

Fertilizer Effects on the Nitrogen Isotope Composition of Soil and Different Leaf Locations of Potted Camellia sinensis over a Growing Season


 , ,
, ,  and
and
Abstract

1. Introduction
2. Materials and Methods
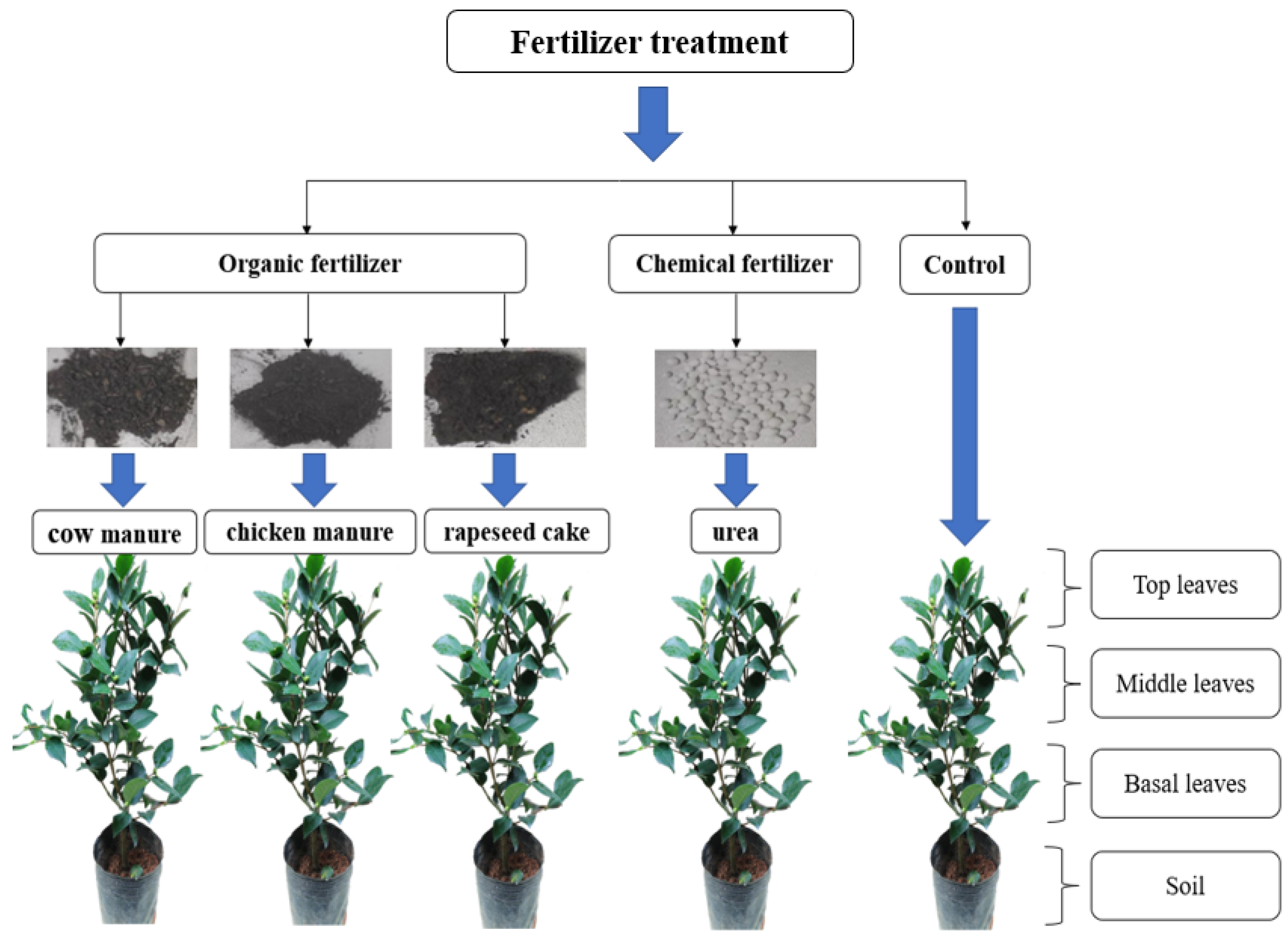
2.1. Pot Experiments and Sampling
2.2. Stable Isotope Analysis
2.3. Statistical Analysis
3. Results and Discussion
3.1. Fertilizer Effect on Nitrogen Content and δ15N Values of Top Tea Leaves and Soil
3.2. Temporal δ15N Variations in Tea Leaves Treated with Different Fertilizers
3.3. δ15 N Differences between the Top, Middle, and Basal Tea Leaves
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pan, S.; Nie, Q.; Tai, H.; Song, X.; Tong, X.; Zhang, L.; Wu, X.; Lin, Z.; Zhang, Y.; Ye, D.; et al. Tea and tea drinking: China’s outstanding contributions to the mankind. Chin. Med. 2022, 17, 1–40. [Google Scholar] [CrossRef]
- Engelhardt, U.H. Tea chemistry—What do and what don’t we know?—A micro review. Food Res. Int. 2020, 132, 109120. [Google Scholar] [CrossRef]
- Zhao, C.; Tang, G.; Cao, S.; Xu, X.; Gan, R.; Liu, Q.; Mao, Q.; Shang, A.; Li, H. Phenolic profiles and antioxidant activities of 30 tea infusions from green, black, oolong, white, yellow and dark teas. Antioxidants 2019, 8, 215. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, X.; Ho, C.; Huang, Q. Chemistry and health effect of tea polyphenol (−)-epigallocatechin 3-O-(3-O-methyl) gallate. J. Agric. Food Chem. 2018, 67, 5374–5378. [Google Scholar] [CrossRef]
- Zhou, J.; Ding, L.; Chen, W.; Wang, Y. Green tea catechin epigallocatechin gallate alleviates high-fat diet-induced obesity in mice by regulating the gut–brain axis. Food Front. 2023, 4, 1413–1425. [Google Scholar] [CrossRef]
- Giampieri, F.; Mazzoni, L.; Cianciosi, D.; Alvarez-Suarez, J.M.; Regolo, L.; Sánchez-González, C.; Capocasa, F.; Xiao, J.; Mezzetti, B.; Battino, B. Organic vs. conventional plant-based foods: A review. Food Chem. 2022, 383, 132352. [Google Scholar] [CrossRef]
- Hoefkens, C.; Vandekinderen, I.; Meulenaer, D.B.; Devlieghere, F.; Baert, K.; Sioen, I.; Henauw, S.D.; Verbeke, W.; Camp, J.V. A literature-based comparison of nutrient and contaminant contents between organic and conventional vegetables and potatoes. Br. Food J. 2009, 111, 1078–1097. [Google Scholar] [CrossRef]
- Muñoz-Redondo, J.M.; Montenegro, J.C.; Moreno-Rojas, J.M. Using Nitrogen Stable Isotopes to Authenticate Organically and Conventionally Grown Vegetables: A New Tracking Framework. Agronomy 2022, 13, 131. [Google Scholar] [CrossRef]
- Li, C.; Kang, X.; Nie, J.; Li, A.; Farag, M.A.; Liu, C.; Rogers, K.M.; Xiao, J.; Yuan, Y. Recent advances in Chinese food authentication and origin verification using isotope ratio mass spectrometry. Food Chem. 2023, 398, 133896. [Google Scholar] [CrossRef]
