Abstract
Green leaf volatiles (GLVs) play pivotal roles in plant anti-herbivore defense. This study investigated whether the rice 13-lipoxygense gene OsRCI-1 is involved in GLV production and plant defense in rice. The overexpression of OsRCI-1 (oeRCI lines) in rice resulted in increased wound-induced levels of two prominent GLVs, cis-3-hexen-1-ol and cis-3-hexenal. In a previous study, we found that the overexpression of OsRCI-1 reduced the colonization by the rice brown planthopper (BPH, Nilaparvata lugens) but increased the attractiveness to the egg parasitoid Anagrus nilaparvatae compared to wild-type (WT) plants. This study found that when cis-3-hexen-1-ol, but not cis-3-hexenal, was added to WT plants, it could change the BPH’s colonization preference, i.e., more BPHs preferred to colonize the oeRCI lines. The exogenous application of cis-3-hexen-1-ol or cis-3-hexenal to BPH-infested WT plants could weaken or overturn the preference of A. nilaparvatae for oeRCI lines. However, field experiments revealed that only cis-3-hexenal was attractive to the parasitoid and increased the parasitism rates of BPH eggs. These results indicate that OsRCI-1 is involved in rice GLV production and therefore modulates both direct and indirect defense in rice.
1. Introduction
Lipoxygenase (LOX), the pivotal enzyme within the fatty acid metabolism pathway, can be divided into 9-LOXs and 13-LOXs based on the position of oxygenation in the substrate carbon chain, participating in various aspects of plant growth, development, and responses to both biotic and abiotic stresses [1]. Two branch pathways of the LOX pathway, the allene oxide synthase pathway (AOS) and the hydroperoxide lyase pathway (HPL), have been well studied in induced plant defense against herbivores. Green leaf volatiles (GLVs) including C6 alcohols, aldehydes, and esters, acting through the HPL pathway, play important roles in plant resistance against herbivores, as well as the well-known signal molecules, jasmonates (JAs), produced from the AOS pathway [2].
GLVs, known for their distinct and strong aromas, reminiscent of fresh grass and green leaves, are mostly produced during the early stage of plant development and are rapidly released when cells are mechanically damaged or exposed to herbivores for feeding or oviposition [3]. Research shows that GLVs play a crucial role in both direct and indirect plant defense against insect pests [4]. In particular, emissions of the GLV compound cis-3-hexen-1-ol from plants such as rice (Oryza sativa L.; Gramineae: Oryza), tomato (Solanum lycopersicum L.; Solanaceae: Solanum), and tea (Camellia sinensis (L.) Kuntze; Theaceae: Camellia) have been shown to decrease pest numbers by attracting their natural enemies [5,6,7]. Specifically, studies focusing on key genes of the oxylipin pathway, such as rice OsPLDa4, -a5, and OsHPL3, have shown that cis-3-hexen-1-ol, released by rice plants, acts as a repellent against the rice brown planthopper (BPH, Nilaparvata lugens Stål; Hemiptera: Delphacidae), while attracting its natural enemy [5,8]. In contrast, the release of cis-3-hexenal from rice plants was found to reduce their resistance to the white-backed planthopper (WBPH, Sogatella furcifera Horváth; Hemiptera: Delphacidae) [9].
In rice, 14 LOX genes have been identified, of which seven genes have been cloned and characterized, including OsRLL/OsHI-LOX, LOX-2, LOX-3, Osr9-LOX1, OsRCI-1, and OsLOX1 [10,11,12,13,14,15]. The resistance of rice against the rice brown planthopper was reduced in the antisense-suppressed lines of OsLOX1, while it was enhanced in the overexpression lines, and OsLOX1 was related to the synthesis of JA and cis-3-hexen-1-ol [16]. Zhou et al. (2009) cloned the OsHI-LOX gene from the rice 13-LOXs, and the antisense-suppressed mutant rice exhibited enhanced resistance to piercing-sucking brown planthoppers but reduced resistance to chewing insects like the striped stem borer (Chilo suppressalis Walker; Lepidoptera: Pyralidae) and rice leaf folder (Cnaphalocrocis medinalis Guenée; Lepidoptera: Pyralidae), but they found that OsHI-LOX was only involved in the synthesis of JA, rather than the production of GLVs [13]. OsRCI-1 has been identified as a member of the 13-LOX family [15]. OsRCI-1 is induced by compounds such as benzo-(1,2,3)-thiadiazole-7-carbothioic S-methyl ester (BTH) and benzo-(1,2,3)-thiadiazole-7-carbothioic S-methyl ester (INA). The overexpression of OsRCI-1 in rice transiently resulted in the upregulation of the pathogenesis-related protein PR1 gene [17]. OsRCI-1 is also reported to be involved in BPH-induced JA bursts and plays a positive role in rice defense against BPH [18].
The biosynthesis of GLVs is orchestrated through the oxygenation of acyl groups by pathway-specific lipoxygenases; subsequent cleavage reactions facilitated by hydroperoxide lyases contribute to the formation of these characteristic volatile compounds [19]. In this study, we used overexpression OsRCI-1 rice (oeRCI lines) [18] to investigate whether OsRCI-1 was involved in rice GLV production and whether these GLVs played a role in the regulation of BPH colonization and the behavior of its egg parasitoid wasp, Anagrus nilaparvatae Pang et Wang (Hymenoptera: Mymaridae). We hypothesized that the overexpression of OsRCI-1 in rice would enhance the production of GLVs, therefore playing a role in regulating BPH colonization and the behavior of its egg parasitoid A. nilaparvatae.
