
Phosphorus-Solubilizing Bacteria Enhance Cadmium Immobilization and Gene Expression in Wheat Roots to Reduce Cadmium Uptake

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
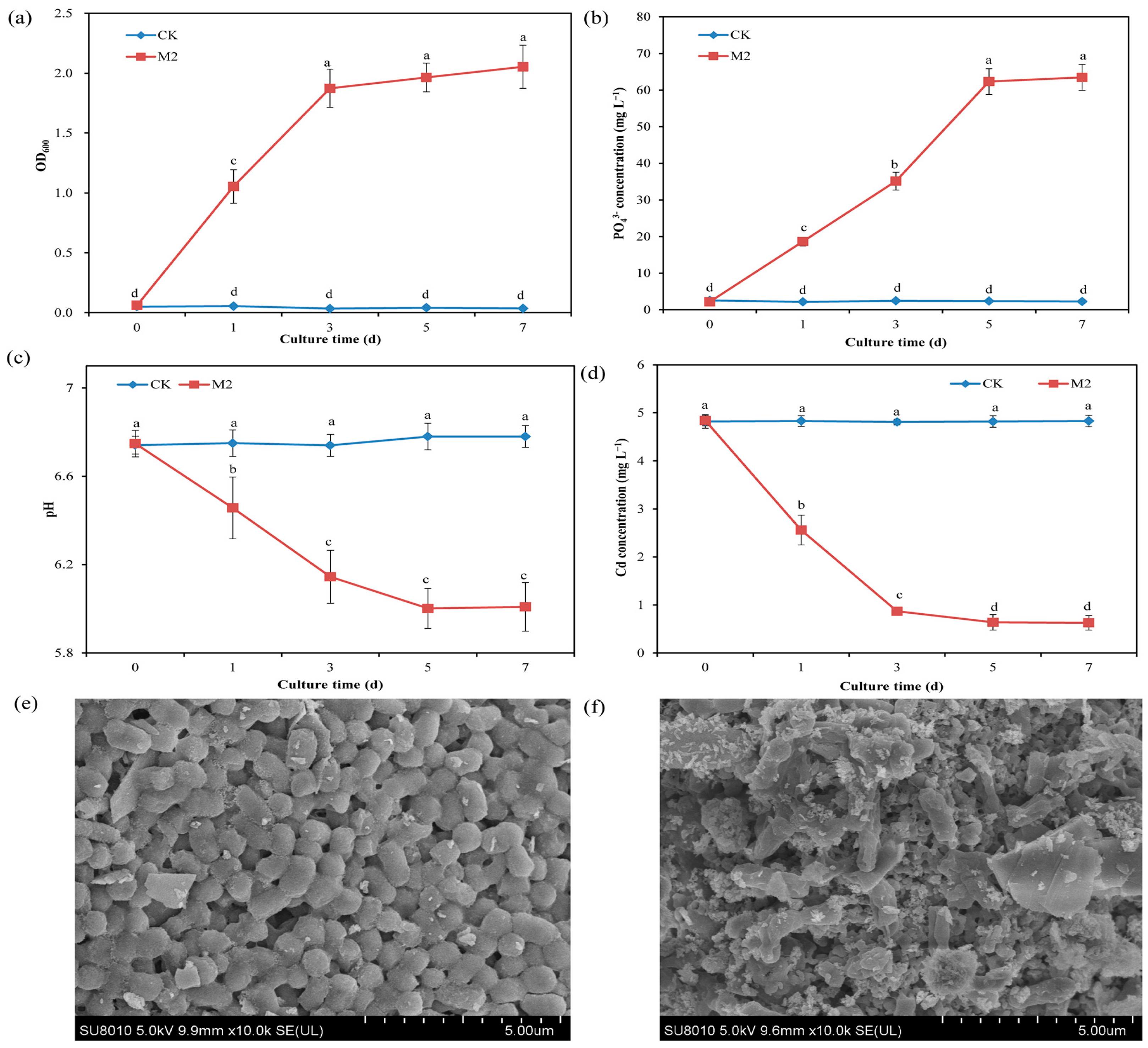
2.1. Mechanisms of Cd Immobilization by Strain M2
2.2. Effects of Strain M2 on the Growth of Wheat and Accumulation of Cd
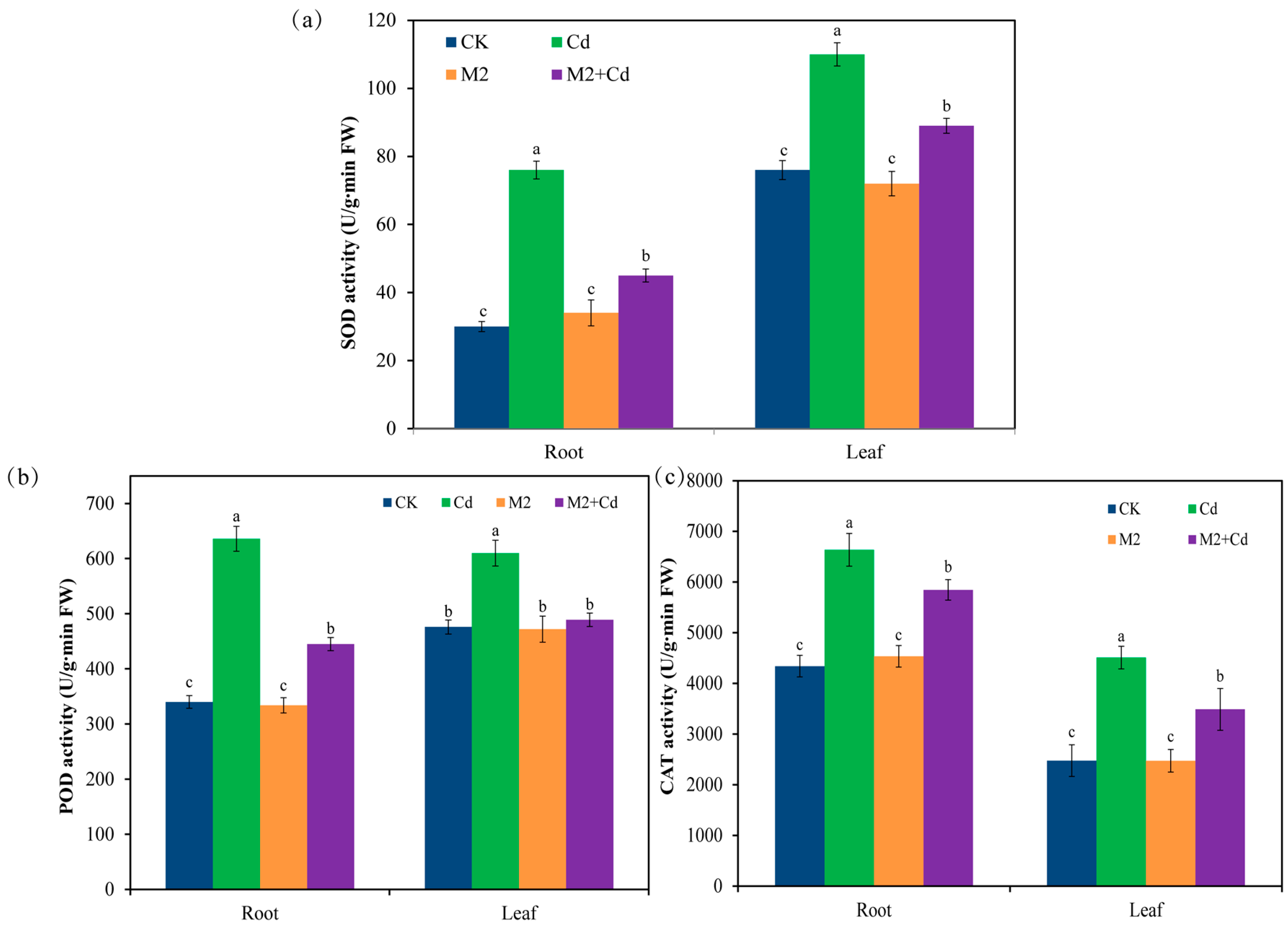
2.3. Effects of Strain M2 on Antioxidant Enzyme Activity in Wheat
2.4. Effects of Strain M2 on Cd Adsorption on Wheat Roots
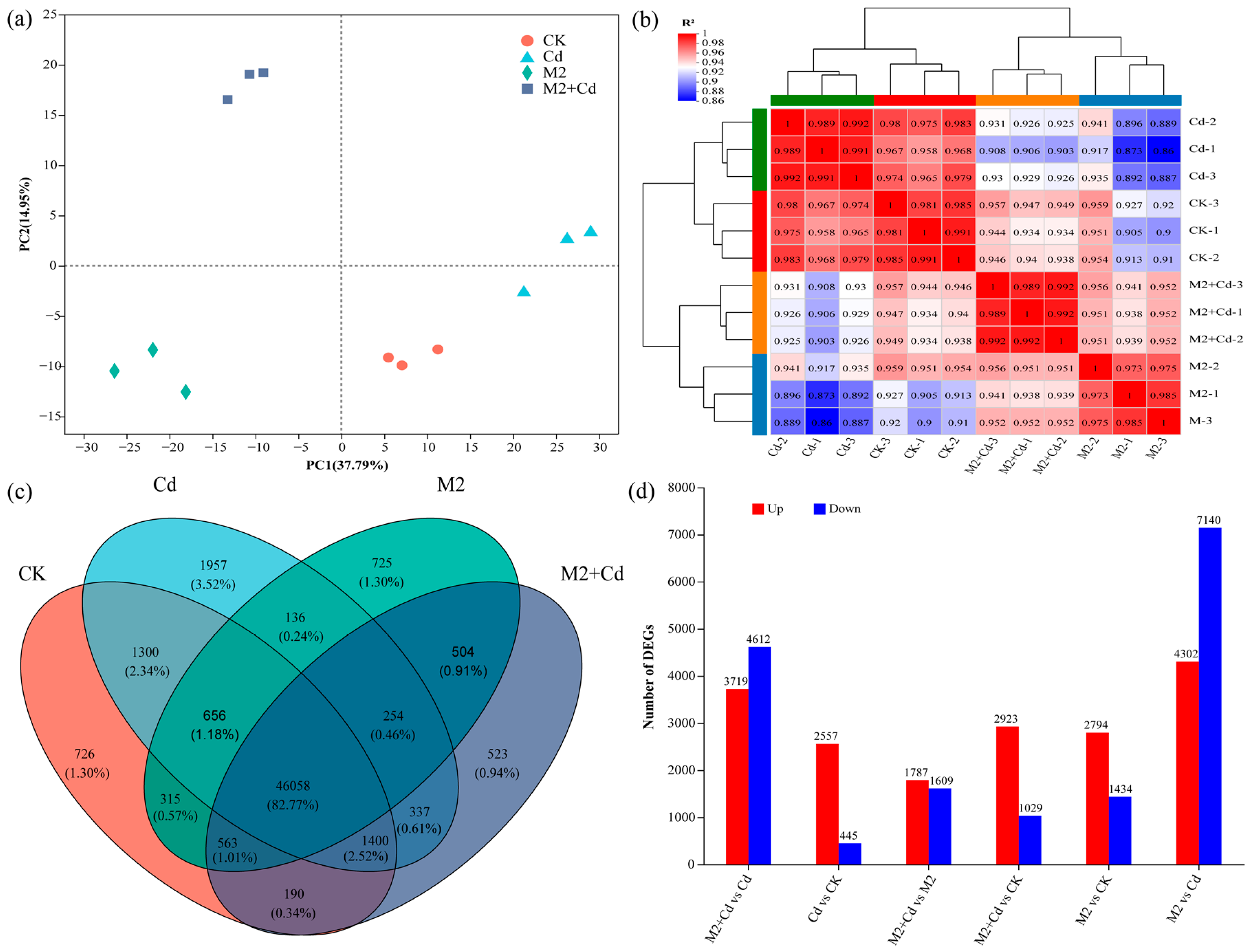
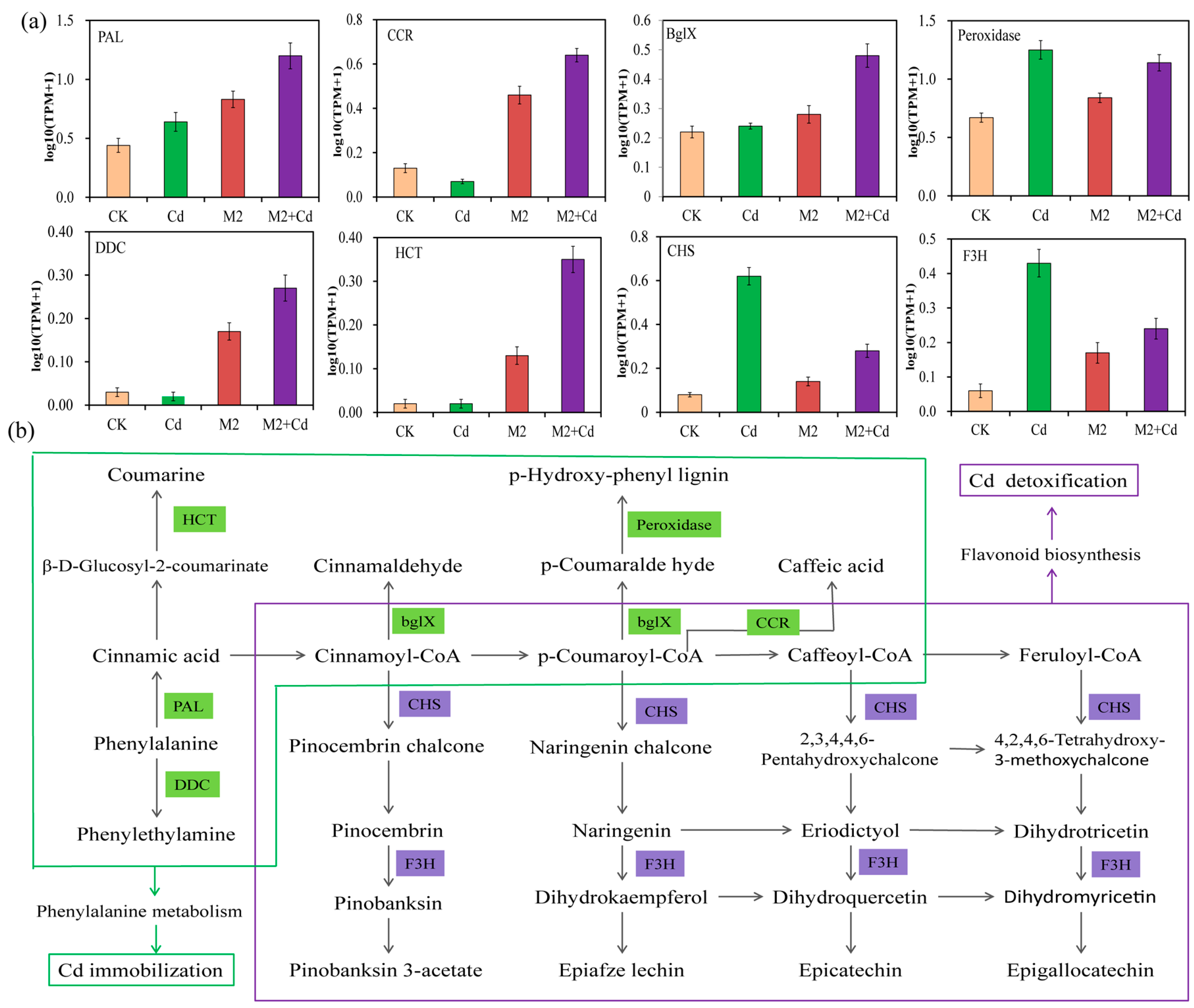
2.5. Effects of Strain M2 on Gene Expression in Wheat Roots
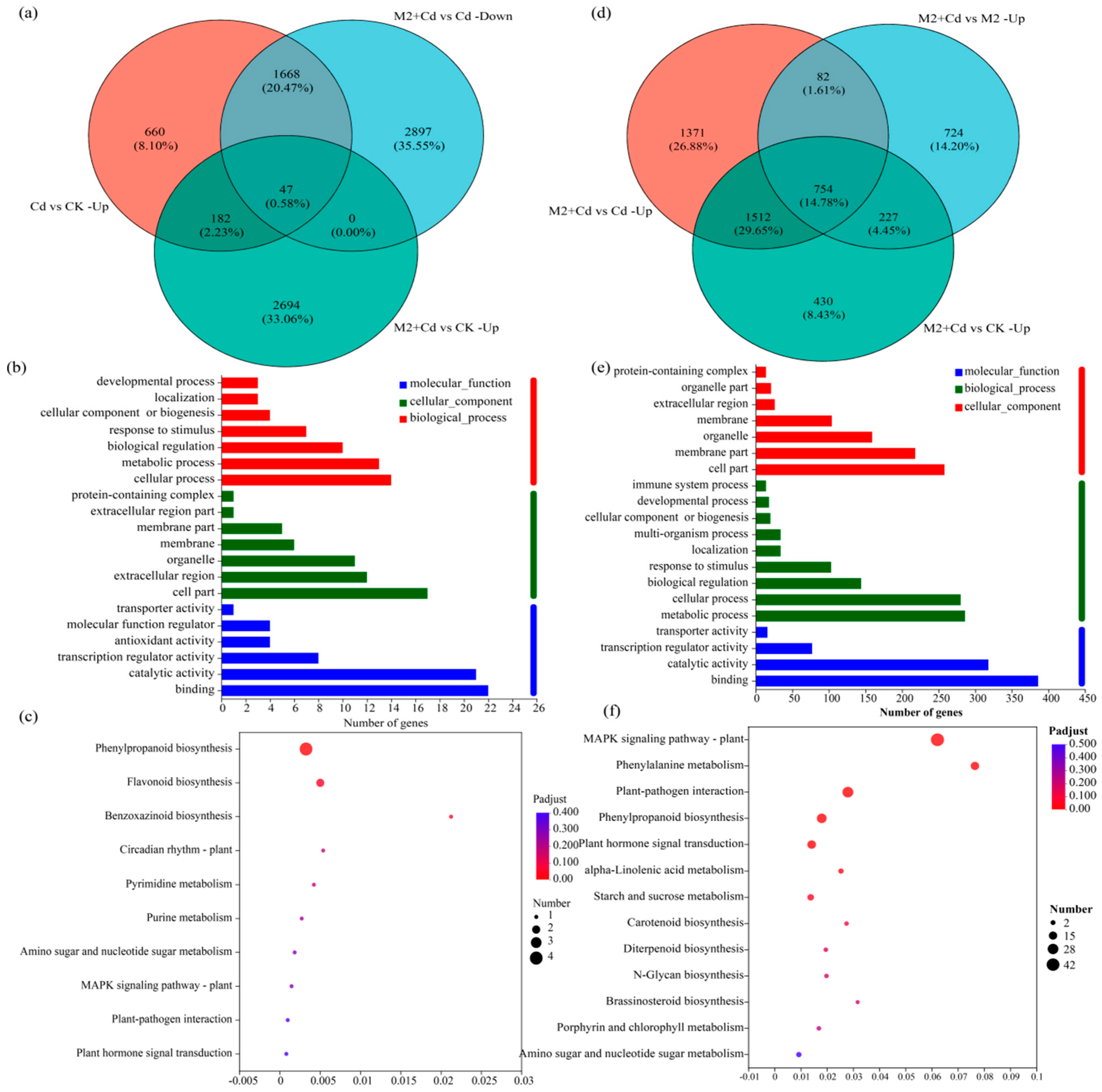
2.6. Transcriptional Regulation of M2-Mediated Cd Detoxification
2.7. Transcriptional Regulation of Strain M2-Mediated Cd Immobilization
2.8. Activity of Signaling Pathways Induced by Strain M2
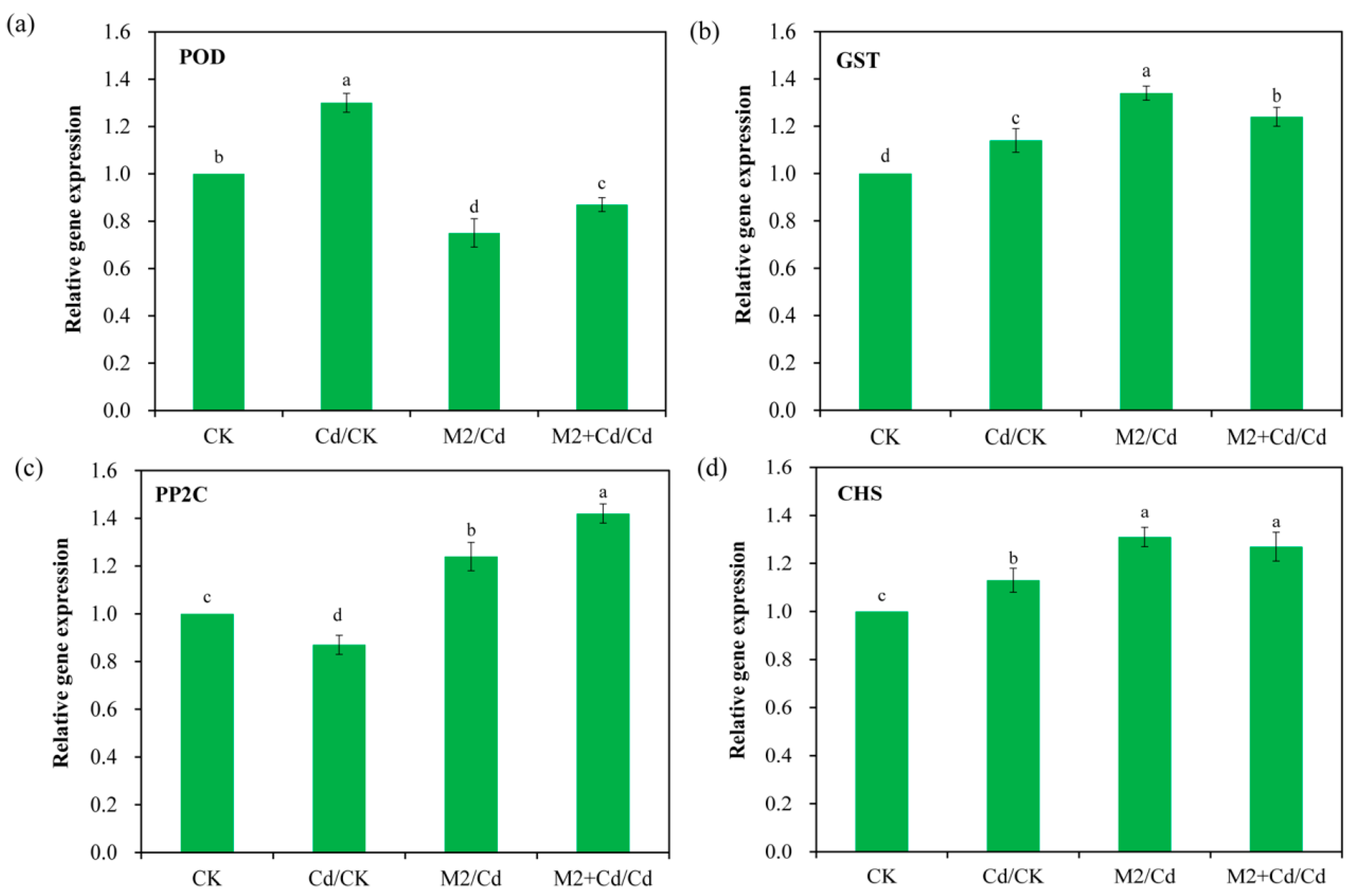
2.9. qRT–PCR Verification
3. Discussion
3.1. Effects of Strain M2 Inoculation on the Uptake and Separation of Cd in Wheat Roots
3.2. Effects of Strain M2 Inoculation on Cd Detoxification in Wheat Roots
3.3. Effects of Strain M2 Inoculation on Cd Immobilization in Wheat Roots
4. Materials and Methods
4.1. Phosphorus-Solubilizing Bacteria
4.2. Cd Immobilization Effect of Strain M2
4.3. Wheat Cultivation Experiment
4.4. Determination of Wheat Root and Leaf Dry Weights and Cd Content
4.5. Determination of the Cd Content on the Surface of Wheat Roots
4.6. Cd Subcellular Distribution in Wheat Roots
4.7. Determination of Differentially Expressed Genes in Wheat Roots
4.8. Quantitative Real-Time PCR (qRT–PCR) Verification
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ren, S.; Song, C.; Ye, S.; Cheng, C.; Gao, P. The spatiotemporal variation in heavy metals in China’s farmland soil over the past 20 years: A meta-analysis. Sci. Total Environ. 2022, 806, 150322. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-X.; Liu, Y.-M.; Zhao, Q.-Y.; Cao, W.-Q.; Chen, X.-P.; Zou, C.-Q. Health risk assessment associated with heavy metal accumulation in wheat after long-term phosphorus fertilizer application. Environ. Pollut. 2020, 262, 114348. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, Y.; Yang, Y.; Wang, M.; Chen, W. Bayesian risk prediction model: An accessible strategy to predict cadmium contamination risk in wheat grain grown in alkaline soils. Environ. Pollut. 