

Phytochemical Profiles and Cytotoxic Activity of Bursera fagaroides (Kunth) Engl. Leaves and Its Callus Culture
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Obtaining Leaves from the Cultivation of Cuttings
2.2. Callus Induction
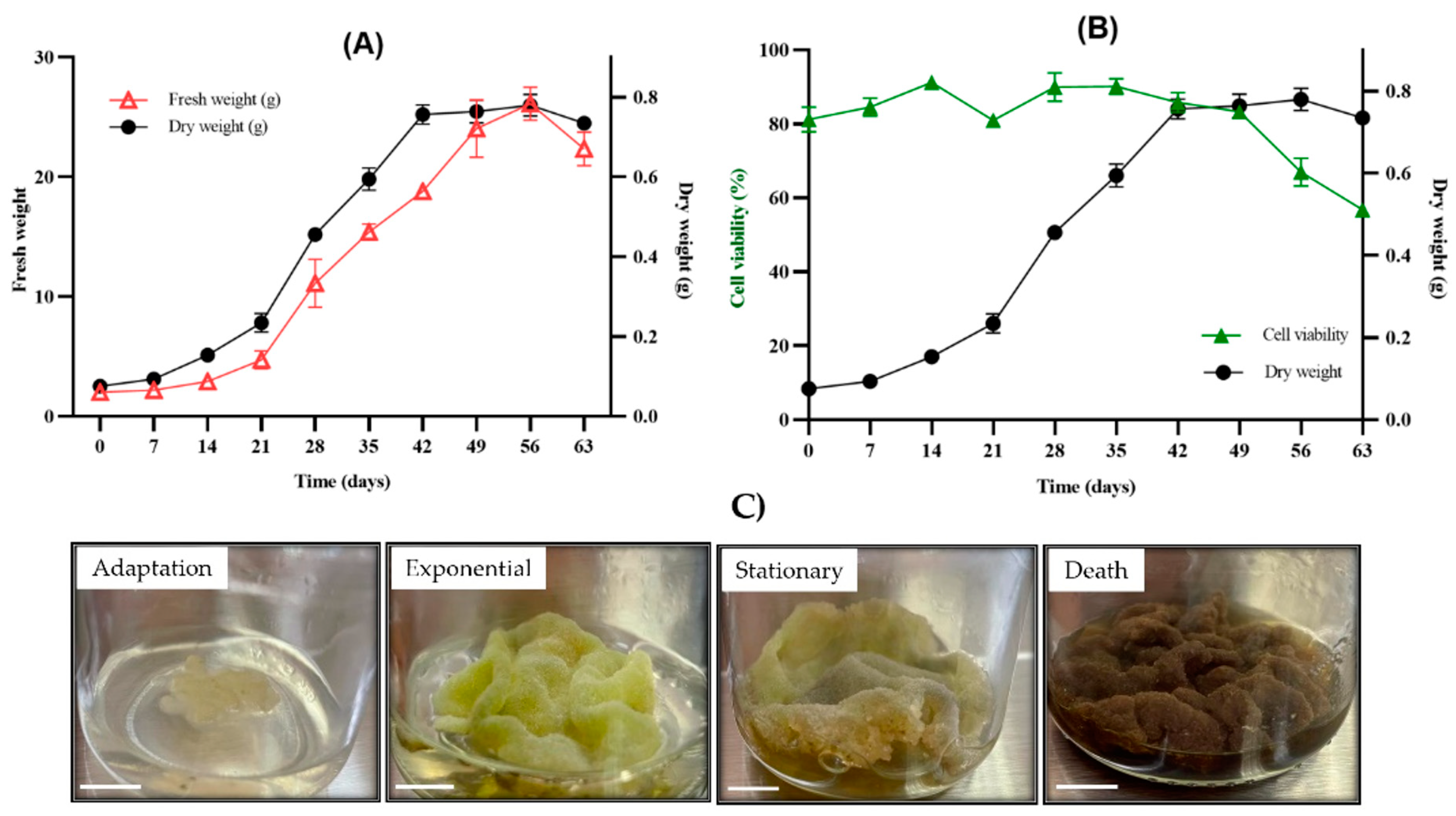
2.3. Growth Kinetics, Percentage of Viability and Appearance of the Callus Culture
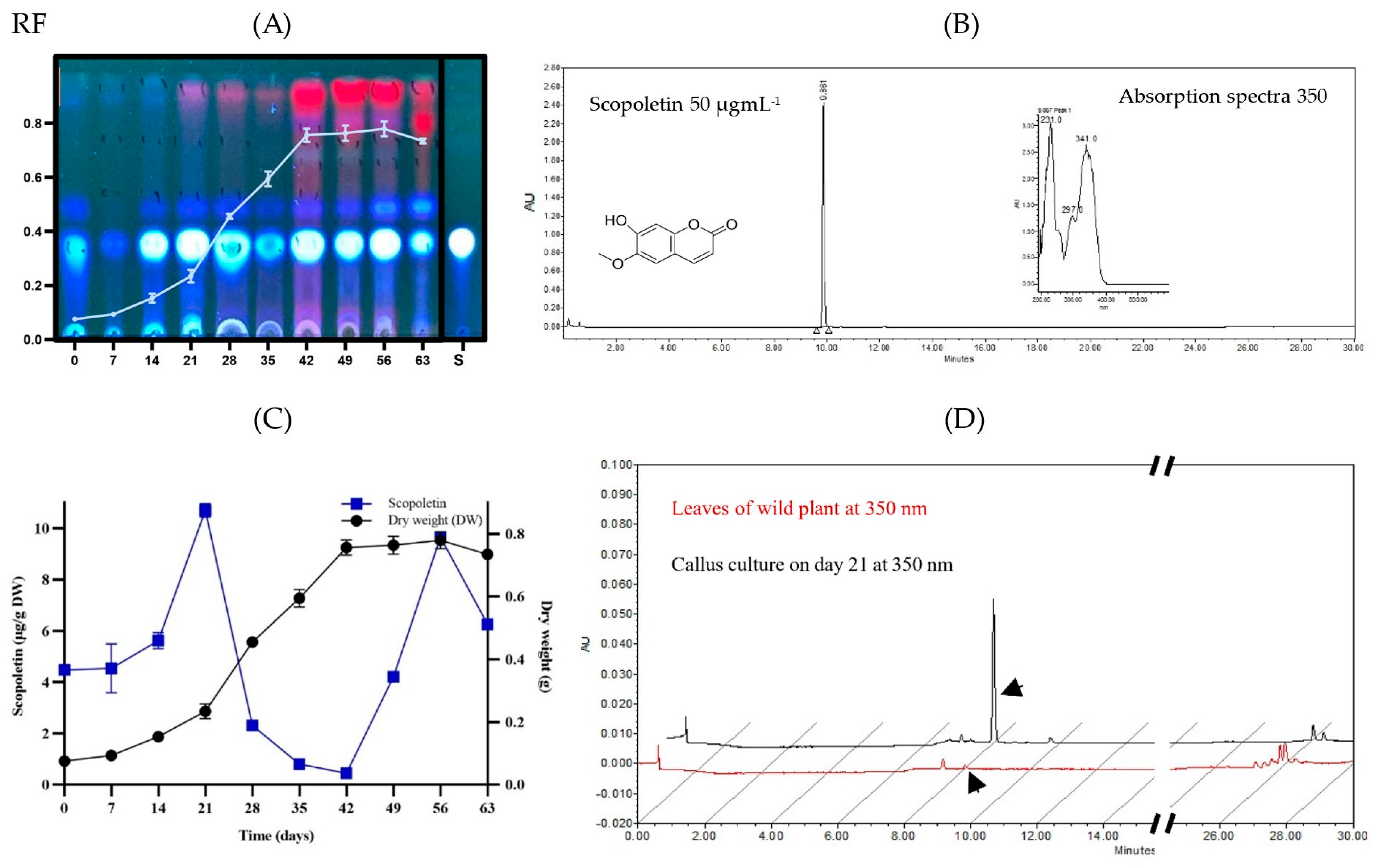
2.4. Chemical Profile of B. fagaroides Callus Culture
2.5. Chemical Profile of Wild B. fagaroides Leaves
2.6. Evaluation of Cytotoxic Activity
3. Discussion
4. Materials and Methods
4.1. General
4.2. Identification of Plant Material
4.3. Obtaining Plant Material
4.4. Callus Induction
4.5. Growth Kinetics of Callus Culture
4.6. Cell Morphology and Viability
4.7. Phytochemical Analysis of the Callus Culture
4.7.1. Obtaining the Extracts
4.7.2. Chemical Profile and Identification of Major Compounds
4.8. Phytochemical Analysis of Wild Plant Leaves
4.8.1. Obtaining the Extracts
4.8.2. Chemical Profile and Identification of Major Compounds
4.9. Cytotoxicity Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noge, K.; Becerra, J.X. Germacrene D, a common sesquiterpene in the genus Bursera (Burseraceae). Molecules 2009, 14, 5289–5297. [Google Scholar] [CrossRef] [PubMed]
- Rzedowski, J.; Medina, L.R.; Calderón, D.R.G. Las especies de Bursera (Burseraceae) en la cuenca superior del río Papaloapan (México). Acta Bot. Mex. 2004, 66, 23–151. [Google Scholar] [CrossRef]
- Rzedowski, J.; Medina, L.R.; Calderón, D.R.G. Inventario del conocimiento taxonómico, así como de la diversidad y del endemismo regionales de las especies mexicanas de Bursera (Burseraceae). Acta Bot. Mex. 2005, 70, 85–111. [Google Scholar] [CrossRef]
- Marcotullio, M.C.; Curini, M.; Becerra, J.X. An Ethnopharmacological, Phytochemical and Pharmacological Review on Lignans from Mexican Bursera spp. Molecules 2018, 23, 1976. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Sepúlveda, A.M.; Mendieta-Serrano, M.; Antúnez, M.M.Y.; Salas-Vidal, E.; Marquina, S.; Villarreal, M.L.; Puebla, A.M.; Delgado, J.; Alvarez, L. Cytotoxic podophyllotoxin type-lignans from the steam bark of Bursera fagaroides var. fagaroides. Molecules 2012, 17, 9506–9519. [Google Scholar] [CrossRef] [PubMed]
