

Comparative Analysis Highlights Uniconazole’s Efficacy in Enhancing the Cold Stress Tolerance of Mung Beans by Targeting Photosynthetic Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
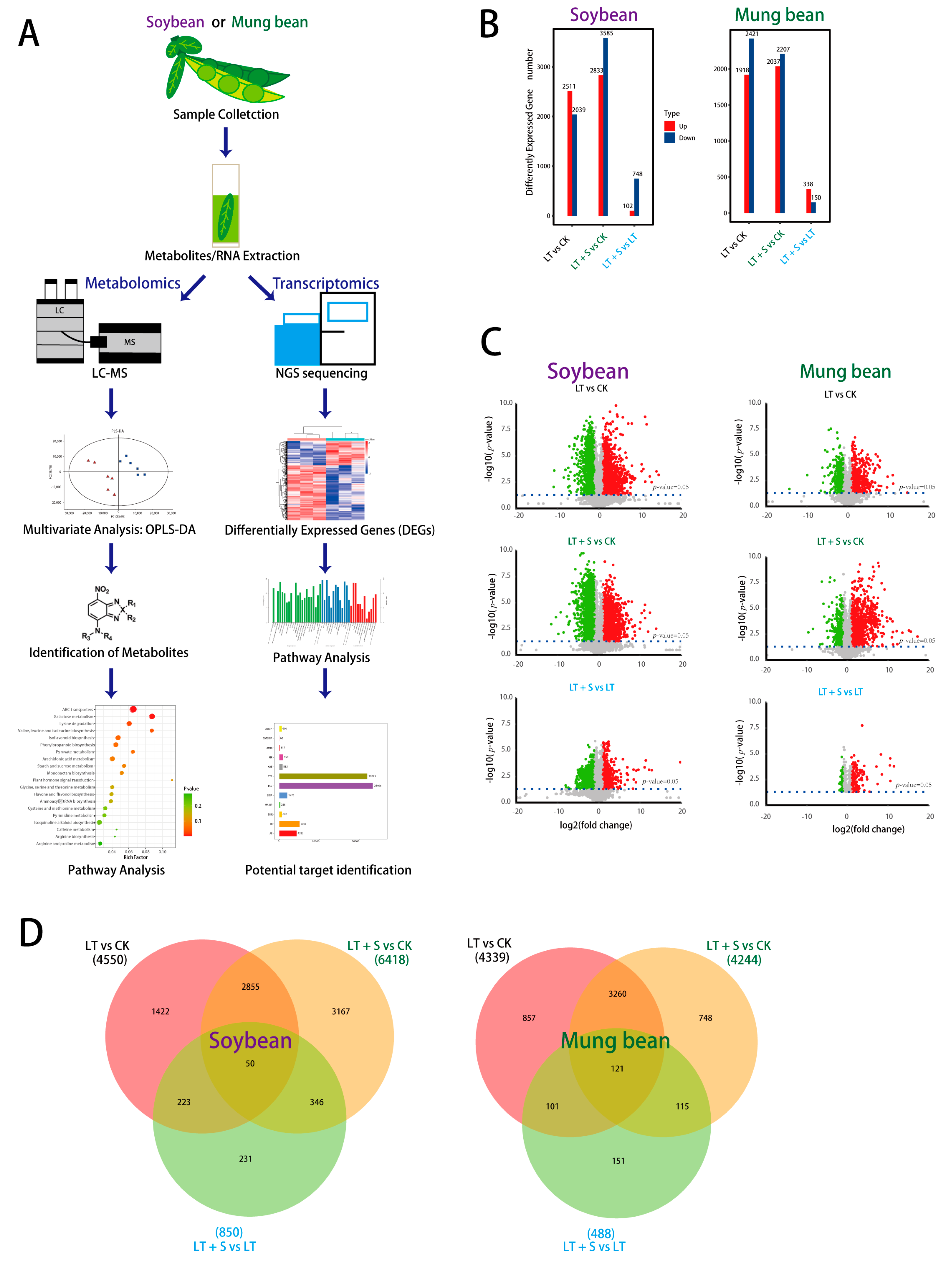
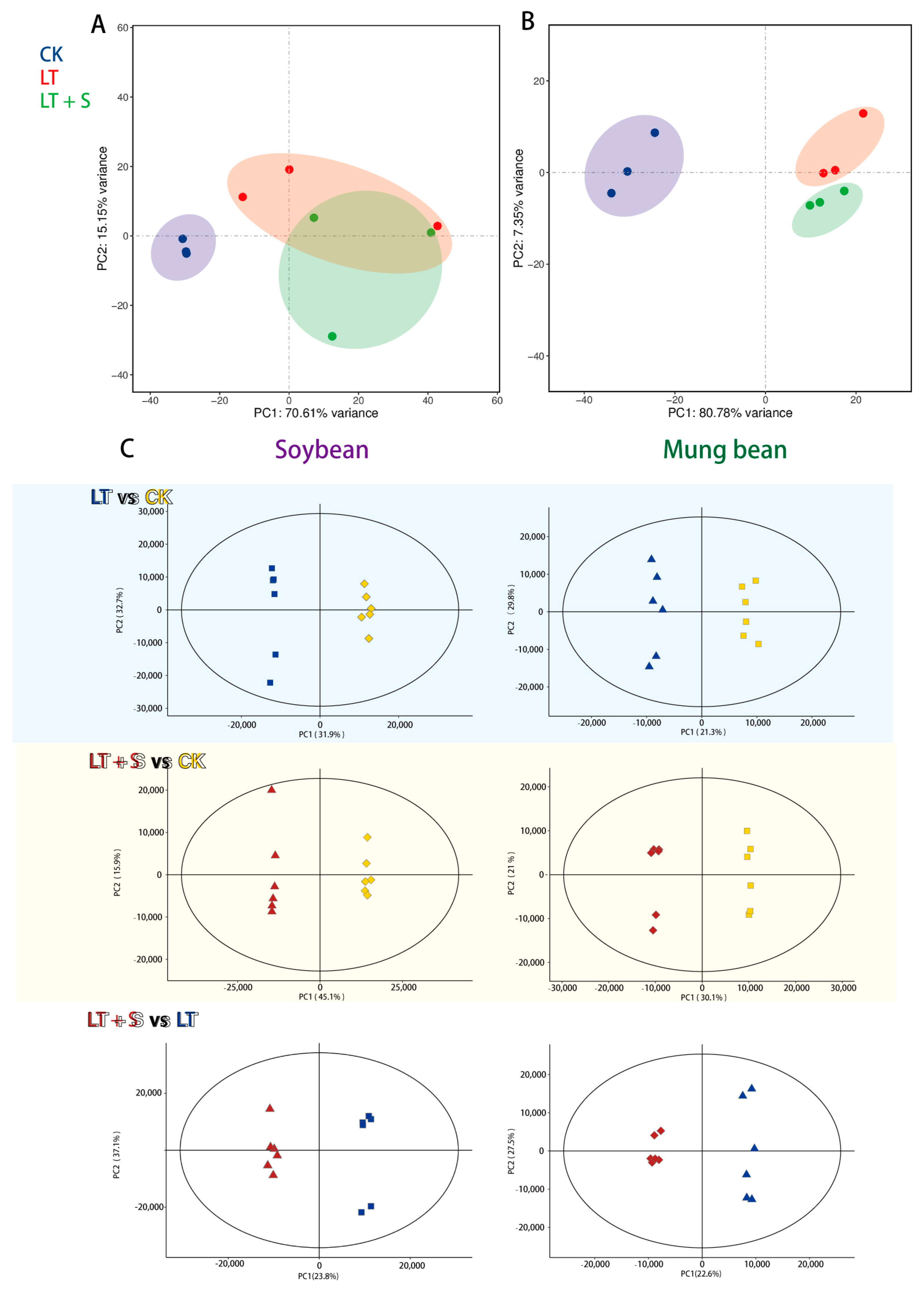
2.1. Transcriptional and Metabolic Alterations under Cold Stress
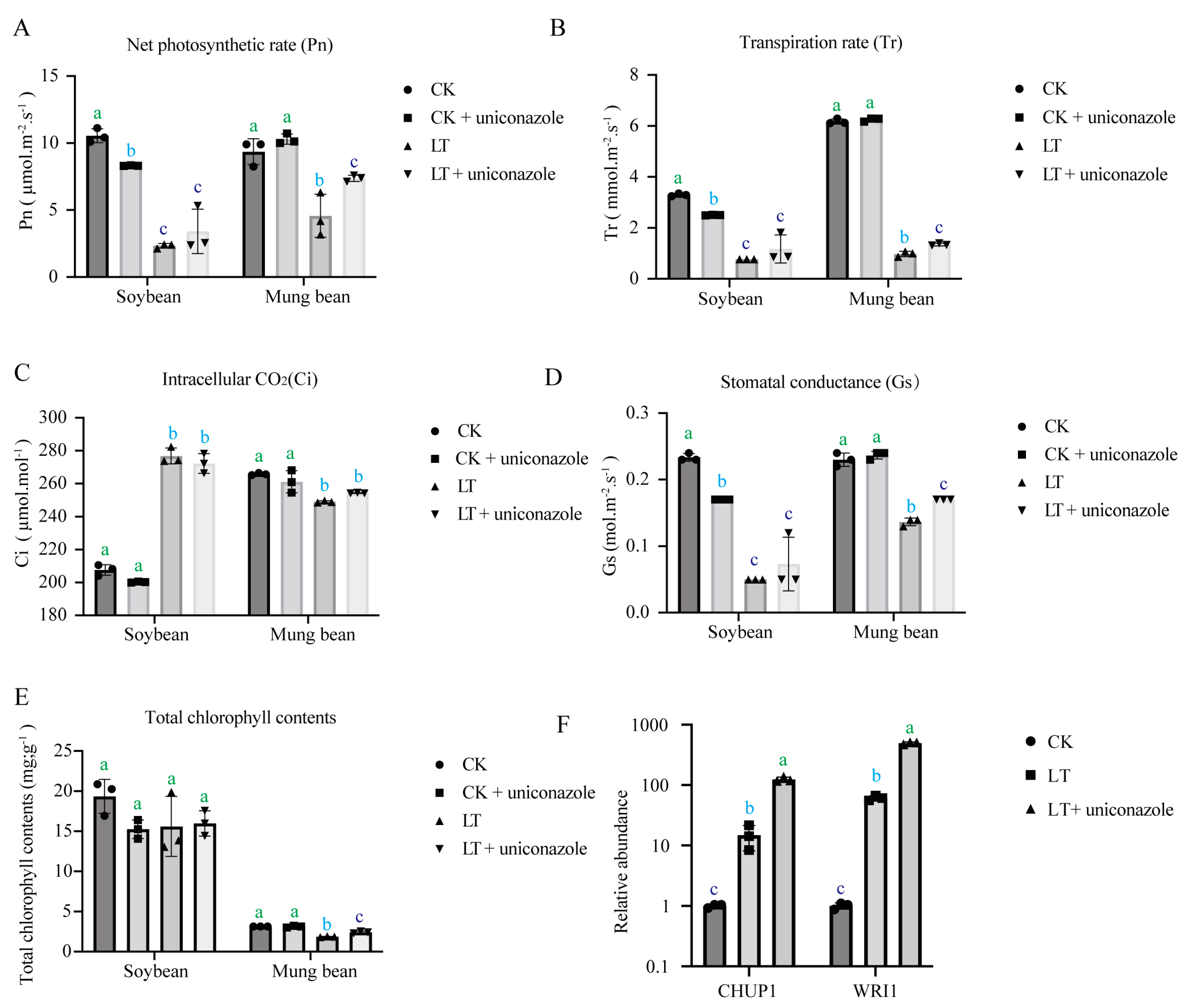
2.2. Uniconazole’s Impact on Photosynthesis under Cold Stress
2.3. Unraveling the Differential Impact on Photosynthesis-Related Pathways in Soybean and Mung Bean under Uniconazole Treatment
2.4. Investigating Uniconazole’s Interaction with Photosynthesis-Related Proteins
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Cultivated Condition
4.2. Chemical and Stress Treatment
4.3. RNA-Seq and Transcriptomics Analysis
4.4. Sample Preparation and Metabolite Analysis
4.5. Assessment of Photosynthetic Gas Exchange Parameters
4.6. Quantification of Total Chlorophyll Content
4.7. Real-Time Quantitative PCR (RT-qPCR) Assay
- β-Actin Forward (F): CGAACCACCAACCGTAGTAAA
- β-Actin Reverse (R): CAGATGAGAATGCCCGAGAG
- CHUP1 Forward (F): GGCTGCTACCAAGATTTCTACC
- CHUP1 Reverse (R): AAGCGTCTCCTCCAGTGTCA
- WRI-1 Forward (F): TCACCACCCTCACCACATTC
- WRI-1 Reverse (R): TTCAGGGTTTGAGGAAGACATT
- GPDH Forward (F): GGACCAATCACAGAATGACAAGT
- GPDH Reverse (R): AAGTTAGAAGGACAGCACACACC
4.8. Detection of Novel Transcripts, Alternative Splicing, and Genetic Variations
4.9. Protein Structure Prediction and Uniconazole Docking Analysis
4.10. Recombinant Protein Purification and ITC Affinity Test
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bouchenak, M.; Lamri-Senhadji, M. Nutritional quality of legumes, and their role in cardiometabolic risk prevention: A review. J. Med. Food 2013, 16, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Tor-Roca, A.; Garcia-Aloy, M.; Mattivi, F.; Llorach, R.; Andres-Lacueva, C.; Urpi-Sarda, M. Phytochemicals in Legumes: A Qualitative Reviewed Analysis. J. Agric. Food Chem. 2020, 68, 13486–13496. [Google Scholar] [CrossRef] [PubMed]