2024, 354, 124169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, C.; Du, B.; Lu, B.; Zhou, D.; Zhou, J.; Zhou, J. Effects of node restriction on cadmium accumulation in eight Chinese wheat (Triticum turgidum) cultivars. Sci. Total Environ. 2020, 725, 138358. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Xu, Y.; Nie, Z.; Liu, H.; Gao, W.; Li, C.; Wang, L.; Zhao, P. Effect of boron on cadmium uptake and expression of Cd transport genes at different growth stages of wheat (Triticum aestivum L.). Ecotoxicol. Environ. Saf. 2022, 241, 113834. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Li, L.; Ippolito, J.A.; Xing, W.; Wang, Y.; Wang, Y.; Cheng, Y.; Qiu, K. Heavy metal distribution in wheat plant components following foliar Cd application. Chemosphere 2023, 322, 138177. [Google Scholar] [CrossRef]
- Janes-Bassett, V.; Blackwell, M.S.A.; Blair, G.; Davies, J.; Haygarth, P.M.; Mezeli, M.M.; Stewart, G. A meta-analysis of phosphatase activity in agricultural settings in response to phosphorus deficiency. Soil Biol. Biochem. 2022, 165, 108537. [Google Scholar] [CrossRef]
- Beheshti, M.; Alikhani, H.A.; Pourbabaee, A.A.; Etesami, H.; Asadi Rahmani, H.; Noroozi, M. Enriching periphyton with phosphate-solubilizing microorganisms improves the growth and concentration of phosphorus and micronutrients of rice plant in calcareous paddy soil. Rhizosphere 2022, 24, 100590. [Google Scholar] [CrossRef]
- Li, Y.; Guo, S.; Zheng, Y.; Yu, J.; Chi, R.; Xiao, C. Bioimmobilization of lead in phosphate mining wasteland by isolated strain Citrobacter farmeri CFI-01. Environ. Pollut. 2022, 307, 119485. [Google Scholar] [CrossRef]
- Ma, S.; Nan, Z.; Hu, Y.; Chen, S.; Yang, X.; Su, J. Phosphorus supply level is more important than wheat variety in safe utilization of cadmium-contaminated calcareous soil. J. Hazard. Mater. 2022, 424, 127224. [Google Scholar] [CrossRef]
- Khourchi, S.; Elhaissoufi, W.; Loum, M.; Ibnyasser, A.; Haddine, M.; Ghani, R.; Barakat, A.; Zeroual, Y.; Rchiad, Z.; Delaplace, P.; et al. Phosphate solubilizing bacteria can significantly contribute to enhance P availability from polyphosphates and their use efficiency in wheat. Microbiol. Res. 2022, 262, 127094. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Min, F.; Hu, X.; Ma, D.; Huo, Z. Biochar assists phosphate solubilizing bacteria to resist combined Pb and Cd stress by promoting acid secretion and extracellular electron transfer. J. Hazard. Mater. 2023, 452, 131176. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Zhang, H.; He, Y.; Chen, Z.; Yao, L.; Han, H. Improving radish phosphorus utilization efficiency and inhibiting Cd and Pb uptake by using heavy metal-immobilizing and phosphate-solubilizing bacteria. Sci. Total Environ. 2023, 868, 161685. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Zeng, M.; Williams, D.V.; Hu, W.; Shabala, S.; Zhou, M.; Cao, F. Integration of Transcriptome and Metabolome Analyses Reveals the Mechanistic Basis for Cadmium Accumulation in Maize. iScience 2022, 25, 105484. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Shen, R.F.; Shao, J.F. Transport of cadmium from soil to grain in cereal crops: A review. Pedosphere 2021, 31, 3–10. [Google Scholar] [CrossRef]
- Wu, L.; Yu, Y.; Sui, X.; Tao, Y.; Zhatova, H.; Song, P.; Li, D.; Guan, Y.; Gao, H.; Volodymyr, T.; et al. A novel wheat β-amylase gene TaBMY1 reduces Cd accumulation in common wheat grains. Environ. Exp. Bot. 2022, 203, 105050. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 Is a Major Transporter Responsible for Manganese and Cadmium Uptake in Rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Qiu, L.; Shen, P.; Wang, Y.; Li, J.; Dai, Z.; Qi, M.; Zhou, Y.; Zou, Z. Transcriptome studies on cadmium tolerance and biochar mitigating cadmium stress in muskmelon. Plant Physiol. Biochem. 2023, 197, 107661. [Google Scholar] [CrossRef]
- Liu, H.; Jiao, Q.; Fan, L.; Jiang, Y.; Alyemeni, M.N.; Ahmad, P.; Chen, Y.; Zhu, M.; Liu, H.; Zhao, Y.; et al. Integrated physio-biochemical and transcriptomic analysis revealed mechanism underlying of Si-mediated alleviation to cadmium toxicity in wheat. J. Hazard. Mater. 2023, 452, 131366. [Google Scholar] [CrossRef]
- Chen, X.; Yang, S.; Ma, J.; Huang, Y.; Wang, Y.; Zeng, J.; Li, J.; Li, S.; Long, D.; Xiao, X.; et al. Manganese and copper additions differently reduced cadmium uptake and accumulation in dwarf Polish wheat (Triticum polonicum L.). J. Hazard. Mater. 2023, 448, 130998. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.; Cheng, L.; Zhai, X.; Wu, R.; Liu, W.; Pan, J.; Shohag, M.J.I.; Xin, X.; He, Z.; Yang, X. Foliar spray of combined metal-oxide nanoparticles alters the accumulation, translocation and health risk of Cd in wheat (Triticum aestivum L.). J. Hazard. Mater. 2022, 440, 129857. [Google Scholar] [CrossRef]
- Zhu, Q.; Xu, P.; Lei, L.; Jing, Y. Transcriptome analysis reveals decreased accumulation and toxicity of Cd in upland rice inoculated with arbuscular mycorrhizal fungi. Appl. Soil Ecol. 2022, 177, 104501. [Google Scholar] [CrossRef]