2. Results
2.1. Overexpressing OsRCI-1 Enhances Production of Mechanical Damage-Induced GLVs in Rice
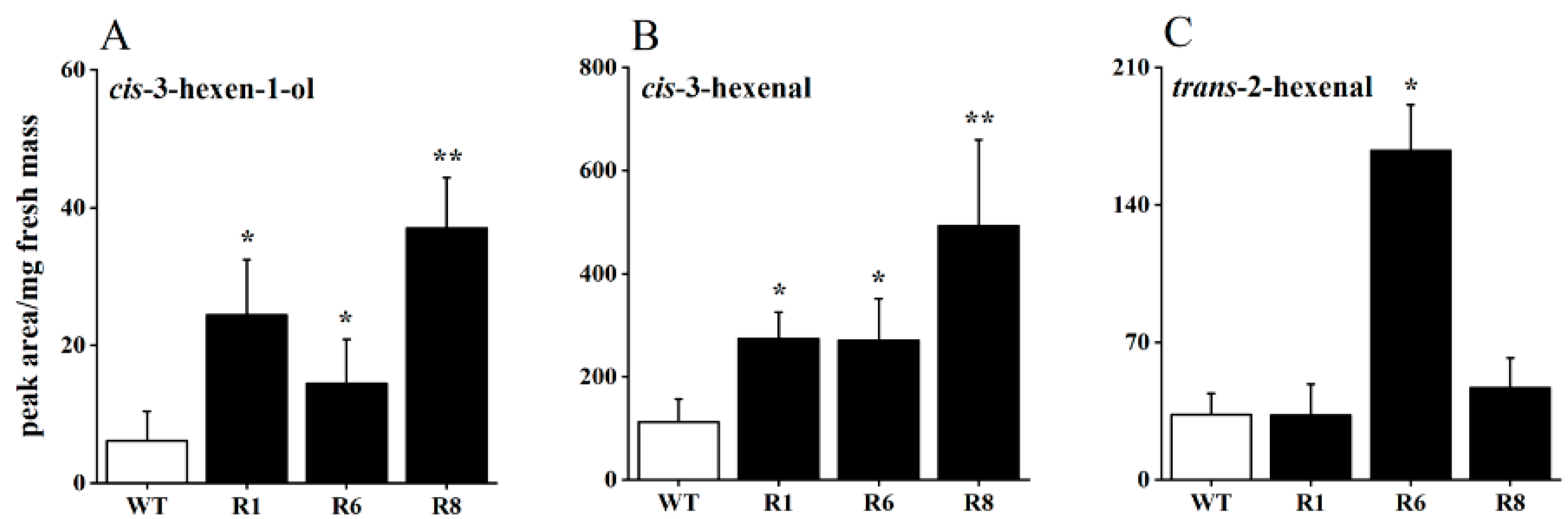
To investigate the impact of OsRCI-1 on the production of rice GLVs, the levels of GLVs from OsRCI-1 overexpression rice (oeRCI lines) and wild-type rice plants were estimated after mechanical damage treatment. After the mechanical damage treatment, the content of three GLVs showed varying degrees of increase in the overexpression lines (Figure 1A–C). It is interesting that, in comparison to WT rice plants, the content of cis-3-hexen-1-ol significantly rose in the R1, R6, and R8 lines, with increases of 3.98, 2.36, and 6.03 times, respectively (Figure 1A). Another GLV, cis-3-hexenal, was also significantly increased in these three oeRCI lines, with an increase of 2.41–4.37 times (Figure 1B). However, the content of trans-2-hexenal was only significantly increased in the R6 line and not in the R1 and R8 lines (Figure 1C). These results indicate that the overexpression of OsRCI-1 may be involved in rice GLV production.
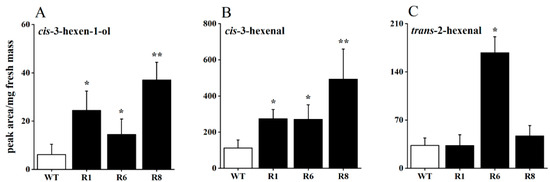
Figure 1.
The content of cis-3-hexen-1-ol (A), cis-3-hexenal (B), and trans-2-hexenal (C) in wild-type (WT) and oeRCI lines (R1, R6, and R8) after mechanical damage treatment. Asterisks indicate significant differences between pairs of samples within time points (*, p < 0.05; **, p < 0.01; Student’s t-test).