- Dhaliwal, S.K.; Talukdar, A.; Gautam, A.; Sharma, P.; Sharma, V.; Kaushik, P. Developments and Prospects in Imperative Underexploited Vegetable Legumes Breeding: A Review. Int. J. Mol. Sci. 2020, 21, 9615. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Sakakibara, H.; Toda, T.; Ashida, H. Insights into the potential benefits of black soybean (Glycine max L.) polyphenols in lifestyle diseases. Food Funct. 2020, 11, 7321–7339. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Yousaf, L.; Xue, Y.; Hu, J.; Wu, J.; Hu, X.; Feng, N.; Shen, Q. Mung Bean (Vigna radiata L.): Bioactive Polyphenols, Polysaccharides, Peptides, and Health Benefits. Nutrients 2019, 11, 1238. [Google Scholar] [CrossRef] [PubMed]
- Yanti; Violina, V.; Putri, C.E.; Lay, B.W. Branched Chain Amino Acid Content and Antioxidant Activity of Mung Bean Tempeh Powder for Developing Oral Nutrition Supplements. Foods 2023, 12, 2789. [Google Scholar] [CrossRef]
- Kim, B.C.; Lim, I.; Ha, J. Metabolic profiling and expression analysis of key genetic factors in the biosynthetic pathways of antioxidant metabolites in mungbean sprouts. Front. Plant Sci. 2023, 14, 1207940. [Google Scholar] [CrossRef]
- Diamond, S.E. Evolutionary potential of upper thermal tolerance: Biogeographic patterns and expectations under climate change. Ann. N. Y. Acad. Sci. 2017, 1389, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Kimura, S. Plant Temperature Sensors. Sensors 2018, 18, 4365. [Google Scholar] [CrossRef]
- Kazemi-Shahandashti, S.S.; Maali-Amiri, R. Global insights of protein responses to cold stress in plants: Signaling, defence, and degradation. J. Plant Physiol. 2018, 226, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Shi, Y.; Yang, S. Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants. New Phytol. 2019, 222, 1690–1704. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Komatsu, S. Improvement of Soybean Products Through the Response Mechanism Analysis Using Proteomic Technique. Adv. Food Nutr. Res. 2017, 82, 117–148. [Google Scholar] [PubMed]
- Li, M.W.; Xin, D.; Gao, Y.; Li, K.P.; Fan, K.; Munoz, N.B.; Yung, W.S.; Lam, H.M. Using genomic information to improve soybean adaptability to climate change. J. Exp. Bot. 2017, 68, 1823–1834. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Nair, R.M.; Lee, J.; Lee, S.H. Genomic resources in mungbean for future breeding programs. Front. Plant Sci. 2015, 6, 626. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, R.; Lone, J.K.; Pandey, R.; Mondal, N.; Dhandapani, R.; Meena, S.K.; Khan, S.; Gayacharan. Insights into morphological and physio-biochemical adaptive responses in mungbean (Vigna radiata L.) under heat stress. Front. Genet. 2023, 14, 1206451. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Singh, S.; Barnaby, J.; Buyer, J.; Reddy, V.; Sicher, R. Effects of growth temperature and carbon dioxide enrichment on soybean seed components at different stages of development. Plant Physiol. Biochem. 2016, 108, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K.; Ichii, M.; Asanuma, K.; Kusutani, A. Optimum and sub-optimal temperature effects on stomata and photosynthesis rate of determinate soybeans. Acta Hortic. 1996, 440, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.; Cai, Z.; Li, Y.; Suo, H.; Yi, R.; Zhang, S.; Nian, H. The Floral Repressor GmFLC-like Is Involved in Regulating Flowering Time Mediated by Low Temperature in Soybean. Int. J. Mol. Sci. 2020, 21, 1322. [Google Scholar] [CrossRef] [PubMed]
- Yuan, K.; Reckling, M.; Ramirez, M.D.A.; Djedidi, S.; Fukuhara, I.; Ohyama, T.; Yokoyama, T.; Bellingrath-Kimura, S.D.; Halwani, M.; Egamberdieva, D.; et al. Characterization of Rhizobia for the Improvement of Soybean Cultivation at Cold Conditions in Central Europe. Microbes Environ. 2020, 35, ME19124. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.R.; Ko, C.Y.; Folk, W.R.; Lin, T.Y. Chilling susceptibility in mungbean varieties is associated with their differentially expressed genes. Bot. Stud. 2017, 58, 7. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.M.; Pandey, A.K.; War, A.R.; Hanumantharao, B.; Shwe, T.; Alam, A.; Pratap, A.; Malik, S.R.; Karimi, R.; Mbeyagala, E.K.; et al. Biotic and Abiotic Constraints in Mungbean Production-Progress in Genetic Improvement. Front. Plant Sci. 2019, 10, 1340. [Google Scholar] [CrossRef] [PubMed]
- Serrano, M.; Robertson, A.E. The Effect of Cold Stress on Damping-Off of Soybean Caused by Pythium sylvaticum. Plant Dis. 2018, 102, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Eremina, M.; Rozhon, W.; Poppenberger, B. Hormonal control of cold stress responses in plants. Cell Mol. Life Sci. 2016, 73, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.A.; Li, S.; Gao, H.; Feng, C.; Sun, P.; Sui, X.; Jing, Y.; Xu, K.; Zhou, Y.; Zhang, W.; et al. Comparative analysis of physiological variations and genetic architecture for cold stress response in soybean germplasm. Front. Plant Sci. 2022, 13, 1095335. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Mochida, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S. Genome-wide expression profiling of soybean two-component system genes in soybean root and shoot tissues under dehydration stress. DNA Res. 2011, 18, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Balestrasse, K.B.; Tomaro, M.L.; Batlle, A.; Noriega, G.O. The role of 5-aminolevulinic acid in the response to cold stress in soybean plants. Phytochemistry 2010, 71, 2038–2045. [Google Scholar] [CrossRef] [PubMed]
- Muncan, J.; Jinendra, B.M.S.; Kuroki, S.; Tsenkova, R. Aquaphotomics Research of Cold Stress in Soybean Cultivars with Different Stress Tolerance Ability: Early Detection of Cold Stress Response. Molecules 2022, 27, 744. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.W.; Murphy, T.M.; Lin, C.H. Hydrogen peroxide-induced chilling tolerance in mung beans mediated through ABA-independent glutathione accumulation. Funct. Plant Biol. 2003, 30, 955–963. [Google Scholar] [CrossRef]
- Yousefi, S.; Marchese, A.; Salami, S.A.; Benny, J.; Giovino, A.; Perrone, A.; Caruso, T.; Gholami, M.; Sarikhani, H.; Buti, M.; et al. Identifying conserved genes involved in crop tolerance to cold stress. Funct. Plant Biol. 2022, 49, 861–873. [Google Scholar] [CrossRef] [PubMed]
- Sanghera, G.S.; Wani, S.H.; Hussain, W.; Singh, N.B. Engineering cold stress tolerance in crop plants. Curr. Genom. 2011, 12, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Anwar, A.; Bai, L.; Miao, L.; Liu, Y.; Li, S.; Yu, X.; Li, Y. 24-Epibrassinolide Ameliorates Endogenous Hormone Levels to Enhance Low-Temperature Stress Tolerance in Cucumber Seedlings. Int. J. Mol. Sci. 2018, 19, 2497. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Li, H.; Chen, X.; Xiang, X.; Guo, Z.; Yu, J.; Zhou, Y. The role of calcium-dependent protein kinase in hydrogen peroxide, nitric oxide and ABA-dependent cold acclimation. J. Exp. Bot. 2018, 69, 4127–4139. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Yang, S.; Wang, Y.; Lu, L.; Sun, M.; He, C.; Wang, J.; Li, Y.; Yu, X.; Li, Q.; et al. Physiological and Molecular Mechanisms of ABA and CaCl2 Regulating Chilling Tolerance of Cucumber Seedlings. Plants 2021, 10, 2746. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.A.; Fariduddin, Q.; Yusuf, M. Lycopersicon esculentum under low temperature stress: An approach toward enhanced antioxidants and yield. Environ. Sci. Pollut. Res. Int. 2015, 22, 14178–14188. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Liu, T.; Ren, K.; Chen, J.; Zhao, G.; Hu, B.; Xu, A.; Jin, Y.; Zhu, Y.; Zou, C. Salicylic Acid Effects on Flue-Cured Tobacco Quality and Curing Characteristics During Harvesting and Curing in Cold-Stressed Fields. Front. Plant Sci. 2020, 11, 580597. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guo, Y.; Lan, Z.; Xu, K.; Chang, J.; Ahammed, G.J.; Ma, J.; Wei, C.; Zhang, X. Methyl jasmonate mediates melatonin-induced cold tolerance of grafted watermelon plants. Hortic. Res. 2021, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Repkina, N.; Ignatenko, A.; Holoptseva, E.; Miszalsk, I.Z.; Kaszycki, P.; Talanova, V. Exogenous Methyl Jasmonate Improves Cold Tolerance with Parallel Induction of Two Cold-Regulated (COR) Genes Expression in Triticum aestivum L. Plants 2021, 10, 1421. [Google Scholar] [CrossRef] [PubMed]
- Elbagoury, M.M.; Turoop, L.; Runo, S.; Sila, D.N. Regulatory influences of methyl jasmonate and calcium chloride on chilling injury of banana fruit during cold storage and ripening. Food Sci. Nutr. 2021, 9, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yue, C.; Xiang, J.; Han, Y.; Wang, J.; Wang, L.; Sun, L. Gene expression profile indicates involvement of uniconazole in Coix lachryma-jobi L. seedlings at low temperature. Food Sci. Nutr. 2020, 8, 534–546. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Wang, S.; Liang, X.; Wang, X.; Xie, H.; Wang, D.; Gai, Z.; Wang, N.; Xiang, P.; Han, D.; et al. Foliar spraying of exogenous uniconazole (S3307) at the flowering stage as an effective method to resist low-temperature stress on mung bean [Vigna radiata (L.) Wilczek]. Sci. Rep. 2023, 13, 22331. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.J.; Feng, N.F.; Wang, X.X.; Cai, G.R.; Cao, M.Y.; Zheng, D.F.; Zhu, H.D. Uniconazole confers chilling stress tolerance in soybean (Glycine max L.) by modulating photosynthesis, photoinhibition, and activating oxygen metabolism system. Photosynthetica 2019, 57, 446–457. [Google Scholar] [CrossRef]