- Antúnez, M.M.; León, A.; Rojas-Sepúlveda, A.M.; Marquina, S.; Mendieta-Serrano, M.A.; Salas-Vidal, E.; Villarreal, M.L.; Alvarez, L. Aryldihydronaphthalene-type lignans from Bursera fagaroides var. fagaroides and their antimitotic mechanism of action. RSC Adv. 2016, 6, 4950–4959. [Google Scholar] [CrossRef]
- Antúnez-Mojica, M.; Romero-Estrada, A.; Hurtado-Díaz, I.; Miranda-Molina, A.; Alvarez, L. Lignans from Bursera fagaroides: Chemistry, Pharmacological Effects and Molecular Mechanism. A Current Review. Life 2021, 11, 685. [Google Scholar] [CrossRef]
- Bianchi, E.; Sheth, K.; Cole, J.R. Antitumor agents from Bursera fagaroides (Burseraceae). (β-peltatin-a-methylether and 5′-desmethoxy-β-peltatin-a-methylether). Tetrahedron Lett. 1969, 10, 2759–2762. [Google Scholar] [CrossRef]
- Velázquez-Jiménez, R.; Torres-Valencia, J.M.; Cerda-García-Rojas, C.M.; Hernández-Hernández, J.D.; Román-Marín, L.U.; Manríquez-Torres, J.J.; Gómez-Hurtado, M.A.; Valdez-Calderón, A.; Motilva, V.; García-Mauriño, S.; et al. Absolute configuration of podophyllotoxin related lignans from Bursera fagaroides using vibrational circular dichroism. Phytochemistry 2011, 72, 2237–2243. [Google Scholar] [CrossRef]
- Acevedo, M.; Nuñez, P.; González-Maya, L.; Cardoso, T.A.; Villarreal, M.L. Cytotoxic and Anti-inflamatory activities of Bursera species from México. J. Clin. Toxicol. 2015, 5, 232. [Google Scholar] [CrossRef]
- Antúnez-Mojica, M.; Rojas-Sepúlveda, A.M.; Mendieta-Serrano, M.A.; Gonzalez-Maya, L.; Marquina, S.; Salas-Vidal, E.; Alvarez, L. Lignans from Bursera fagaroides affect in vivo cell behavior by disturbing the tubulin cytoskeleton in zebrafish embryos. Molecules 2019, 24, 8. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Ordoñez, M.F.; Del Coro Arizmendi, M.; Flores-Enríquez, V.; Márquez-Guzmán, J. Extracción y morfología del embrión de Bursera bipinnata (Burseraceae). Acta Bot. Mex. 2016, 117, 27–35. [Google Scholar] [CrossRef]