- Bateman, A.S.; Kelly, S.D.; Woolfe, M. Nitrogen isotope composition of organically and conventionally grown crops. J. Agric. Food Chem. 2007, 55, 2664–2670. [Google Scholar] [CrossRef]
- Wassenaar, L.I.; Kelly, S.D.; Douence, C.; Islam, M.; Monteiro, L.; Abrahim, A.; Rinke, P. Assessment of rapid low-cost isotope (δ15N, δ18O) analyses of nitrate in fruit extracts by Ti (III) reduction to differentiate organic from conventional production. Rapid Commun. Mass Spectrom. 2022, 36, e9259. [Google Scholar] [CrossRef]
- Rogers, K.M.; Turnbull, R.E.; Martin, A.P.; Baisden, W.T.; Rattenbury, M.S. Stable isotopes reveal human influences on southern New Zealand soils. Appl. Geochem. 2017, 82, 15–24. [Google Scholar] [CrossRef]
- Busari, M.A.; Salako, F.K.; Tuniz, C. Stable isotope technique in the evaluation of tillage and fertilizer effects on soil carbon and nitrogen sequestration and water use efficiency. Eur. J. Agron. 2016, 73, 98–106. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, W.; Zhang, Y.; Liu, Z.; Shao, S.; Zhou, L.; Rogers, K.M. Differentiating organically farmed rice from conventional and green rice harvested from an experimental field trial using stable isotopes and multi-element chemometrics. J. Agric. Food Chem. 2018, 66, 2607–2615. [Google Scholar] [CrossRef]
- Lyu, C.; Yang, J.; Wang, T.; Kang, C.; Wang, S.; Wang, H.; Wan, X.; Zhou, L.; Zhang, W.; Huang, L.; et al. A field trials-based authentication study of conventionally and organically grown Chinese yams using light stable isotopes and multi-elemental analysis combined with machine learning algorithms. Food Chem. 2021, 33, 128506. [Google Scholar] [CrossRef]
- Liu, X.; Liu, Z.; Qian, Q.; Song, W.; Rogers, K.M.; Rao, Q.; Wang, S.; Zhang, Q.; Shao, S.; Tian, M.; et al. Isotope chemometrics determines farming methods and geographical origin of vegetables from Yangtze River Delta Region, China. Food Chem. 2021, 342, 128379. [Google Scholar] [CrossRef]
- Choi, W.J.; Kwak, J.H.; Lim, S.S.; Park, H.; Chang, S.; Lee, S.; Arshad, M.; Yun, S.; Kim, H. Synthetic fertilizer and livestock manure differently affect δ15N in the agricultural landscape: A review. Agric. Ecosyst. Environ. 2017, 237, 1–15. [Google Scholar] [CrossRef]
- Shin, W.J.; Ryu, J.S.; Mayer, B.; Lee, K.S.; Kim, I. Nitrogen, Sulfur, and Oxygen Isotope Ratios of Animal- and Plant-Based Organic Fertilizers Used in South Korea. J. Environ. Qual. 2017, 237, 559–567. [Google Scholar] [CrossRef]