- Yoshida, S. Low Temperature-Induced Cytoplasmic Acidosis in Cultured Mung Bean (Vigna radiata [L.] Wilczek) Cells. Plant Physiol. 1994, 104, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.B.; Wang, C.L.; Wu, J.Y.; Zhou, H.S.; Jiang, X.T.; Wu, J.; Zhang, S.L. Low temperature inhibits pollen tube growth by disruption of both tip-localized reactive oxygen species and endocytosis in Pyrus bretschneideri Rehd. Plant Physiol. Biochem. 2014, 74, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liu, D.; Chong, K. Cold signaling in plants: Insights into mechanisms and regulation. J. Integr. Plant Biol. 2018, 60, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Peng, L.; Ji, L.; Wang, S.; Wang, L.; Wu, J. Genome-wise association study identified genomic regions associated with drought tolerance in mungbean (Vigna radiata (L.) R. Wilczek). Theor. Appl. Genet. 2023, 136, 40. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Ye, L.; Wang, Y.; Zhou, X.; Yang, J.; Wang, J.; Cao, K.; Zou, Z. Melatonin Increases the Chilling Tolerance of Chloroplast in Cucumber Seedlings by Regulating Photosynthetic Electron Flux and the Ascorbate-Glutathione Cycle. Front. Plant Sci. 2016, 7, 1814. [Google Scholar] [CrossRef] [PubMed]
- Bashir, K.; Matsui, A.; Rasheed, S.; Seki, M. Recent advances in the characterization of plant transcriptomes in response to drought, salinity, heat, and cold stress. F1000Res 2019, 8, 658. [Google Scholar] [CrossRef] [PubMed]
- Jha, U.C.; Bohra, A.; Jha, R. Breeding approaches and genomics technologies to increase crop yield under low-temperature stress. Plant Cell Rep. 2017, 36, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, B.; Jappe, U.; Novak, N. Allergy to Peanut, Soybean, and Other Legumes: Recent Advances in Allergen Characterization, Stability to Processing and IgE Cross-Reactivity. Mol. Nutr. Food Res. 2018, 62, 1700446. [Google Scholar] [CrossRef] [PubMed]
- Bidadi, H.; Yamaguchi, S.; Asahina, M.; Satoh, S. Effects of shoot-applied gibberellin/gibberellin-biosynthesis inhibitors on root growth and expression of gibberellin biosynthesis genes in Arabidopsis thaliana. Plant Root 2010, 4, 4–11. [Google Scholar] [CrossRef]