- García-Aguilar, J.Á.; Luna-Cedeño, J.J.; Rodríguez-Ortiz, G.; Leyva-López, J.C. Acumulación de biomasa en estacas de cuatro especies de Bursera en Oaxaca. CES 2021, 28, 1–11. [Google Scholar] [CrossRef]
- Ceballos, G.; Martínez, L.; García, A.; Espinoza, E.; Bezaury, C.J.; Dirzo, R. Diversidad, Amenazas y Áreas Prioritarias para la Conservación de las Selvas Secas del Pacífico de México, 1st ed.; Fondo de Cultura Económica, CONABIO: Mexico City, Mexico, 2010; pp. 9–11. [Google Scholar]
- Bonfil-Sanders, C.; Mendoza-Hernández, P.E.; Ulloa-Nieto, J.A. Root and callus development in cuttings of seven species of the genus Bursera. Agrociencia 2007, 41, 103–109. Available online: https://www.researchgate.net/publication/28138673 (accessed on 30 May 2024).
- Satyanarayana, B.N.; Varghese, D.B. Plant Tissue Culture: Practices and New Experimental Protocols; I. K. International Pub. House. Pvt. Ltd.: New Delhi, India, 2007; pp. 9–54. ISBN 978-81-89866-11-2. [Google Scholar]
- Verpoorte, R.; Alfermann, A.W. Metabolic Engineering of Plant Secondary Metabolism; Kluger Academic Publishers, Springer: Dutch, The Netherlands, 2013; pp. 1–46. ISBN 978-94-015-9423-3. [Google Scholar]
- Rodríguez, B.M.M.; Latsague, V.M.I.; Chacón, F.M.A.; Astorga, B.P.K. In vitro induction of callogenesis and indirect organogenesis from explants of cotyledon, hypocotyl and leaf in Ugni molinae. Bosque 2014, 35, 21–22. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Sugimoto, K.; Iwase, A. Plant Callus: Mechanisms of Induction and Repression. Plant Cell 2013, 25, 3159–3173. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.R.; Reyes, J.; Loyola, V.M. Biosíntes y Bioconversión de Metabolitos Secundarios por Células Cultivadas In Vitro; Cultivo de Tejidos en la Agricultura: Cali, Colombia, 1991; 9th Chapter; pp. 212–231. [Google Scholar]