- Camin, F.; Boner, M.; Bontempo, L.; Fauhl-Hassek, C.; Kelly, S.D.; Riedl, J.; Rossmann, A. Stable isotope techniques for verifying the declared geographical origin of food in legal cases. Trends Food Sci. Technol. 2017, 61, 176–187. [Google Scholar] [CrossRef]
- Fabroni, S.; Bontempo, L.; Campanelli, G.; Canali, S.; Montemurro, F. Innovative Tools for the Nitrogen Fertilization Traceability of Organic Farming Products. Horticulturae 2023, 9, 723. [Google Scholar] [CrossRef]
- Bateman, A.S.; Kelly, S.D. Fertilizer nitrogen isotope signatures. Isot. Environ. 2007, 43, 237–247. [Google Scholar] [CrossRef]
- Hayashi, N.; Ujihara, T.; Tanaka, E.; Kishi, Y.; Ogawa, H.; Matsuo, H. Annual variation of natural 15N abundance in tea leaves and its practicality as an organic tea indicator. J. Agric. Food Chem. 2011, 59, 10317–10321. [Google Scholar] [CrossRef]
- Trapp, T.; Inácio, C.T.; Ciotta, M.N.; Hindersmann, J.; Lima, A.P.; Santos, T.S.; Ferreira, G.W.; Morais, G.P.; Conti, L.; Comin, J.J.; et al. Natural abundance analysis of the role played by 15N as indicator for the certification of organic-system deriving food. J. Sci. Food Agric. 2022, 102, 330–340. [Google Scholar] [CrossRef]
- del Amor, F.M.; Navarro, J.; Aparicio, P.M. Isotopic discrimination as a tool for organic farming certification in sweet pepper. J. Environ. 2008, 37, 182–185. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Y.; Zhang, Y.; Yang, G.; Shao, S.; Nie, J.; Yuan, Y.; Rogers, K.M. Influence of leaf age, species and soil depth on the authenticity and geographical origin assignment of green tea. Rapid Commun. Mass Spectrom. 2019, 33, 625–634. [Google Scholar] [CrossRef]
- Xia, W.; Li, Z.; Yu, C.; Liu, Z.; Nie, J.; Li, C.; Shao, S.; Zhang, Y.; Rogers, K.M.; Yuan, Y. Understanding processing, maturity and harvest period effects to authenticate early-spring Longjing tea using stable isotopes and chemometric analyses. Food Control 2021, 124, 107907. [Google Scholar] [CrossRef]
- Xie, S.; Yang, F.; Feng, H.; Yu, Z.; Liu, C.; Wei, C.; Liang, T. Organic fertilizer reduced carbon and nitrogen in runoff and buffered soil acidification in tea plantations: Evidence in nutrient contents and isotope fractionations. Sci. Total Environ. 2021, 762, 143059. [Google Scholar] [CrossRef]
- Wang, Y.; Niu, G.; Wang, R.; Rousk, K.; Li, A.; Hasi, M.; Wang, C.; Xue, J.; Yang, G.; Lv, X.; et al. Enhanced foliar 15N enrichment with increasing nitrogen addition rates: Role of plant species and nitrogen compounds. Glob. Change Biol. 2023, 29, 1591–1605. [Google Scholar] [CrossRef]
- Coplen, T.B. Guidelines and recommended terms for expression of stable-isotope-ratio and gas-ratio measurement results. Rapid Commun. Mass Spectrom. 2011, 25, 2538–2560. [Google Scholar] [CrossRef]