2.2. Effects of cis-3-hexen-1-ol and cis-3-hexenal on Feeding and Oviposition Preferences of BPH
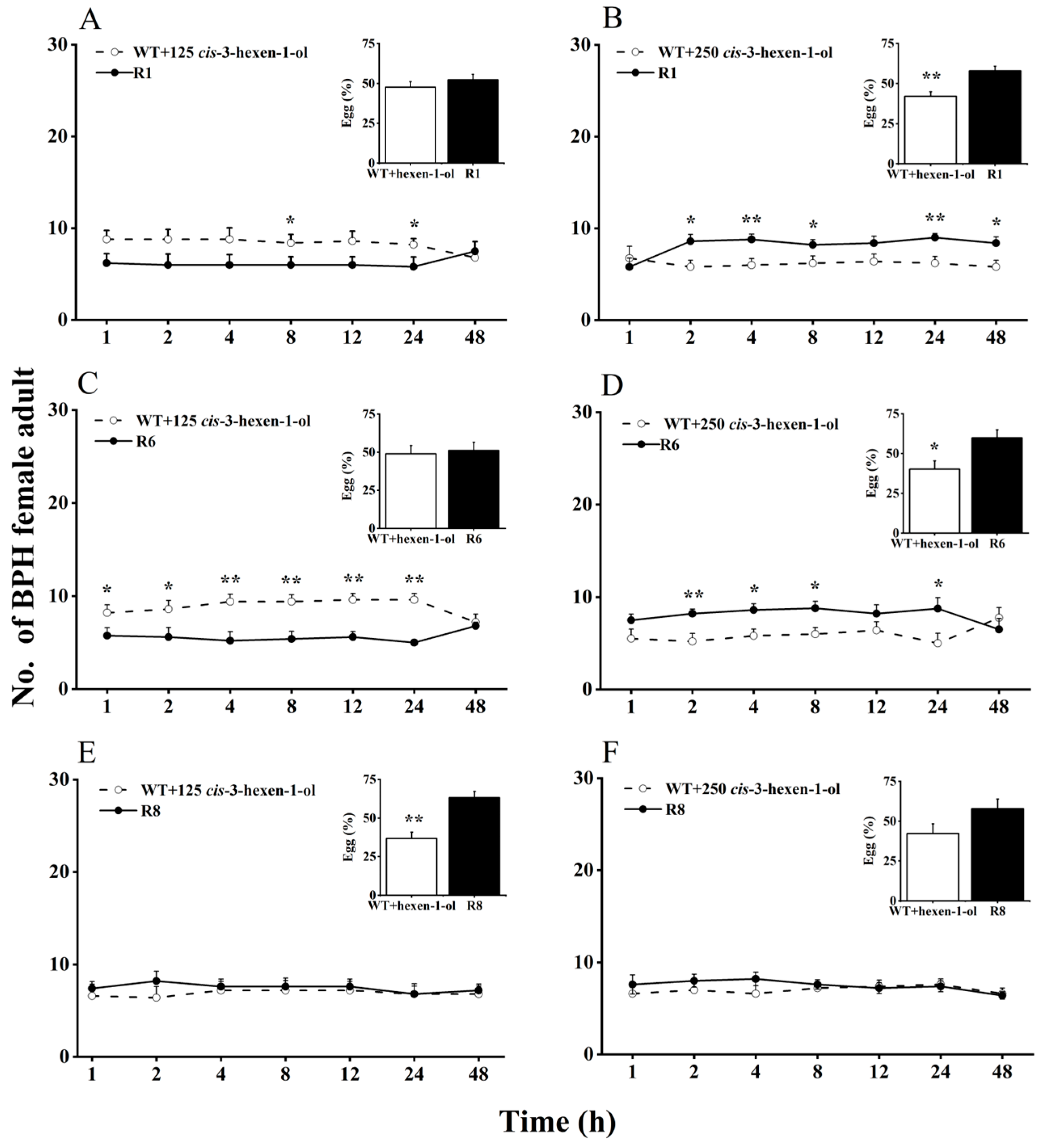
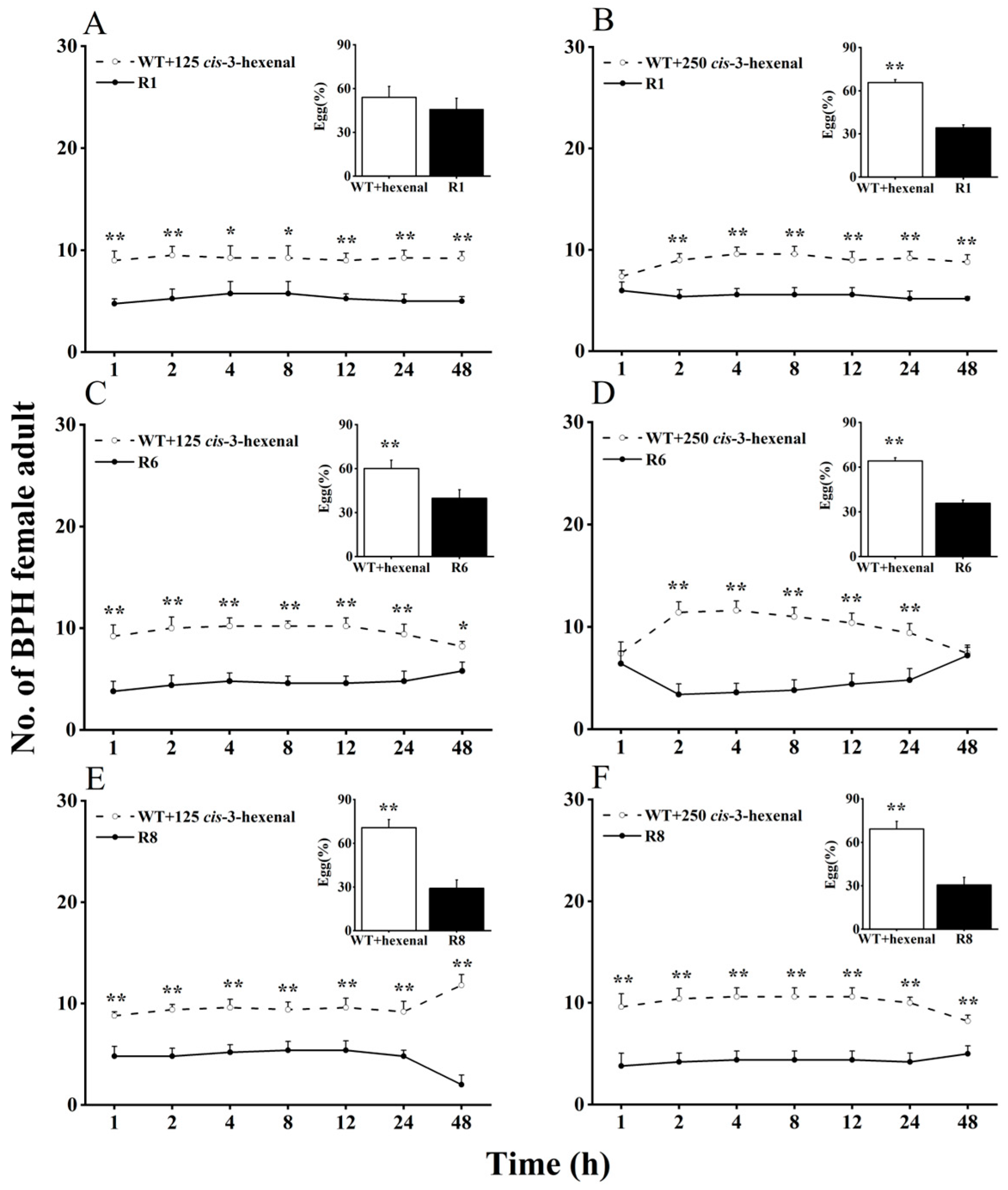
Here, we evaluated BPH selection in WT rice plants supplemented with cis-3-hexen-1-ol or cis-3-hexenal, compared with oeRCI lines, to assess the activity of GLVs on BPH’s feeding and egg laying preference (Figure 2 and Figure 3). When 125 nmol of cis-3-hexen-1-ol was applied to the leaf sheaths of WT plants, BPH still showed a preference to feed and lay eggs on WT plants over oeRCI lines (R1 or R6) (Figure 2A,C). The preference of BPH was not evident in the comparison between cis-3-hexen-1-ol-treated WT plants and the R8 line (Figure 2E). In addition, when 250 nmol of cis-3-hexen-1-ol was applied to WT plants, a reversal in BPH’s feeding and oviposition preference was observed; BPH escaped feeding and oviposition on the R1 or R6 lines (Figure 2B,D). However, the application of either 125 or 250 nmol of another GLV, cis-3-hexenal, on the leaf sheaths of WT rice plants did not affect BPH, preferring cis-3-hexenal-treated WT plants (p < 0.05, Figure 3A–F) as well as untreated WT plants. These findings suggest that cis-3-hexen-1-ol but not cis-3-hexenal showed a repellent effect on the female adult BPH.