- Abdulhafiz, F.; Mohammed, A.; Reduan, M.F.H.; Kari, Z.A.; Wei, L.S.; Goh, K.W. Plant cell culture technologies: A promising alternatives to produce high-value secondary metabolites. Arab. J. Chem. 2022, 15, 104161. [Google Scholar] [CrossRef]
- Motolinía-Alcántara, E.A.; Castillo-Araiza, C.O.; Rodríguez-Monroy, M.; Román-Guerrero, A.; Cruz-Sosa, F. Engineering Considerations to Produce Bioactive Compounds from Plant Cell Suspension Culture in Bioreactors. Plants 2021, 10, 2762. [Google Scholar] [CrossRef]
- Shishodia, S.; Azu, N.; Rosenzweig, J.A.; Jackson, D.A. Guggulsterone for Chemoprevention of Cancer. Curr. Pharm. Des. 2016, 22, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Kumar, A. Biosynthesis of guggulsterone in the callus culture of Commiphora wightii Arnott. bhandari (Burseraceae). Res. J. Pharm. Biol. Chem. Sci. 2010, 1, 35–41. [Google Scholar]
- Al-Abdallat, A.M.; Adayileh, B.K.; Sawwan, J.S.; Shibli, R.; Al-Qudah, T.S.; Abu-Irmaileh, B.; Albdaiwi, R.N.; Almaliti, J.; Bustanji, Y. Secondary Metabolites Profiling, Antimicrobial and Cytotoxic Properties of Commiphora gileadensis L. Leaves, Seeds, Callus, and Cell Suspension Extracts. Metabolites 2023, 13, 537. [Google Scholar] [CrossRef] [PubMed]
- Nikam, T.D.; Ghorpade, R.P.; Nitnaware, K.M.; Ahire, M.L.; Lokhande, V.H.; Chopra, A. Micropropagation and non-steroidal anti-inflammatory and anti-arthritic agent boswellic acid production in callus cultures of Boswellia serrata Roxb. Physiol. Mol. Biol. Plants 2013, 19, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Ozyigit, I.I.; Dogan, I.; Hocaoglu-Ozyigit, A.; Yalcin, B.; Erdogan, A.; Yalcin, I.E.; Cabi, E.; Kaya, Y. Production of secondary metabolites using tissue culture-based biotechnological applications. Front. Plant Sci. 2023, 14, 1132555. [Google Scholar] [CrossRef] [PubMed]
- Trejo-Tapia, G.; Rodríguez-Monroy, M. La agregación celular en la producción de metabolitos secundarios en cultivos vegetales in vitro. Interciencia 2007, 32, 669–674. Available online: http://www.redalyc.org/articulo.oa?id=33901004 (accessed on 30 May 2024).