- Yue, Z.; Chen, Y.; Chen, C.; Ma, K.; Tian, E.; Wang, Y.; Liu, H.; Sun, Z. Endophytic Bacillus altitudinis WR10 alleviates Cu toxicity in wheat by augmenting reactive oxygen species scavenging and phenylpropanoid biosynthesis. J. Hazard. Mater. 2021, 405, 124272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, Y.; Wang, D.; Yu, D.; Wu, C. Lignin-based adsorbents for heavy metals. Ind. Crops Prod. 2023, 193, 116119. [Google Scholar] [CrossRef]
- Ge, Y.; Li, Z. Application of Lignin and Its Derivatives in Adsorption of Heavy Metal Ions in Water: A Review. ACS Sustain. Chem. Eng. 2018, 6, 7181–7192. [Google Scholar] [CrossRef]
- Adhikari, P.; Pandey, A. Phosphate solubilization potential of endophytic fungi isolated from Taxus wallichiana Zucc. roots. Rhizosphere 2019, 9, 2–9. [Google Scholar] [CrossRef]
- Biswas, S.S.; Biswas, D.R.; Ghosh, A.; Sarkar, A.; Das, A.; Roy, T. Phosphate solubilizing bacteria inoculated low-grade rock phosphate can supplement P fertilizer to grow wheat in sub-tropical inceptisol. Rhizosphere 2022, 23, 100556. [Google Scholar] [CrossRef]
- Rawat, P.; Das, S.; Shankhdhar, D.; Shankhdhar, S.C. Phosphate-Solubilizing Microorganisms: Mechanism and Their Role in Phosphate Solubilization and Uptake. J. Soil Sci. Plant Nutr. 2021, 21, 49–68. [Google Scholar] [CrossRef]
- Zhang, J.; Xiao, Q.; Wang, P. Phosphate-solubilizing bacterium Burkholderia sp. strain N3 facilitates the regulation of gene expression and improves tomato seedling growth under cadmium stress. Ecotoxicol. Environ. Saf. 2021, 217, 112268. [Google Scholar] [CrossRef]
- Cheng, C.; Wang, Q.; Wang, Q.-X.; He, L.-Y.; Sheng, X.-F. Wheat-associated Pseudomonas taiwanensis WRS8 reduces cadmium uptake by increasing root surface cadmium adsorption and decreasing cadmium uptake and transport related gene expression in wheat. Environ. Pollut. 2021, 268, 115850. [Google Scholar] [CrossRef] [PubMed]
- Priya, A.K.; Gnanasekaran, L.; Dutta, K.; Rajendran, S.; Balakrishnan, D.; Soto-Moscoso, M. Biosorption of heavy metals by microorganisms: Evaluation of different underlying mechanisms. Chemosphere 2022, 307, 135957. [Google Scholar] [CrossRef]
