. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Whole-Plant Corn | SEM |
|---|---|---|
| DM (g/kg FW) | 372.54 | 4.286 |
| CP (g/kg DM) | 79.84 | 0.541 |
| aNDF (g/kg DM) | 375.72 | 18.067 |
| ADF (g/kg DM) | 173.84 | 10.825 |
| WSC (g/kg DM) | 113.70 | 10.085 |
| Ash (g/kg DM) | 41.51 | 3.850 |
| Lactic acid bacteria (lg CFU/g) | 5.64 | 0.106 |
| Coliform bacteria (lg CFU/g) | 5.90 | 0.174 |
| Yeasts (lg CFU/g) | 6.39 | 0.019 |
| Filamentous fungi (lg CFU/g) | <2.00 | - |
| Item | Treatments | SEM | p Value | ||
|---|---|---|---|---|---|
| CK | LB | BL | |||
| DM (g/kg FW) | 371.05 | 374.90 | 374.33 | 3.173 | 0.869 |
| CP (g/kg DM) | 83.65 | 85.98 | 82.65 | 0.613 | 0.904 |
| aNDF (g/kg DM) | 352.37 | 343.40 | 340.00 | 11.480 | 0.766 |
| ADF (g/kg DM) | 171.33 | 171.60 | 169.87 | 6.520 | 0.993 |
| WSC (g/kg DM) | 46.93 | 39.83 | 39.25 | 1.382 | 0.089 |
| Ash (g/kg DM) | 46.53 a | 41.60 ab | 40.90 b | 0.725 | <0.05 |
| Item | D | Treatments | SEM | p Value | ||||
|---|---|---|---|---|---|---|---|---|
| CK | LB | BL | D | T | D*T | |||
| Shannon | 0 | 2.93 Aa | 2.67 Ba | 3.04 Aa | 0.034 | <0.01 | <0.01 | <0.01 |
| 3 | 2.86 Aa | 0.81 Bb | 0.50 Bb | |||||
| 7 | 0.60 Ab | 0.59 Ab | 0.35 Bb | |||||
| Simpson | 0 | 0.10 Bb | 0.14 Ab | 0.10 Bb | 0.019 | <0.01 | <0.01 | <0.01 |
| 3 | 0.10 Bb | 0.70 Aa | 0.77 Aa | |||||
| 7 | 0.63 a | 0.70 a | 0.60 a | |||||
| Chao | 0 | 281.53 Ba | 387.13 Aa | 350.13 Aa | 3.933 | <0.01 | <0.01 | <0.01 |
| 3 | 217.60 Ab | 164.03 Ab | 59.20 Bb | |||||
| 7 | 10.92 Ac | 8.25 Bc | 8.63 Bb | |||||
| Coverage (%) | 0 | 99.83 b | 99.78 c | 99.82 b | 0.000 | <0.01 | 0.464 | 0.130 |
| 3 | 99.88 b | 99.91 b | 99.93 a | |||||
| 7 | 99.99 a | 99.99 a | 99.99 a | |||||
| Item | D | Treatments | SEM | p Value | ||||
|---|---|---|---|---|---|---|---|---|
| CK | LB | BL | D | T | D*T | |||
| Shannon | 0 | 2.68 a | 2.94 a | 2.89 a | 0.037 | <0.01 | <0.01 | <0.01 |
| 3 | 2.91 a | 2.57 b | 2.96 a | |||||
| 7 | 0.99 Bb | 1.53 ABc | 2.09 Ab | |||||
| Simpson | 0 | 0.21 Ab | 0.14 Cc | 0.18 Bab | 0.009 | <0.01 | <0.01 | <0.01 |
| 3 | 0.11 Bb | 0.20 Ab | 0.13 Bb | |||||
| 7 | 0.68 Aa | 0.41 Ba | 0.24 Ba | |||||
| Chao | 0 | 630.02 Ba | 750.19 Aa | 653.06 Ba | 7.387 | <0.01 | <0.05 | <0.01 |
| 3 | 745.05 a | 647.32 b | 684.13 a | |||||
| 7 | 268.08 Ab | 88.27 Bc | 302.61 Ab | |||||
| Coverage (%) | 0 | 99.66 b | 99.62 b | 99.71 b | 0.000 | <0.05 | 0.875 | 0.451 |
| 3 | 99.66 b | 99.67 b | 99.66 b | |||||
| 7 | 99.86 a | 99.91 a | 99.86 a | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, H.; Zhao, M.; Yang, R.; Sun, J.; Yu, Z.; Bai, C.; Xue, Y. Effect of Regulation of Whole-Plant Corn Silage Inoculated with Lactobacillus buchneri or Bacillus licheniformis Regarding the Dynamics of Bacterial and Fungal Communities on Aerobic Stability. Plants 2024, 13, 1471. https://doi.org/10.3390/plants13111471
Yin H, Zhao M, Yang R, Sun J, Yu Z, Bai C, Xue Y. Effect of Regulation of Whole-Plant Corn Silage Inoculated with Lactobacillus buchneri or Bacillus licheniformis Regarding the Dynamics of Bacterial and Fungal Communities on Aerobic Stability. Plants. 2024; 13(11):1471. https://doi.org/10.3390/plants13111471
Chicago/Turabian StyleYin, Hang, Meirong Zhao, Rui Yang, Juanjuan Sun, Zhu Yu, Chunsheng Bai, and Yanlin Xue. 2024. "Effect of Regulation of Whole-Plant Corn Silage Inoculated with Lactobacillus buchneri or Bacillus licheniformis Regarding the Dynamics of Bacterial and Fungal Communities on Aerobic Stability" Plants 13, no. 11: 1471. https://doi.org/10.3390/plants13111471