

Application and Evaluation of the Antifungal Activities of Glandular Trichome Secretions from Air/Sun-Cured Tobacco Germplasms against Botrytis cinerea
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Assays of Tobacco Glandular Trichome Extract Fractions
2.1.1. UPLC-Q-TOF MS Assay
2.1.2. GC-MS Assay
2.2. Identification of Antifungal Activity in the Tobacco Glandular Trichome Extracts
2.2.1. Activity Assay of Glandular Trichome Extracts from Four Different Parts of the Tobacco Plant
2.2.2. Identification of Antifungal Activities among the 80 Glandular Trichome Extracts
2.3. OPLS Model Analysis of Antimicrobial Activity and Secretion Components
3. Discussion
3.1. Tobacco Glandular Trichomes
3.2. Excellent Tobacco Resources Evaluated Based on Antifungal Activity
3.3. Mining and Utilization of Antimicrobial Substances
4. Conclusions
5. Materials and Methods
5.1. Materials and Reagents
5.2. Methods
5.2.1. Planting of Germplasm Resources
5.2.2. Extraction of Glandular Trichome Secretions
5.2.3. Detection Assays
5.2.4. Antifungal Activity
5.3. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Peak1 | Peak2 | Peak3 | Peak4 | Peak5 | Peak6 | Peak7 | Peak8 | Peak9 | Peak10 | Peak11 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| X1 | 48.2 | 98.4 | 90.6 | 104.3 | 238.3 | 277.4 | 2022.2 | 1457.3 | 302.4 | 932.0 | 20.6 |
| X2 | 10.6 | 12.3 | 0.0 | 234.1 | 182.6 | 189.0 | 1792.6 | 1371.0 | 171.0 | 614.1 | 113.8 |
| X3 | 59.4 | 60.1 | 88.9 | 77.6 | 126.2 | 152.6 | 1655.5 | 1203.4 | 376.1 | 545.2 | 154.1 |
| X4 | 89.4 | 40.0 | 60.6 | 85.3 | 135.2 | 148.0 | 2269.6 | 1632.7 | 288.0 | 20.1 | 33.4 |
| X5 | 41.8 | 47.2 | 43.5 | 195.5 | 116.4 | 134.1 | 1842.5 | 1462.7 | 401.9 | 763.2 | 220.7 |
| X6 | 89.2 | 122.6 | 104.5 | 128.3 | 225.3 | 211.9 | 1384.4 | 1134.6 | 242.2 | 569.0 | 172.3 |
| X7 | 13.7 | 106.8 | 108.3 | 108.1 | 235.9 | 188.8 | 1666.8 | 1322.4 | 329.3 | 570.7 | 253.8 |
| X8 | 38.8 | 45.9 | 48.3 | 142.7 | 89.6 | 112.9 | 1009.8 | 803.5 | 428.2 | 425.4 | 81.0 |
| X9 | 39.4 | 40.2 | 27.0 | 152.3 | 141.3 | 112.4 | 598.4 | 679.1 | 206.2 | 855.4 | 59.9 |
| X10 | 33.2 | 33.7 | 41.4 | 132.7 | 75.0 | 87.8 | 671.9 | 528.9 | 146.8 | 584.1 | 23.7 |
| X11 | 6.1 | 55.3 | 87.0 | 166.4 | 162.0 | 191.9 | 1522.9 | 1218.8 | 418.8 | 898.8 | 154.3 |
| X12 | 82.9 | 82.5 | 64.6 | 158.3 | 167.6 | 127.0 | 1523.4 | 1078.3 | 518.2 | 11.7 | 0.0 |