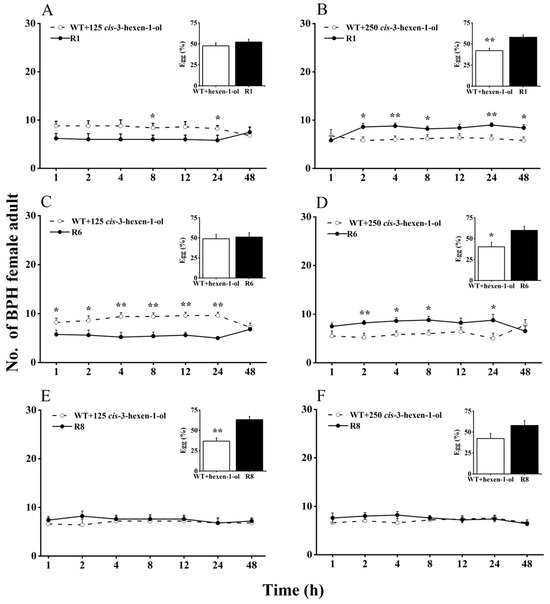
Figure 2.
Effects of 125 (A,C,E) or 250 nmol (B,D,F) cis-3-hexen-1-ol on feeding and oviposition preference of BPH female adults (+SE, n = 5) in WT and OsRCI-1 overexpression lines. Asterisks denote significant differences between pairs of groups within each time point (*, p < 0.05; **, p < 0.01; Student’s t-test).
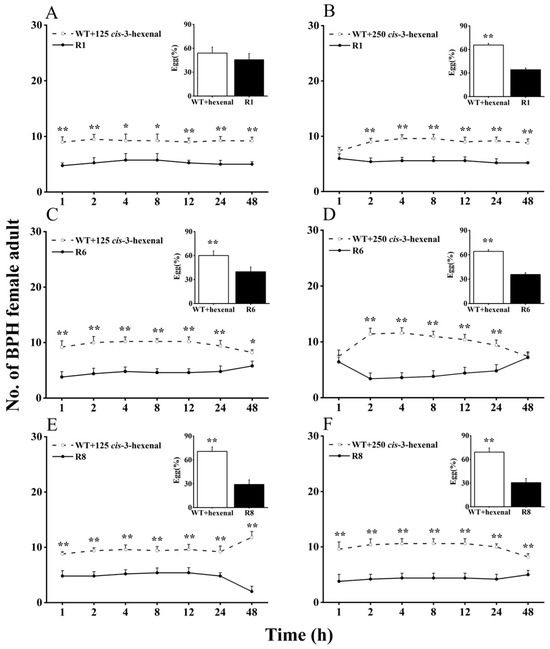
Figure 3.
Effects of 125 (A,C,E) or 250 nmol (B,D,F) cis-3-hexenal on feeding and oviposition preference of BPH female adults (+SE, n = 5) in WT and OsRCI-1 overexpression lines. Asterisks denote significant differences between pairs of members within time points (*, p < 0.05; **, p < 0.01; Student’s t-test).
2.3. GLVs Modify Rice’s Attractiveness to Parasitoids after BPH Infestation
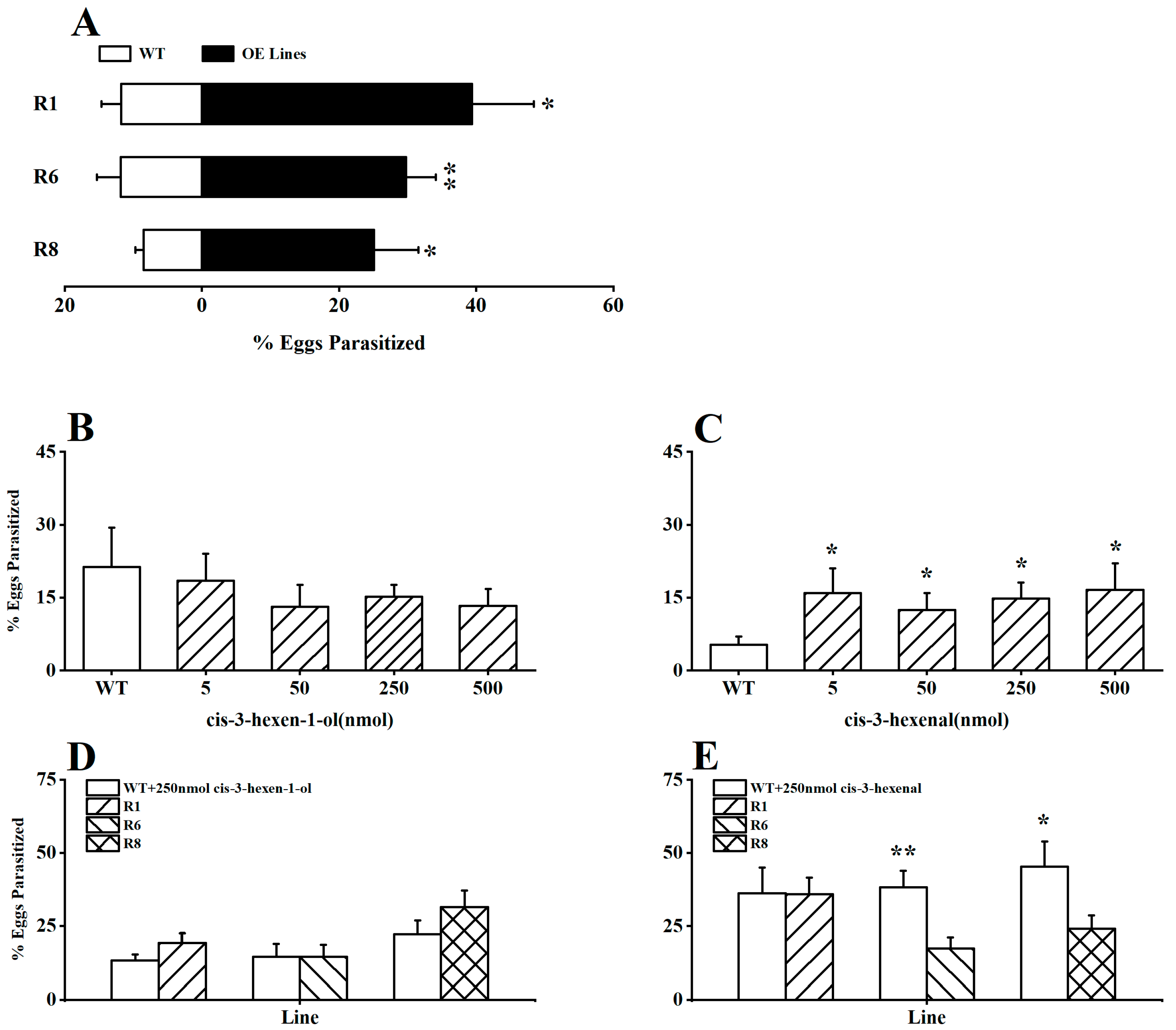
To explore the potential impact of GLVs in rice on the parasitic preference of the parasitoid A. nilaparvatae (BPH egg parasitoid), we initially introduced the parasitoids into the OsRCI-1 overexpression oeRCI lines and WT plants infested with BPH in a laboratory setting. It is interesting that the parasitism rates of BPH eggs on OsRCI-1-overexpressing plants were significantly higher than those on WT plants (p < 0.05), with the parasitism rates of BPH eggs on the R1, R6, and R8 lines being 3.35, 2.52, and 2.95 times higher than those on WT plants, respectively (Figure 4A). Furthermore, whether cis-3-hexen-1-ol or cis-3-hexen-al could increase the parasitism rates of BPH eggs on WT plants was assessed in the field. The application of cis-3-hexen-1-ol (5–500 nmol) on WT plants showed no significant difference in the parasitism rates of BPH eggs between treated and untreated WT plants (Figure 4B); however, the application of cis-3-hexenal at concentrations ranging from 5 to 500 nmol significantly elevated the parasitism rates of BPH eggs on WT plants in the field (p < 0.05; Figure 4C). The application of 250 nmol cis-3-hexen-1-ol to WT plants had no significant effect on the BPH egg parasitism rates in comparison to oeRCI lines in indoor experiments (p > 0.05; Figure 4D); unexpectedly, applying 250 nmol cis-3-hexenal to WT plants could deactivate or reverse A. nilaparvatae’s preference, parasitizing BPH eggs on oeRCI lines (Figure 4E), and the parasitism rates of BPH eggs on cis-3-hexenal-treated WT plants were apparently higher than those on the R6 and R8 rice plants (p < 0.05; Figure 4E).