- Larson, C.G.; Gómez, C.; Sánchez-Olate, M.; Ríos, D. Induction of indirect callogenesis in Eucalyptus globulus. Bosque 2006, 27, 250–257. [Google Scholar] [CrossRef]
- Feeney, M.; Bhagwat, B.; Mitchell, J.S.; Lane, W.D. Shoot regeneration from organogenic callus of sweet cherry (Prunus avium L.). Plant Cell Tissue Organ Cult. 2007, 90, 201–214. [Google Scholar] [CrossRef]
- Rashmi, R.; Trivedi, M.P. Effect of Various Growth Hormone Concentration and Combination on Callus Induction, Nature of Callus and Callogenic Response of Nerium odorum. Appl. Biochem. Biotechnol. 2014, 172, 2562–2570. [Google Scholar] [CrossRef] [PubMed]
- Bonfil-Sanders, C.; Cajero-Lázaro, I.; Evans, R.Y. Seed germination of six Bursera species from central México. Agrociencia 2008, 42, 827–834. [Google Scholar]
- Cultid-Medina, C.A.; Rico, Y. Los aliados emplumados de los Copales y Cuajiotes de México: Aves y la dispersión de semillas de Bursera. Rev. Digit. Univ. 2020, 21, 1–9. [Google Scholar] [CrossRef]
- Rzedowski, J.; Guevara, F.F. Familia Burceraceae, Flora del Bajío y de Regiones Adyacentes, 2nd ed.; Rzedowski, J., Calderón, G., Eds.; Instituto de Ecología A.C., Departamento de Publicaciones Apdo: Xalapa, Veracruz, 1992; Fascículo 3; pp. 14–20. [Google Scholar]
- Mc-Caughey-Espinoza, D.; Ayala-Astorga, G.; García-Baldenegro, C.; Buitimea-Cantúa, N.; Buitimea-Cantúa, G.; Ochoa-Meza, A. In vitro germination and induction of callus and root in Bursera laxiflora S. Watson. Abanico Agrofor. 2020, 2, 1–14. [Google Scholar]
- Pavón-Reyes, L.; Evangelista-Lozano, S.; Sepúlveda-Jiménez, G.; Chávez, A.V.; Rodríguez-Monroy, M. Cell Culture of Bursera linanoe in a Stirred Tank Bioreactor for Production of Linalool and Linalyl Acetate. Nat. Prod. Commun. 2017, 12, 319–322. [Google Scholar] [PubMed]
- Malik, S.I.; Rashid, H.; Yasmin, T.; Minhas, N.M. Effect of 2,4-dichlorophenoxyacetic Acid on Callus Induction from Mature Wheat (Triticum aestivum L.) Seeds. Int. J. Agric. Biol. 2003, 6, 156–159. Available online: https://www.researchgate.net/publication/242453014 (accessed on 30 May 2024).