- Zandi, P.; Xia, X.; Yang, J.; Liu, J.; Remusat, L.; Rumpel, C.; Bloem, E.; Krasny, B.B.; Schnug, E. Speciation and distribution of chromium (III) in rice root tip and mature zone: The significant impact of root exudation and iron plaque on chromium bioavailability. J. Hazard. Mater. 2023, 448, 130992. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Yang, Y.; Shan, Q.; Zhang, H.; Zhou, A.; Gong, S.; Chai, T.; Qiao, K. Plant cadmium resistance 10 enhances tolerance to toxic heavy metals in poplar. Plant Physiol. Biochem. 2023, 203, 108043. [Google Scholar] [CrossRef]
- Chen, K.; Xue, W.; Di, X.; Sun, T.; Gao, W.; Sun, Y. Effects of nitrogen forms on Cd uptake and tolerance in wheat seedlings. Sci. Total Environ. 2024, 936, 173451. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Zhang, Y.; Li, D.; Shan, Q.; Hu, Z.; Chai, T.; Zhou, A.; Qiao, K. Synergistic role of phenylpropanoid biosynthesis and citrate cycle pathways in heavy metal detoxification through secretion of organic acids. J. Hazard. Mater. 2024, 476, 135106. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-f.; Chen, D.-w.; Ma, J.; Gao, R.-c.; Bai, J.; Hou, Q.-z. Transcriptomic and physiological analyses of Symphytum officinale L. in response to multiple heavy metal stress. Ecotoxicol. Environ. Saf. 2024, 277, 116361. [Google Scholar] [CrossRef]
- Wang, X.-H.; Luo, W.-W.; Wang, Q.; He, L.-Y.; Sheng, X.-F. Metal(loid)-resistant bacteria reduce wheat Cd and As uptake in metal(loid)-contaminated soil. Environ. Pollut. 2018, 241, 529–539. [Google Scholar] [CrossRef]
- Sharma, P.; Tripathi, S.; Sirohi, R.; Kim, S.H.; Ngo, H.H.; Pandey, A. Uptake and mobilization of heavy metals through phytoremediation process from native plants species growing on complex pollutants: Antioxidant enzymes and photosynthetic pigments response. Environ. Technol. Innov. 2021, 23, 101629. [Google Scholar] [CrossRef]
- Parwez, R.; Aftab, T.; Khan, M.M.A.; Naeem, M. Exogenous abscisic acid fine-tunes heavy metal accumulation and plant’s antioxidant defence mechanism to optimize crop performance and secondary metabolite production in Trigonella foenum-graecum L. under nickel stress. Plant Sci. 2023, 332, 111703. [Google Scholar] [CrossRef]
- Romero-Puertas, M.C.; Terrón-Camero, L.C.; Peláez-Vico, M.Á.; Olmedilla, A.; Sandalio, L.M. Reactive oxygen and nitrogen species as key indicators of plant responses to Cd stress. Environ. Exp. Bot. 2019, 161, 107–119. [Google Scholar] [CrossRef]
- Berni, R.; Luyckx, M.; Xu, X.; Legay, S.; Sergeant, K.; Hausman, J.-F.; Lutts, S.; Cai, G.; Guerriero, G. Reactive oxygen species and heavy metal stress in plants: Impact on the cell wall and secondary metabolism. Environ. Exp. Bot. 2019, 161, 98–106. [Google Scholar] [CrossRef]