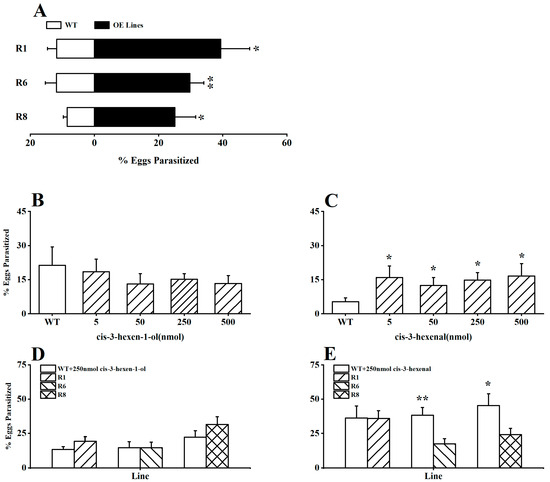
Figure 4.
(A) The percentage (+SE, n = 7) of BPH eggs parasitized by A. nilaparvatae on R1, R6, R8, and WT plants. (B,C) The mean (+SE, n = 7) parasitism rates (%) of BPH eggs on WT plants treated with 5–500 nmol cis-3-hexen-1-ol or cis-3-hexenal in the field. (D,E) The mean (+SE, n = 7) parasitism rates (%) of BPH eggs in indoor experiments, with 250 nmol cis-3-hexen-1-ol or cis-3-hexenal applied to WT plants versus oeRCI lines. Asterisks indicate significant differences in R lines compared with WT plants (*, p < 0.05; **, p < 0.01; Student’s t-test).
3. Discussion
The expression of the OsRCI-1 gene in rice can be rapidly and consistently induced by Chilo suppressalis and BPH infestation. C. suppressalis feeding on OsRCI-1-silenced lines increased in body weight by 1.58–2.15 times compared to feeding on WT plants [20]. The overexpression of OsRCI-1 not only increased the levels of BPH-induced JA and JA-Ile but also decreased the feeding and oviposition preferences of BPH female adults, ultimately reducing BPH’s performance [18]. In this study, we found that the overexpression of OsRCI-1 increased the levels of GLVs, including cis-3-hexen-1-ol and cis-3-hexenal. The increased GLVs, especially cis-3-hexenal and cis-3-hexen-1-ol (but not cis-3-hexenal alone), significantly regulated the BPH’s feeding and oviposition performance. Additionally, we observed that the wasp A. nilaparvatae preferred to parasitize BPH eggs on OsRCI-1 overexpression rice plants. This preference of the wasp could be partly attributed to the increased levels of GLVs produced by the OsRCI-1 overexpression plants. In conclusion, the rice lipoxygenase OsRCI-1 is involved in GLV production of the HPL branch pathway, as well as the JA generation of the AOS branch pathway, in the rice jsamonate pathway, and the GLVs play an important role in protecting rice from invasive pests.