- Zheng, M.Y.; Konzak, C.F. Effect of 2,4-dichlorophenoxyacetic acid on callus induction and plant regeneration in anther culture of wheat (Triticum aestivum L.). Plant Cell Rep. 1999, 19, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Carsono, N.; Juwendah, E.; Liberty, L.; Sari, S.; Damayanti, F.; Rachmadi, M. Optimize 2,4-D concentration and callus induction time enhance callus proliferation and plant regeneration of three rice genotypes. Biodiversitas 2021, 22, 2555–2560. [Google Scholar] [CrossRef]
- Ozias-Akins, P.; Vasil, I.K. Callus induction and growth from the mature embryo of Triticum aestivum (Wheat). Protoplasma 1983, 115, 104–113. [Google Scholar] [CrossRef]
- Pádua, M.S.; Paiva, L.V.; Silva, L.C.D.; Livramento, K.G.D.; Alves, E.; Castro, A.H.F. Morphological characteristics and cell viability of coffee plants calli. Cienc. Rural 2014, 44, 660–665. [Google Scholar] [CrossRef]
- Taguchi, G.; Fujikawa, S.; Yazawa, T.; Kodaira, R.; Hayashida, N.; Shimosaka, M.; Okazaki, M. Scopoletin uptake from culture medium and accumulation in the vacuoles after conversion to scopolin in 2,4-D-treated tobacco cells. Plant Sci. 2000, 151, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, M.; Hino, F.; Kominami, K.; Miura, Y. Effects of Plant Hormones on Formation of Scopoletin and Scopolin in Tobacco Tissue Cultures. Agric. Biol. Chem. 1982, 46, 609–614. [Google Scholar] [CrossRef]
- Cisneros-Torres, D.; Cruz-Sosa, F.; González-Cortazar, M.; Martínez-Trujillo, A.; Nicasio-Torres, P. Enhancing the production of scopoletin and quercetin 3-O-β-d-glucoside from cell suspension cultures of Tilia americana var. mexicana by modulating the copper and nitrate concentrations. Plant Cell Tissue Organ Cult. 2019, 139, 305–316. [Google Scholar] [CrossRef]
- Siatka, T.; Chlebekb, J.; Hoštálková, A. Copper (II) Sulfate Stimulates Scopoletin Production in Cell Suspension Cultures of Angelica archangelica. Nat. Prod. Commun. 2017, 12, 1779–1780. [Google Scholar] [CrossRef]
- Gnonlonfin, G.J.B.; Sanni, A.; Brimer, L. Review Scopoletin—A Coumarin Phytoalexin with Medicinal Properties. Crit. Rev. Plant Sci. 2012, 31, 47–56. [Google Scholar] [CrossRef]
- Bagni, N.; Fracassini, D.S. The effect of coumarin derivatives on organogenesis and callus growth of Cichorium Intybus roots and Helianthus tuberosus tubers in vitro. Experientia 1971, 27, 1239–1241. [Google Scholar] [CrossRef]
- Ekiert, H.; Gomółka, E. Coumarin compounds in Ammi majus L. callus cultures. Die Pharm. 2000, 55, 684–687. [Google Scholar]
- Fritig, B.; Hirth, L.; Ourisson, G. Biosynthesis of the coumarins: Scopoletin formation in tobacco tissue cultures. Phytochemistry 1970, 9, 1963–1975. [Google Scholar] [CrossRef]
- Modafar, C.E.; Clerivet, A.; Fleuriet, A.; Macheix, J.J. Inoculation of Platanus acerifolia with Ceratocystis fimbriata F. Sp. Platani induces scopoletin and umbelliferone accumulation. Phytochemistry 1993, 34, 1271–1276. [Google Scholar]