- Magdas, D.A.; Dehelean, A.; Feher, I.; Radu, S. Isotopic and multielemental fingerprinting of organically and conventionally grown potatoes. Isot. Environ. Health Stud. 2017, 53, 610–619. [Google Scholar] [CrossRef]
- Chi, H.Y.; Kim, W.R.; Kim, J.Y.; Kim, S.H. Improved organic and pesticide-free rice (Oryza sativa L.) authentication based on multiple stable isotope ratio analysis and rice milling state. Heliyon 2024, 10, E26725. [Google Scholar] [CrossRef]
- Ye, J.; Wang, Y.; Kang, J.; Chen, Y.; Hong, L.; Li, M.; Jia, Y.; Wang, Y.; Jia, X.; Wu, Z.; et al. Effects of long-term use of organic fertilizer with different dosages on soil improvement, nitrogen transformation, tea yield and quality in acidified tea plantations. Plants 2022, 12, 122. [Google Scholar] [CrossRef]
- Lee, J.; Jo, N.Y.; Shim, S.Y.; Linh, L.T.Y.; Kim, S.; Lee, M.; Hwang, S. Effects of Hanwoo (Korean cattle) manure as organic fertilizer on plant growth, feed quality, and soil bacterial community. Front. Plant Sci. 2023, 14, 1135947. [Google Scholar] [CrossRef]
- Kubar, M.S.; Zhang, Q.; Feng, M.; Wang, C.; Yang, W.; Kubar, K.A.; Riaz, S.; Gul, H.; Samoon, H.Z.; Sun, H.; et al. Growth, Yield and Photosynthetic Performance of Winter Wheat as Affected by Co-Application of Nitrogen Fertilizer and Organic Manures. Life 2022, 12, 1000. [Google Scholar] [CrossRef]
- Kalcsits, L.A.; Buschhaus, H.A.; Guy, R.D. Nitrogen isotope discrimination as an integrated measure of nitrogen fluxes, assimilation and allocation in plants. Physiol. Plant. 2014, 151, 293–304. [Google Scholar] [CrossRef]
- Trandel, M.A.; Vigardt, A.; Walters, S.A.; Lefticariu, M.; Kinsel, M. Nitrogen isotope composition, nitrogen amount, and fruit yield of tomato plants affected by the soil–fertilizer types. ACS Omega 2018, 3, 6419–6426. [Google Scholar] [CrossRef]
- Bouwman, A.F.; Boumans, L.J.M.; Batjes, N.H. Estimation of global NH3 volatilization loss from synthetic fertilizers and animal manure applied to arable lands and grasslands. Glob. Biogeochem. Cycles 2002, 16, 8-1–8-14. [Google Scholar] [CrossRef]
- Dadrasnia, A.; de Bona Muñoz, I.; Yáñez, E.H.; Lamkaddam, I.U.; Mora, M.; Ponsá, S.; Ahmed, M.; Argelaguet, L.L.; Williams, P.M.; Oatley-Radcliffe, D.L. Sustainable nutrient recovery from animal manure: A review of current best practice technology and the potential for freeze concentration. J. Clean. Prod. 2021, 315, 128106. [Google Scholar] [CrossRef]
- Ruan, L.; Wei, K.; Wang, L.; Cheng, H.; Zhang, F.; Wu, L.; Bai, P.; Zhang, C. Characteristics of NH4+ and NO3− fluxes in tea (Camellia sinensis) roots measured by scanning ion-selective electrode technique. Sci. Rep. 2016, 6, 38370. [Google Scholar] [CrossRef]