- Khan, Z.; Fan, X.; Khan, M.N.; Khan, M.A.; Zhang, K.; Fu, Y.; Shen, H. The toxicity of heavy metals and plant signaling facilitated by biochar application: Implications for stress mitigation and crop production. Chemosphere 2022, 308, 136466. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Ugbaja, R.N.; Akinhanmi, T.F.; Onunkwor, B.O.; Ugwor, E.I.; James, A.S.; Babalola, A.A.; Babayemi, D.O.; Ezenandu, E.O.; Ugbaja, V.C. Flavonoid-rich fractions of C. volubile and V. amygdalina alleviates arsenic-induced neurotoxicity by improving neurosignaling and antioxidant capacity in rats’ brain. Brain Disord. 2022, 7, 100050. [Google Scholar] [CrossRef]
- Li, Q.; Sun, W.; Chen, C.; Dong, D.; Cao, Y.; Dong, Y.; Yu, L.; Yue, Z.; Jin, X. Overexpression of histone demethylase gene SlJMJ524 from tomato confers Cd tolerance by regulating metal transport-related protein genes and flavonoid content in Arabidopsis. Plant Sci. 2022, 318, 111205. [Google Scholar] [CrossRef] [PubMed]
- Kazemitabar, S.K.; Faraji, S.; Najafi-Zarrini, H. Identification and in silico evaluation of bHLH genes in the Sesamum indicum genome: Growth regulation and stress dealing specially through the metal ions homeostasis and flavonoid biosynthesis. Gene Rep. 2020, 19, 100639. [Google Scholar] [CrossRef]
- Wang, B.; Wu, C.; Wang, G.; He, J.; Zhu, S. Transcriptomic analysis reveals a role of phenylpropanoid pathway in the enhancement of chilling tolerance by pre-storage cold acclimation in cucumber fruit. Sci. Hortic. 2021, 288, 110282. [Google Scholar] [CrossRef]
- Yu, M.; Zhuo, R.; Lu, Z.; Li, S.; Chen, J.; Wang, Y.; Li, J.; Han, X. Molecular insights into lignin biosynthesis on cadmium tolerance: Morphology, transcriptome and proteome profiling in Salix matsudana. J. Hazard. Mater. 2023, 441, 129909. [Google Scholar] [CrossRef]
- Zhao, T.; Zhang, K.; Chen, J.; Shi, X.; Li, X.; Ma, Y.; Fang, G.; Xu, S. Changes in heavy metal mobility and availability in contaminated wet-land soil remediated using lignin-based poly(acrylic acid). J. Hazard. Mater. 2019, 368, 459–467. [Google Scholar] [CrossRef]
- Du, B.; Li, W.; Zhu, H.; Xu, J.; Wang, Q.; Shou, X.; Wang, X.; Zhou, J. A functional lignin for heavy metal ions adsorption and wound care dressing. Int. J. Biol. Macromol. 2023, 239, 124268. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, S.; Shan, X.; Mu, H. Phyto-availability and speciation change of heavy metals in soils amended with lignin as micro-fertilizer. Sci. China Ser. C Life Sci. 2005, 48, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Ur Rahman, S.; Li, Y.; Hussain, S.; Hussain, B.; Khan, W.-u.-D.; Riaz, L.; Nadeem Ashraf, M.; Athar Khaliq, M.; Du, Z.; Cheng, H. Role of phytohormones in heavy metal tolerance in plants: A review. Ecol. Indic. 2023, 146, 109844. [Google Scholar] [CrossRef]