- Jacobsen, S.E.; Olszewski, N.E. Mutations at the SPINDLY locus of Arabidopsis alter gibberellin signal transduction. Plant Cell 1993, 5, 887–896. [Google Scholar] [PubMed]
- Ingolia, N.T.; Brar, G.A.; Rouskin, S.; McGeachy, A.M.; Weissman, J.S. The ribosome profiling strategy for monitoring translation in vivo by deep sequencing of ribosome-protected mRNA fragments. Nat. Protoc. 2012, 7, 1534–1550. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation (Camb) 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed]
- Siren, J.; Valimaki, N.; Makinen, V. Indexing Graphs for Path Queries with Applications in Genome Research. IEEE/ACM Trans. Comput. Biol. Bioinform. 2014, 11, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome Project Data Processing, S. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Danecek, P.; McCarthy, S.A. BCFtools/csq: Haplotype-aware variant consequences. Bioinformatics 2017, 33, 2037–2039. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2021, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, M.; Dai, S.; Chen, D.; Zhu, P.; Feng, N.; Zheng, D. Comparative Analysis Highlights Uniconazole’s Efficacy in Enhancing the Cold Stress Tolerance of Mung Beans by Targeting Photosynthetic Pathways. Plants 2024, 13, 1885. https://doi.org/10.3390/plants13141885
Chen M, Dai S, Chen D, Zhu P, Feng N, Zheng D. Comparative Analysis Highlights Uniconazole’s Efficacy in Enhancing the Cold Stress Tolerance of Mung Beans by Targeting Photosynthetic Pathways. Plants. 2024; 13(14):1885. https://doi.org/10.3390/plants13141885
Chicago/Turabian StyleChen, Mingming, Shuangfeng Dai, Daming Chen, Peiyi Zhu, Naijie Feng, and Dianfeng Zheng. 2024. "Comparative Analysis Highlights Uniconazole’s Efficacy in Enhancing the Cold Stress Tolerance of Mung Beans by Targeting Photosynthetic Pathways" Plants 13, no. 14: 1885. https://doi.org/10.3390/plants13141885
APA StyleChen, M., Dai, S., Chen, D., Zhu, P., Feng, N., & Zheng, D. (2024). Comparative Analysis Highlights Uniconazole’s Efficacy in Enhancing the Cold Stress Tolerance of Mung Beans by Targeting Photosynthetic Pathways. Plants, 13(14), 1885. https://doi.org/10.3390/plants13141885