- Gurmesa, G.; Hobbie, E.; Zhang, S.; Wang, A.; Zhu, W.; Koba, K.; Yoh, M.; Wang, C.; Zhang, Q.; Fang, Y. Natural 15N abundance of ammonium and nitrate in soil profiles: New insights into forest ecosystem nitrogen saturation. Ecosphere 2022, 13, e3998. [Google Scholar] [CrossRef]
- Takebayashi, Y.; Koba, K.; Sasaki, Y.; Fang, Y.; Yoh, M. The natural abundance of 15N in plant and soil-available N indicates a shift of main plant N resources to NO from NH along the N leaching gradient. Rapid Commun. Mass Spectrom. 2010, 24, 1001–1008. [Google Scholar] [CrossRef]
- Xin, W.; Zhang, J.; Yu, Y.; Tian, Y.; Li, H.; Chen, X.; Li, W.; Liu, Y.; Lu, T.; He, B.; et al. Root microbiota of tea plants regulate nitrogen homeostasis and theanine synthesis to influence tea quality. Curr. Biol. 2024, 34, 868–880.e6. [Google Scholar] [CrossRef]
- Das, P.P.; Singh, K.R.B.; Nagpure, G.; Mansoori, A.; Singh, R.P.; Ghazi, I.A.; Kumar, A.; Singh, J. Plant-soil-microbes: A tripartite interaction for nutrient acquisition and better plant growth for sustainable agricultural practices. Environ. Res. 2022, 214, 113821. [Google Scholar] [CrossRef]
- Chernov, T.I.; Semenov, M.V. Management of soil microbial communities: Opportunities and prospects (a review). Eurasian Soil Sci. 2021, 54, 1888–1902. [Google Scholar] [CrossRef]
- Bai, X.; Wang, Y.; Huo, X.; Salim, R.; Bloch, H.; Zhang, H. Assessing fertilizer use efficiency and its determinants for apple production in China. Ecol. Indic. 2019, 104, 268–278. [Google Scholar] [CrossRef]
- Yan, R.; Wu, H.; Yang, X.; Yang, C.; Lyu, H.; Zhang, H.; Li, S.; Liu, T.; Li, R.; Yao, Y. Soil decreases N2O emission and increases TN content during combined composting of wheat straw and cow manure by inhibiting denitrification. Chem. Eng. J. 2023, 477, 147306. [Google Scholar] [CrossRef]
- Lin, S.; Chen, Z.; Chen, T.; Deng, W.; Wan, X.; Zhang, Z. Theanine metabolism and transport in tea plants (Camellia sinensis L.): Advances and perspectives. Crit. Rev. Biotechnol. 2023, 43, 327–341. [Google Scholar] [CrossRef]
- Xia, W.; Li, C.; Nie, J.; Shao, S.; Rogers, K.M.; Zhang, Y.; Li, Z.; Yuan, Y. Stable isotope and photosynthetic response of tea grown under different temperature and light conditions. Food Chem. 2022, 368, 130771. [Google Scholar] [CrossRef]
- Li, F.; Dong, C.; Yang, T.; Ma, J.; Zhang, S.; Wei, S.; Wan, X.; Zhang, Z. Seasonal theanine accumulation and related gene expression in the roots and leaf buds of tea plants (Camellia sinensis L.). Front. Plant Sci. 2019, 10, 1397. [Google Scholar] [CrossRef]
- Craine, J.M.; Brookshire, E.N.J.; Cramer, M.D.; Hasselquist, N.J.; Koba, K.; Marin-Spiotta, E.; Wang, L. Ecological interpretations of nitrogen isotope ratios of terrestrial plants and soils. Plant Soil 2015, 396, 1–26. [Google Scholar] [CrossRef]