- Habibullah, M.; Al-Mansur, M.A.; Mahboob, M.; Siddiqi, A.; Chowdhury, A.M.S.; Sohrab, M.; Hasan, C.M. Scopoletin Isolated from Stem Bark of Bursera serrata Wall. With Antimicrobial and Cytotoxic Activities of the Crude Extract. Dhaka Univ. J. Sci. 2010, 58, 287–289. [Google Scholar]
- Ara, K.; Rahman, M.S.; Rahman, A.H.M.; Hasan, C.M.; Rashid, M.A. Terpenoids and Coumarin from Bursera serrata Wall. Dhaka Univ. J. Pharm. Sci. 2009, 8, 107–110. [Google Scholar] [CrossRef]
- Aguilar, S.L.; Romero, C.O.; González, C.M.; Tortoriello, J. Efecto de Bursera grandiflora sobre el peso corporal y lipemia en ratones obesos. Boletín Latinoam. Caribe Plantas Med. Aromáticas 2012, 11, 138–146. [Google Scholar]
- Matos, M.J.; Santana, L.; Uriarte, E.; Abreu, O.A.; Molina, E.; Yordi, E.G. Coumarins—An Important Class of Phytochemicals. Phytochemicals—Isolation, Characterization. In Human Health; Rao, A.V., Rao, L.G., Eds.; InTech: London, UK, 2015; Volume 20, pp. 113–140. [Google Scholar] [CrossRef]
- Rojas-Sepúlveda, A.M. Búsqueda de Metabolitos con Actividad Citotóxica y Antitumoral en Bursera fagaroides var. fagaroides y Bursera morelensis, y Evaluación de su Efecto como Inhibidores del Ciclo Celular en el Modelo de pez Cebra. Ph.D. Thesis, Universidad Autónoma del Estado de Morelos, Cuernavaca, Mexico, June 2012. [Google Scholar]
- Infante-Rodríguez, D.A.; Landa-Cansigno, C.; Gutiérrez-Sánchez, A.; Murrieta-León, D.L.; Reyes-López, C.; Castillejos-Pérez, A.B.; Pucheta-Fiscal, J.E.; Velázquez-Narváez, A.C.; Monribot-Villanueva, J.L.; Guerrero-Analco, J.A. Análisis fitoquímico y actividad antidiabética, antibacteriana y antifúngica de hojas de Bursera simaruba (Burseraceae). Acta Bot. Mex. 2022, 129, 1–15. [Google Scholar] [CrossRef]
- Tripathi, N.; Kumar, S.; Singh, R.; Singh, C.J.; Singh, P.; Varshney, V.K. Isolation and Identification of γ- Sitosterol by GC-MS from the Leaves of Girardinia heterophylla (Decne). Open Bioact. Compd. J. 2013, 4, 25–27. [Google Scholar] [CrossRef]
- Shahzad, N.; Khan, W.; Md, S.; Ali, A.; Saluja, S.S.; Sharma, S. Phytosterols as a natural anticancer agent: Current status and future perspective. Biomed. Pharmacother. 2017, 88, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, P.D.; Pollier, J.; Panda, S.; Szymanski, J.; Massalha, H.; Yona, M.; Unger, T.; Malitsky, S.; Arendt, P.; Pauwels, L.; et al. Plant cholesterol biosynthetic pathway overlaps with phytosterol metabolism. Nat. Plants 2016, 3, 16205. [Google Scholar] [CrossRef]
- Rubis, B.; Paszel, A.; Kaczmarek, M.; Rudzinska, M.; Jelen, H.; Rybczynska, M. Beneficial or harmful influence of phytosterols on human cells? Br. J. Nutr. 2008, 100, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Raicht, R.F.; Cohen, B.I.; Fazzini, E.P.; Sarwal, A.N.; Takahashi, M. Protective effect of plant sterols against chemically induced colon tumors in rats. Cancer Res. 1980, 40, 403–405. [Google Scholar]
- Tian, Q.; Wang, L.; Sun, X.; Zeng, F.; Pan, Q.; Xue, M. Scopoletin exerts anticancer effects on human cervical cancer cell lines by triggering apoptosis, cell cycle arrest, inhibition of cell invasion and PI3K/AKT signalling pathway. JBUON 2019, 24, 997–1002. [Google Scholar]