The different GLVs showed disparate biological functions in the interactions between plants, herbivores, and the enemies of herbivores. In the OsRCI-1 overexpression lines, the levels of cis-3-hexen-1-ol and cis-3-hexenal but not trans-2-hexenal significantly increased, but it was found that cis-3-hexen-1-ol and cis-3-hexenal had different effects on the behavior of BPH and A. nilaparvatae (Figure 2, Figure 3 and Figure 4). The supplementation of cis-3-hexen-1-ol in WT rice plants could apparently alter the feeding and oviposition preferences of BPH adults (Figure 2), working as a repellent, and its repellent activity appeared to be dependent on its concentration (Figure 2). However, when adding cis-3-hexen-1-ol to WT rice plants in the field, no difference was found in the parasitism rates of BPH eggs on control rice and cis-3-hexen-1-ol-treated WT plants (Figure 4). Other studies showed that cis-3-hexen-1-ol is involved in regulating direct and indirect defense pathways in tea plants [21] and can activate defense responses against insect pests in plants like maize [22,23,24]. In contrast, cis-3-hexenal emitted from OsRCI-1-overexpressed rice lines did not change the feeding and oviposition preference of BPH adults (Figure 3), but significantly increased the parasitism rate of BPH eggs (Figure 4). Therefore, the rice lipoxygenase gene OsRCI-1 could repel BPH by releasing cis-3-hexen-1-ol, functioning as a “push” strategy, and attract A. nilaparvatae by releasing cis-3-hexenal, therefore increasing the parasitism rate of BPH eggs, functioning as a “pull” strategy. Similar phenomena have been reported; Lamy et al. [25] found that the volatile dimethyl disulfide exhibited a “push” strategy by reducing the eggs laid on broccoli by the cabbage root fly (Delia radicum Linnaeus; Diptera: Anthomyiidae). Meanwhile, cis-3-hexenyl acetate exhibited a “pull” strategy by attracting the natural enemy of D. radicum.
The lipoxygenase pathway genes can regulate the emission of GLVs; therefore, they play an important role in the direct and indirect defense responses of rice. For example, compared to wild-type (WT) plants, mutants of antisense-suppressed phospholipase D (PLD) genes (OsPLDa4 and a5) exhibited reduced levels of trypsin inhibitors and GLVs, specifically cis-3-hexenal and cis-3-hexen-1-ol [5]. This resulted in the increased growth and reproduction of BPH and C. suppressalis, along with decreased attraction to the natural enemy of C. suppressalis. Applying these two GLVs externally restored the resistance of the mutants to BPH and C. suppressalis [5]. The loss-of-function or antisense inhibition mutants of the rice hydrogen peroxide lyase gene (OsHPL3) exhibited reduced levels of cis-3-hexen-1-ol and cis-3-hexenal, resulting in decreased resistance to BPH but increased resistance against WBPH, and also increased attraction to the parasitoid A. nilaparvatae [8,26]. In this study, we found that the overexpression of the rice lipoxygenase gene OsRCI-1 increased the levels of cis-3-hexen-1-ol and cis-3-hexenal (Figure 1), therefore reducing the feeding and oviposition preferences of BPH but enhancing the attraction of A. nilaparvatae (Figure 4E). These findings suggest that elevated levels of green leaf volatiles (cis-3-hexen-1-ol and cis-3-hexenal) contribute to enhancing the direct and indirect resistance of rice against BPH.
In summary, our study provides compelling evidence for the involvement of OsRCI-1 in regulating the release of GLVs including cis-3-hexenal and cis-3-hexen-1-ol in rice; OsRCI-1 is another LOX gene that contributes to both JA and GLV synthesis in rice as OsLOX1 [16]. OsRCI-1 overexpression in rice lines could significantly reduce BPH’s feeding and oviposition preferences and increase the attractiveness to egg parasitoid A. nilaparvatae [18]. Another new insight is the increase in cis-3-hexenal in rice plants, which functions as a repellent to BPH, and cis-3-hexen-1-ol in rice functions as an attractant towards the herbivore’s natural enemy. These results highlight the role of GLVs mediated by OsRCI-1 in regulating both the direct and indirect defenses of rice against BPH. The rice lipoxygenase gene OsRCI-1 and its overexpression in rice have potential applications in pest-resistant rice breeding.
4. Materials and Methods
4.1. Rice Materials
The rice genotype Xiushui 11 (WT) and three OsRCI-1-overexpressing transgenic oeRCI lines (R1, R6, and R8) [18] were used in this study. The rice seeds were first germinated in a growth chamber and then transplanted into 25-liter plastic pots filled with rice culture solution. Both seed germination and plant growth were monitored under controlled conditions (27 ± 2 °C, relative humidity 70–80%, and light–dark = 16:8). Rice seedlings at 35–40 days old were used for experiments.
4.2. Insect Materials
BPH populations were collected from rice fields in Lin’an, Zhejiang, China, and maintained on TN1 rice seedlings for at least ten generations in a controlled climate room at 27 °C ± 2 °C, 70–80% relative humidity, and a 12 h light phase, as reported by Liao et al. [18]. TN1 rice plants with BPH eggs were then placed in a rice field at Zhejiang A & F University to attract the parasitoid A. nilaparvatae. In the growth chamber, BPH female adults lay eggs on rice seedlings and then we offered these seedlings to the parasitoid.
4.3. Reagents and Instruments
The reagents used in this study were cis-3-hexen-1-ol, cis-3-hexenal, trans-2-hexenal, and lanolin, which were purchased from Shanghai Aladdin Biochemical Technology Co., Ltd., Shanghai, China. The instruments used were the RXZ-380B intelligent growth chamber (Ningbo Jiangnan Instrument Factory, Ningbo, Zhejiang, China) and the Motic binocular dissecting microscope (Meike Audi Industrial Group Co., Ltd., Xiamen, Fujian, China).