- Zhang, W.; Ni, K.; Long, L.; Ruan, J. Nitrogen transport and assimilation in tea plant (Camellia sinensis): A review. Front. Plant Sci. 2023, 14, 1249202. [Google Scholar] [CrossRef]
- Zhu, B.; Guo, J.; Dong, C.; Li, F.; Qiao, S.; Lin, S.; Yang, T.; Wu, Y.; Bao, S.; Lucas, W.J.; et al. CsAlaDC and CsTSI work coordinately to determine theanine biosynthesis in tea plants (Camellia sinensis L.) and confer high levels of theanine accumulation in a non-tea plant. Plant Biotechnol. 2021, 19, 2395. [Google Scholar] [CrossRef]
- Zhao, J.; Li, P.; Xia, T.; Wan, X. Exploring plant metabolic genomics: Chemical diversity, metabolic complexity in the biosynthesis and transport of specialized metabolites with the tea plant as a model. Crit. Rev. Biotechnol. 2020, 40, 667–688. [Google Scholar] [CrossRef]
- Liu, W.; Cui, S.; Wu, L.; Qi, W.; Chen, J.; Ye, Z.; Ma, J.; Liu, D. Effects of Bio-organic Fertilizer on Soil Fertility, Yield, and Quality of Tea. J. Soil Sci. Plant Nutr. 2023, 23, 5109–5121. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Fertilizer Type | Name | Nitrogen Content (%) | Fertilizer Dose/Pot (g) | Nitrogen Dose/Pot (g) | δ15N Value (‰) |
|---|---|---|---|---|---|
| OF | Chicken manure | 2.3 | 69.6 | 1.6 | 6.5 |
| Rapeseed cake | 7.8 | 20.5 | 1.6 | 1.9 | |
| Cow manure | 1.5 | 106.7 | 1.6 | 6.9 | |
| CF | Urea | 44.8 | 3.6 | 1.6 | −0.6 |
| Treatment | δ15N (‰) Mean ± SD | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 2022 20 October | 2022 18 November | 2023 17 February | 2023 17 March | 2023 21 April | 2023 10 May | 2023 20 May | 2023 30 May | ||
| OF | Chicken manure | 6.5 ± 0.7 a | 6.5 ± 0.7 a | 6.0 ± 0.3 a | 5.4 ± 0.6 a | 6.5 ± 0.3 a | 6.2 ± 0.2 a | 5.1 ± 0.1 a | 4.9 ± 0.1 ab |
| Rapeseed cake | 5.5 ± 0.4 b | 5.3 ± 0.4 b | 5.6 ± 0.7 a | 5.3 ± 0.8 a | 6.4 ± 1.0 a | 6.6 ± 0.3 a | 5.8 ± 0.3 a | 5.2 ± 0.6 a | |
| Cow manure | 3.7 ± 0.4 c | 3.6 ± 0.4 c | 3.5 ± 0.9 b | 3.4 ± 0.6 b | 3.7 ± 0.9 b | 4.1 ± 0.7 b | 3.4 ± 0.6 b | 3.7 ± 1.1 b | |
| CF | Urea | 0.4 ± 0.3 d | 0.6 ± 0.3 d | −0.3 ± 0.4 c | 0.2 ± 0.1 c | −0.1 ± 0.2 c | 0.2 ± 0.7 c | −0.2 ± 0.4 c | −0.9 ± 0.7 c |
| Control | No fertilizer | 2.2 ± 0.6 e | 1.9 ± 0.9 e | 1.5 ± 0.3 d | 1.5 ± 0.8 d | 0.8 ± 0.8 c | 1.1 ± 0.3 d | 1.2 ± 0.2 d | 0.3 ± 0.5 c |
| Treatment | δ15N (‰) Mean ± SD | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 2022 20 October | 2022 18 November | 2023 17 February | 2023 17 March | 2023 21 April | 2023 10 May | 2023 20 May | 2023 30 May | ||
| OF | Chicken manure | 4.6 ± 0.2 a | 4.8 ± 0.2 b | 4.4 ± 0.1 a | 4.4 ± 0.2 b | 4.8 ± 0.2 a | 4.5 ± 0.1 ab | 4.6 ± 0.1 a | 4.5 ± 0.2 b |