- Barry, L.N. Use of Fluorescein Diacetate in Citrus Tissue cultures for the determination of cell viability and the selection of mutants. Sci. Hortic. 1989, 39, 15–21. [Google Scholar]
- Jones, K.H.; Senft, J.A. An improved method to determine cell viability by viability by simultaneous staining with fluorescein diacetate-propidium Iodide. J. Histochem. Cytochem. 1985, 33, 77–79. [Google Scholar] [CrossRef]
- Zhu, L.; Li, M.; Yang, W.; Zhang, J.; Yang, X.; Zhang, Q.; Wang, Q. Effects of Different Drying Methods on Drying Characteristics and Quality of Glycyrrhiza uralensis (Licorice). Foods 2023, 12, 1652. [Google Scholar] [CrossRef]
- Bork, P.M.; Schmitz, M.L.; Kuhnt, M.; Escher, C.; Heinrich, M. Sesquiterpene lactone containing Mexican Indian medicinal plants and pure sesquiterpene lactones as potent inhibitors of transcription factor NF-kappaB. FEBS Lett. 1997, 402, 85–90. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MEDIA | PGR (mgL−1) | CALLUS (%) ± DE | |
|---|---|---|---|
| 2,4-D | KINETIN | ||
| MS | 2 | 0 | 0.00 ± 0.00 d |
| MS | 2 | 1 | 0.00 ± 0.00 d |
| MS | 2 | 2 | 86.67 ± 5.77 a |
| MS | 3 | 1 | 0.00 ± 0.00 d |
| MS | 3 | 2 | 0.00 ± 0.00 d |
| MS | 3 | 3 | 0.00 ± 0.00 d |
| MS | 4 | 1 | 0.00 ± 0.00 d |
| MS | 4 | 2 | 33.33 ± 5.77 b |
| MS | 4 | 3 | 0.00 ± 0.00 d |
| MS | 4 | 4 | 26.67 ± 5.77 b |
| B5 | 2 | 0 | 0.00 ± 0.00 d |
| B5 | 2 | 1 | 13.33 ± 5.77 c |
| B5 | 2 | 2 | 16.67 ± 5.77 c |
| B5 | 3 | 1 | 0.00 ± 0.00 d |
| B5 | 3 | 2 | 0.00 ± 0.00 d |
| B5 | 3 | 3 | 0.00 ± 0.00 d |
| B5 | 4 | 1 | 0.00 ± 0.00 d |
| B5 | 4 | 2 | 0.00 ± 0.00 d |
| B5 | 4 | 3 | 0.00 ± 0.00 d |
| B5 | 4 | 4 | 0.00 ± 0.00 d |
| Extract | HepG2 | HeLa | PC3 | H1299 | HFF-1 |
|---|---|---|---|---|---|
| Leaves b | 20.3 ± 3.38 **** | 18.6 ± 5.2 *** | 12.6 ± 4.6 **** | 69.1 ± 6.2 **** | 79.6 ± 5.2 * |
| Callus b | 155.4 ± 4.95 **** | 72 ± 5 *** | 141 ± 5.8 **** | 210.4 ± 5 **** | 95.4 ± 6.2 * |
| Paclitaxel a | 10.12 ± 2.5 | 40.23 ± 7.2 | 17.5 ± 2.8 | 45.25 ± 7 | 989 ± 21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Mejía, N.; Villarreal, M.L.; Sánchez-Carranza, J.N.; González-Maya, L.; González-Cortazar, M.; Ortíz-Caltempa, A.; Alvarez, L. Phytochemical Profiles and Cytotoxic Activity of Bursera fagaroides (Kunth) Engl. Leaves and Its Callus Culture. Plants 2024, 13, 1622. https://doi.org/10.3390/plants13121622
Pérez-Mejía N, Villarreal ML, Sánchez-Carranza JN, González-Maya L, González-Cortazar M, Ortíz-Caltempa A, Alvarez L. Phytochemical Profiles and Cytotoxic Activity of Bursera fagaroides (Kunth) Engl. Leaves and Its Callus Culture. Plants. 2024; 13(12):1622. https://doi.org/10.3390/plants13121622
Chicago/Turabian StylePérez-Mejía, Nancy, María Luisa Villarreal, Jessica Nayelli Sánchez-Carranza, Leticia González-Maya, Manasés González-Cortazar, Anabel Ortíz-Caltempa, and Laura Alvarez. 2024. "Phytochemical Profiles and Cytotoxic Activity of Bursera fagaroides (Kunth) Engl. Leaves and Its Callus Culture" Plants 13, no. 12: 1622. https://doi.org/10.3390/plants13121622