- Meng, Y.; Huang, J.; Jing, H.; Wu, Q.; Shen, R.; Zhu, X. Exogenous abscisic acid alleviates Cd toxicity in Arabidopsis thaliana by inhibiting Cd uptake, translocation and accumulation, and promoting Cd chelation and efflux. Plant Sci. 2022, 325, 111464. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.T.; Kao, C.H. Role of abscisic acid in cadmium tolerance of rice (Oryza sativa L.) seedlings. Plant Cell Environ. 2003, 26, 867–874. [Google Scholar] [CrossRef]
- Pan, W.; You, Y.; Shentu, J.-L.; Weng, Y.-N.; Wang, S.-T.; Xu, Q.-R.; Liu, H.-J.; Du, S.-T. Abscisic acid (ABA)-importing transporter 1 (AIT1) contributes to the inhibition of Cd accumulation via exogenous ABA application in Arabidopsis. J. Hazard. Mater. 2020, 391, 122189. [Google Scholar] [CrossRef] [PubMed]
- Baharudin, N.F.; Osman, N.I. Plant development, stress responses, and secondary metabolism under ethylene regulation. Plant Stress 2023, 7, 100146. [Google Scholar] [CrossRef]
- Bakshe, P.; Jugade, R. Phytostabilization and rhizofiltration of toxic heavy metals by heavy metal accumulator plants for sustainable management of contaminated industrial sites: A comprehensive review. J. Hazard. Mater. Adv. 2023, 10, 100293. [Google Scholar] [CrossRef]
- Ravanbakhsh, M.; Kowalchuk, G.A.; Jousset, A. Optimization of plant hormonal balance by microorganisms prevents plant heavy metal accumulation. J. Hazard. Mater. 2019, 379, 120787. [Google Scholar] [CrossRef]
- Guan, C.; Ji, J.; Li, X.; Jin, C.; Wang, G. LcMKK, a MAPK kinase from Lycium chinense, confers cadmium tolerance in transgenic tobacco by transcriptional upregulation of ethylene responsive transcription factor gene. J. Genet. 2016, 95, 875–885. [Google Scholar] [CrossRef]
- Feng, Y.; Yang, J.; Liu, W.; Yan, Y.; Wang, Y. Hydroxyapatite as a passivator for safe wheat production and its impacts on soil microbial communities in a Cd-contaminated alkaline soil. J. Hazard. Mater. 2021, 404, 124005. [Google Scholar] [CrossRef] [PubMed]