4.4. GLV Analysis
GLVs (cis-3-hexenal, cis-3-hexen-1-ol, and trans-2-hexenal) were analyzed with a portable gas analyzer (zNose 4200, Electronic Sensor Technology, Newbury Park, CA, USA) using the method described by Zhou et al. [6]. Rice leaves (0.02 g) from the three overexpression lines R1, R6, and R8 and their wild-type XS11 were cut into small pieces (about 2 mm long) and immediately placed in a 50 mL glass chamber (Jiangyin Elite Experimental Instrument Co., Ltd., Jiangyin, Jiangsu, China), respectively; after 5 min, GLVs were sampled for 40 s and analyzed by the zNose 4200. At least eight replicates were carried out.
4.5. BPH Bioassays
To estimate the impact of GLVs, specifically cis-3-hexen-1-ol or cis-3-hexenal, on the colonization and oviposition preference of BPH between the WT and oeRCI rice lines, pots with two plants were used. One plant was an oeRCI line treated with 10 μL lanolin and the other was a WT rice plant treated with 10 μL lanolin containing 125 or 250 nmol GLVs (cis-3-hexen-1-ol or cis-3-hexenal). The pots were confined within a glass cylinder (diameter, 4 cm; height, 8 cm; with 24 small holes (diameter 0.8 mm) evenly distributed on the cylinder wall) sealed with a sponge, into which 15 gravid BPH females were introduced. The number of BPHs on each plant was recorded at 1, 2, 4, 8, 12, 24, and 48 h. After 48 h, the number of BPH eggs on each plant was counted under a dissecting microscope. The experiment was repeated at least five times for each pair of plants.
4.6. Field Experiment
To evaluate the role of GLVs (cis-3-hexen-1-ol or cis-3-hexenal) in rice–parasitoid interactions in the field, WT plants (one plant per pot) were individually treated with 0, 5, 50, 250, or 500 nmol cis-3-hexen-1-ol or cis-3-hexenal in 10 μL lanolin and were infested with 8 BPH females for 24 h; then, the BPHs were removed as described by Liao et al. [19]. Five potted plants (each of 0, 5, 50, 250, or 500 nmol cis-3-hexen-1-ol- or cis-3-hexenal-treated WT plants) were placed and arranged within a 40 cm circular radius as located in a rice field. The location was 1 m from the edge of the rice field, and the distance between each location was kept at 2 m. Two days later, the plants were transferred to a controlled climate room at 27 ± 2 °C with a 14 h light phase and 70% relative humidity for five days. Then, the total number of BPH eggs and eggs parasitized by the wasps (these eggs became red) were counted. Each treatment was repeated at least seven times.
4.7. Wasp Bioassays in Controlled Climate Chamber
The WT and oeRCI rice lines were infested with 12 BPH females and oviposited for 12 h, respectively, and then the BPHs were removed. Pots with two plants (one oeRCI line with BPH eggs vs. a WT plant with BPH eggs) were kept 1 cm apart and were confined within glass cylinders (diameter 4 cm, height 8 cm, with 24 small holes), into which six newly emerged A. nilaparvatae wasps (3♀ and 3♂) were introduced, as described by Liao et al. [19]. After 24 h, the parasitoid wasps were removed, and the rice plants were kept in a controlled climate room at 27 ± 2 °C with a 14 h light phase and 70% relative humidity for five days. Then, the total number of BPH eggs and eggs parasitized by the wasps were counted. Each treatment was repeated at least seven times.
Author Contributions
Conceptualization, K.M., G.Z. and W.X.; methodology, H.Z.; software, K.M. and C.L.; validation, C.L. and Y.W.; formal analysis, K.M. and C.L.; investigation, K.M.; resources, G.Z. and W.X.; data curation, K.M.; writing—original draft preparation, G.Z., Y.L., K.M., W.X. and H.Z.; writing—review and editing, G.Z., Y.L., K.M., W.X. and C.L.; visualization, K.M. and W.X.; supervision, G.Z., Y.L. and W.X.; project administration, G.Z.; funding acquisition, G.Z., Y.L. and W.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32072508, 31572020), the “Pioneer” and “Leading Goose” R&D Program of Zhejiang Province (2022C02020), the “San Nong Jiu Fang” Program of Zhejiang Provincial Department of Agriculture and Rural Affairs (2022SNJF011), and the Natural Science Foundation of Zhejiang Province (LQ23C140006; LQ23C140005). This project was also supported by the Student Scientific Research Training Program of Zhejiang A&F University (2023KX008).