| Rapeseed cake | 3.9 ± 0.4 a | 4.0 ± 0.3 c | 3.3 ± 0.2 b | 3.4 ± 0.6 c | 3.4 ± 0.1 b | 3.3 ± 0.4 bc | 3.3 ± 0.3 b | 3.6 ± 0.2 c | |
| Cow manure | 4.9 ± 1.0 a | 6.0 ± 0.3 a | 5.0 ± 0.9 a | 5.4 ± 0.7 a | 5.4 ± 0.4 a | 4.9 ± 1.4 a | 4.6 ± 1.1 a | 5.3 ± 0.4 a | |
| CF | Urea | 1.5 ± 0.6 b | 1.3 ± 0.3 d | 1.8 ± 0.6 c | 1.6 ± 0.5 d | 1.6 ± 0.7 c | 1.6 ± 0.4 d | 1.9 ± 0.7 c | 1.2 ± 0.2 e |
| Control | No fertilizer | 2.2 ± 0.3 b | 2.0 ± 0.4 e | 2.5 ± 0.2 bc | 2.5 ± 0.5 cd | 2.2 ± 0.2 c | 2.4 ± 0.2 cd | 2.3 ± 0.4 bc | 2.1 ± 0.3 d |
| Fertilizer | Sampling Positions | Autumn | Early Spring | Late Spring | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| δ15N (‰) | Δ15N | δ15N (‰) | Δ15N | δ15N (‰) | Δ15N | |||||
| Average | SD | Average | SD | Average | SD | |||||
| Rapeseed cake | Top | 5.4 | 0.3 | - | 6.1 | 0.9 | - | 5.5 | 0.5 | - |
| Middle | 5.6 | 0.5 | −0.2 | 6.2 | 0.8 | −0.1 | 5.6 | 0.6 | −0.1 | |
| Basal | 4.8 | 0.5 | 0.6 | 5.5 | 0.9 | 0.7 | 5.4 | 0.6 | 0.2 | |
| Urea | Top | 0.5 | 0.3 | - | 0.1 | 0.4 | - | −0.5 | 0.6 | - |
| Middle | −0.5 | 0.8 | 1.0 | −0.7 | 0.6 | 0.8 | −0.6 | 0.8 | 0.1 | |
| Basal | 0 | 0.5 | −0.5 | −0.1 | 0.4 | −0.6 | −1.2 | 0.3 | 0.6 | |
| Control | Top | 2.1 | 0.7 | - | 1.1 | 0.6 | - | 0.7 | 0.6 | - |
| Middle | 2.4 | 0.7 | −0.3 | 1.3 | 0.8 | −0.2 | 0.4 | 0.5 | 0.3 | |
| Basal | 1.9 | 0.4 | 0.5 | 1.1 | 0.7 | 0.2 | 0.1 | 0.8 | 0.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Z.; Li, C.; Li, X.; Shao, S.; Rogers, K.M.; Li, Q.; Li, D.; Guo, H.; Huang, T.; Yuan, Y. Fertilizer Effects on the Nitrogen Isotope Composition of Soil and Different Leaf Locations of Potted Camellia sinensis over a Growing Season. Plants 2024, 13, 1628. https://doi.org/10.3390/plants13121628
Guo Z, Li C, Li X, Shao S, Rogers KM, Li Q, Li D, Guo H, Huang T, Yuan Y. Fertilizer Effects on the Nitrogen Isotope Composition of Soil and Different Leaf Locations of Potted Camellia sinensis over a Growing Season. Plants. 2024; 13(12):1628. https://doi.org/10.3390/plants13121628
Chicago/Turabian StyleGuo, Zuchuang, Chunlin Li, Xin Li, Shengzhi Shao, Karyne M. Rogers, Qingsheng Li, Da Li, Haowei Guo, Tao Huang, and Yuwei Yuan. 2024. "Fertilizer Effects on the Nitrogen Isotope Composition of Soil and Different Leaf Locations of Potted Camellia sinensis over a Growing Season" Plants 13, no. 12: 1628. https://doi.org/10.3390/plants13121628
APA StyleGuo, Z., Li, C., Li, X., Shao, S., Rogers, K. M., Li, Q., Li, D., Guo, H., Huang, T., & Yuan, Y. (2024). Fertilizer Effects on the Nitrogen Isotope Composition of Soil and Different Leaf Locations of Potted Camellia sinensis over a Growing Season. Plants, 13(12), 1628. https://doi.org/10.3390/plants13121628