- Shyla, B.; Mahadevaiah; Nagendrappa, G. A simple spectrophotometric method for the determination of phosphate in soil, detergents, water, bone and food samples through the formation of phosphomolybdate complex followed by its reduction with thiourea. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2011, 78, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, Z.; Zhao, F.; Yu, L.; Yang, W.; Si, M.; Liao, Q. Organic acid, phosphate, sulfate and ammonium co-metabolism releasing insoluble phosphate by Klebsiella aerogenes to simultaneously stabilize lead and cadmium. J. Hazard. Mater. 2023, 443, 130378. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Cai, H.; Wang, X.; Hu, X.; Chen, Z.; Yao, L. Heavy metal-immobilizing bacteria increase the biomass and reduce the Cd and Pb uptake by pakchoi (Brassica chinensis L.) in heavy metal-contaminated soil. Ecotoxicol. Environ. Saf. 2020, 195, 110375. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; He, L.-y.; Wang, Q.; Sheng, X.-f. Synergistic effects of plant growth-promoting Neorhizobium huautlense T1-17 and immobilizers on the growth and heavy metal accumulation of edible tissues of hot pepper. J. Hazard. Mater. 2016, 312, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Ren, Y.; Yu, G.; Shi, C.; Liu, L.; Guo, Q.; Han, C.; Zhang, D.; Zhang, L.; Liu, B.; Gao, H.; et al. Majorbio Cloud: A one-stop, comprehensive bioinformatic platform for multiomics analyses. iMeta 2022, 1, e12. [Google Scholar] [CrossRef]
- Fisher, K.A.; Yarwood, S.A.; James, B.R. Soil urease activity and bacterial ureC gene copy numbers: Effect of pH. Geoderma 2017, 285, 1–8. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kan, D.; Tian, M.; Ruan, Y.; Han, H. Phosphorus-Solubilizing Bacteria Enhance Cadmium Immobilization and Gene Expression in Wheat Roots to Reduce Cadmium Uptake. Plants 2024, 13, 1989. https://doi.org/10.3390/plants13141989
Kan D, Tian M, Ruan Y, Han H. Phosphorus-Solubilizing Bacteria Enhance Cadmium Immobilization and Gene Expression in Wheat Roots to Reduce Cadmium Uptake. Plants. 2024; 13(14):1989. https://doi.org/10.3390/plants13141989
Chicago/Turabian StyleKan, Delong, Minyu Tian, Ying Ruan, and Hui Han. 2024. "Phosphorus-Solubilizing Bacteria Enhance Cadmium Immobilization and Gene Expression in Wheat Roots to Reduce Cadmium Uptake" Plants 13, no. 14: 1989. https://doi.org/10.3390/plants13141989
APA StyleKan, D., Tian, M., Ruan, Y., & Han, H. (2024). Phosphorus-Solubilizing Bacteria Enhance Cadmium Immobilization and Gene Expression in Wheat Roots to Reduce Cadmium Uptake. Plants, 13(14), 1989. https://doi.org/10.3390/plants13141989