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors thank the anonymous reviewers for their constructive comments on a draft of the paper.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Feussner, I.; Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol. 2002, 53, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Greco, M.; Chiappetta, A.; Bruno, L.; Bitonti, M.B. In Posidonia oceanica cadmium induces changes in DNA methylation and chromatin patterning. J. Exp. Bot. 2012, 63, 695–709. [Google Scholar] [CrossRef]
- Selli, S.; Kelebek, H.; Ayseli, M.T.; Tokbas, H. Characterization of the most aroma-active compounds in cherry tomato by application of the aroma extract dilution analysis. Food Chem. 2014, 165, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Ameye, M.; Allmann, S.; Verwaeren, J.; Smagghe, G.; Haesaert, G.; Schuurink, R.C.; Audenaert, K. Green leaf volatile production by plants: A meta-analysis. New Phytol. 2018, 220, 666–683. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Zhou, G.; Yang, L. The chloroplast-localized phospholipases D α4 and α5 regulate herbivore-induced direct and indirect defense in rice. Plant Physiol. 2011, 157, 1987–1999. [Google Scholar] [CrossRef] [PubMed]
- Xin, Z.; Zhang, L.; Zhang, Z.; Chen, Z.; Sun, X. A tea hydroperoxide lyase gene, CsiHPL1, regulates tomato defense response against Prodenia litura (Fabricius) and Alternaria alternata f. sp. Lycopersici by modulating green leaf volatiles (GLVs) release and jasmonic acid (JA) gene expression. Plant Mol. Biol. Rep. 2013, 32, 62–69. [Google Scholar]
- Yang, F.; Zhang, Q.; Yao, Q.; Chen, G.; Tong, H.; Zhang, J.; Li, C.; Su, Q.; Zhang, Y. Direct and indirect plant defenses induced by (Z)-3-hexenol in tomato against whitefly attack. J. Pest Sci. 2020, 93, 1243–1254. [Google Scholar] [CrossRef]
- Tong, X.; Qi, J.; Zhu, X.; Mao, B.; Zeng, L.; Wang, B.; Li, Q.; Zhou, G.; Xu, X.; Lou, Y.; et al. The rice hydroperoxide lyase OsHPL3 functions in defense responses by modulating the oxylipin pathway. Plant J. 2012, 71, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhou, G.; Xin, Z.; Ji, R.; Lou, Y. (Z)-3-hexenal, one of the green leaf volatiles, increases susceptibility of rice to the white-backed planthopper Sogatella furcifera. Plant Mol. Biol. 2014, 33, 377–387. [Google Scholar] [CrossRef]
- Peng, Y.L.; Shirano, Y.; Ohta, H.; Hibino, T.; Tanaka, K.; Shibata, D. A novel lipoxygenase from rice: Primary structure and specific expression upon incompatible infection with rice blast fungus. J. Biol. Chem. 1994, 269, 3755–3761. [Google Scholar] [CrossRef]
- Ohta, H.; Shirano, Y.; Tanaka, K.; Morita, Y.; Shibata, D. cDNA cloning of rice lipoxygenase L-2 and characterization using an active enzyme expressed from the cDNA in Escherichia coli. Eur. J. Biochem. 1992, 206, 331–336. [Google Scholar] [CrossRef]
- Suzuki, Y.; Miura, K.; Shigemune, A.; Sasahara, H.; Ohta, H.; Uehara, Y.; Ishikawa, T.; Hamada, S.; Shirasawa, K. Marker-assisted breeding of a LOX-3-null rice line with improved storability and resistance to preharvest sprouting. Theor. Appl. Genet. 2015, 128, 1421–1430. [Google Scholar] [CrossRef]
- Zhou, G.; Qi, J.; Ren, N.; Cheng, J.; Erb, M.; Mao, B.; Lou, Y. Silencing OsHI-LOX makes rice more susceptible to chewing herbivores, but enhances resistance to a phloem feeder. Plant J. 2009, 60, 638–648. [Google Scholar] [CrossRef]
- Mizuno, K.; Iida, T.; Takano, A.; Yokoyama, M.; Fujimura, T. A new 9-lipoxygenase cDNA from developing rice seeds. Plant Cell Physiol. 2003, 44, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Schaffrath, U.; Zabbai, F.; Dudler, R. Characterization of RCI-1, a chloroplastic rice lipoxygenase whose synthesis is induced by chemical plant resistance activators. Eur. J. Biochem. 2000, 267, 5935–5942. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Shen, W.; Liu, L.; Jiang, L.; Liu, Y.; Su, N.; Wan, J. A novel lipoxygenase gene from developing rice seeds confers dual position specificity and responds to wounding and insect attack. Plant Mol. Biol. Rep. 2008, 66, 401–414. [Google Scholar] [CrossRef]
- Zabbai, F.; Jarosch, B.; Schaffrath, U. Over-expression of chloroplastic lipoxygenase RCI1 causes PR1 transcript accumulation in transiently transformed rice. Physiol. Mol. Plant Pathol. 2004, 64, 37–43. [Google Scholar] [CrossRef]
- Liao, Z.; Wang, L.; Li, C.; Cao, M.; Wang, J.; Yao, Z.; Zhou, S.; Zhou, G.; Zhang, D.; Lou, Y. The lipoxygenase gene OsRCI-1 is involved in the biosynthesis of herbivore-induced JAs and regulates plant defense and growth in rice. Plant Cell Environ. 2022, 45, 2827–2840. [Google Scholar] [CrossRef]
- Matsui, K. Green leaf volatiles: Hydroperoxide lyase pathway of oxylipin metabolism. Curr. Opin. Plant Biol. 2006, 9, 274–280. [Google Scholar] [CrossRef]
- Yao, Z.; Cao, M.; Wang, X.; Miao, J.; Shen, L.; Deng, J.; Wu, H.; Xu, Z.; Lou, Y.; Zhou, G. Rice lipoxygenase gene OsRCI-1 mediated pathway positively regulates resistance to rice striped stem borer. J. Environ. Entomol. 2014, 36, 507–515. [Google Scholar]
- Liao, Y.; Tan, H.; Jian, G.; Zhou, X.; Huo, L.; Jia, Y.; Zeng, L.; Yang, Z. Herbivore-induced (Z)-3-hexen-1-ol is an airborne signal that promotes direct and indirect defenses in tea (Camellia sinensis) under light. J. Agric. Food Chem. 2021, 3, 43. [Google Scholar] [CrossRef] [PubMed]
- Engelberth, J.; Alborn, H.T.; Schmelz, E.A.; Tumlinson, J.H. Airborne signals prime plants against insect herbivore attack. Proc. Natl. Acad. Sci. USA 2004, 101, 1781–1785. [Google Scholar] [CrossRef] [PubMed]
- Engelberth, J.; Contreras, C.F.; Dalvi, C.; Li, T.; Engelberth, M.; Heil, M. Early transcriptome analyses of Z-3-hexenol-treated Zea mays revealed distinct transcriptional networks and anti-herbivore defense potential of green leaf volatiles. PLoS ONE 2013, 8, e77465. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Fokar, M.; Abd, H.; Zhang, H.; Allen, R.D.; Pare, P.W. (Z)-3-Hexenol induces defense genes and downstream metabolites in maize. Planta 2005, 220, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Lamy, F.; Dugravot, S.; Cortesero, A.M.; Chaminade, V.; Faloya, V.; Poinsot, D. One more step toward a push-pull strategy combining both a trap crop and plant volatile organic compounds against the cabbage root fly Delia radicum. Environ. Sci. Pollut. Res. Int. 2018, 25, 29868–29879. [Google Scholar] [CrossRef]
- Wei, J.; Wang, L.; Zhu, J.; Zhang, S.; Nandi, O.I.; Kang, L. Plants attract parasitic wasps to defend themselves against insect pests by releasing hexenol. PLoS ONE 2007, 2, e852. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).