| X13 | 17.3 | 15.4 | 39.2 | 158.4 | 133.3 | 150.5 | 824.7 | 689.0 | 154.9 | 595.0 | 81.9 |
| X14 | 27.8 | 33.0 | 44.3 | 155.0 | 105.2 | 100.5 | 991.4 | 898.3 | 161.1 | 677.6 | 67.9 |
| X15 | 26.1 | 11.8 | 0.0 | 185.1 | 84.8 | 98.2 | 1045.1 | 866.3 | 250.4 | 474.9 | 49.6 |
| X16 | 50.6 | 60.6 | 57.1 | 127.8 | 98.0 | 99.8 | 657.1 | 568.5 | 362.1 | 750.1 | 8.7 |
| X17 | 41.5 | 68.0 | 78.6 | 146.5 | 133.7 | 172.1 | 1162.6 | 866.5 | 479.2 | 710.7 | 48.1 |
| X18 | 69.4 | 71.7 | 82.8 | 127.7 | 162.3 | 184.0 | 971.6 | 790.1 | 433.5 | 680.3 | 107.1 |
| X19 | 35.5 | 71.9 | 85.7 | 157.3 | 142.1 | 139.8 | 909.9 | 727.2 | 664.7 | 746.8 | 0.0 |
| X20 | 24.4 | 28.3 | 27.3 | 130.4 | 75.6 | 80.6 | 789.3 | 597.1 | 403.9 | 572.9 | 0.0 |
| X21 | 31.6 | 40.2 | 28.6 | 156.6 | 96.6 | 127.2 | 1019.4 | 786.3 | 354.6 | 680.7 | 26.0 |
| X22 | 39.9 | 58.0 | 88.3 | 171.1 | 132.6 | 138.8 | 1221.9 | 911.4 | 545.8 | 661.0 | 71.8 |
| X23 | 111.2 | 84.4 | 87.2 | 190.8 | 174.4 | 214.6 | 2075.4 | 1543.3 | 517.1 | 0.0 | 190.2 |
| X24 | 110.2 | 84.7 | 89.2 | 258.0 | 148.2 | 178.6 | 1596.8 | 1186.2 | 372.0 | 377.1 | 209.5 |
| X25 | 37.8 | 28.7 | 0.0 | 241.5 | 114.0 | 137.0 | 1669.0 | 1314.2 | 399.6 | 389.5 | 34.0 |
| X26 | 68.7 | 31.3 | 49.4 | 177.8 | 171.5 | 146.7 | 1632.4 | 1308.4 | 406.3 | 526.7 | 11.5 |
| X27 | 69.3 | 62.8 | 85.2 | 199.7 | 114.1 | 100.4 | 1784.4 | 1322.0 | 471.1 | 356.2 | 30.9 |
| X28 | 186.5 | 116.8 | 118.4 | 280.3 | 188.5 | 188.5 | 2339.3 | 1648.8 | 425.5 | 5.1 | 0.0 |
| X29 | 233.2 | 164.6 | 147.7 | 284.4 | 264.3 | 222.4 | 2786.3 | 2255.7 | 598.3 | 0.0 | 104.6 |
| X30 | 58.6 | 64.4 | 94.6 | 191.6 | 132.2 | 147.5 | 2004.2 | 1506.9 | 487.7 | 214.4 | 191.6 |
| X31 | 69.5 | 77.3 | 96.5 | 159.2 | 137.2 | 126.0 | 1887.1 | 1426.2 | 322.7 | 4.8 | 93.2 |
| X32 | 32.3 | 62.2 | 63.3 | 139.2 | 99.3 | 92.6 | 1845.1 | 1300.6 | 484.0 | 865.5 | 89.7 |
| X33 | 200.9 | 109.8 | 78.5 | 145.6 | 157.8 | 164.4 | 1013.4 | 637.5 | 444.5 | 741.1 | 53.0 |
| X34 | 40.6 | 68.3 | 75.4 | 145.0 | 104.9 | 100.2 | 1800.3 | 1346.6 | 358.6 | 406.1 | 120.5 |
| X35 | 114.0 | 87.1 | 76.4 | 158.1 | 102.6 | 94.8 | 1317.4 | 899.3 | 558.9 | 201.0 | 5.1 |
| X36 | 0.0 | 0.0 | 16.1 | 5.7 | 5.8 | 11.1 | 123.1 | 92.8 | 170.7 | 0.0 | 0.0 |
| X37 | 103.3 | 117.5 | 96.7 | 147.3 | 180.8 | 143.1 | 1575.7 | 1101.2 | 364.2 | 624.7 | 86.1 |
| X38 | 91.6 | 75.7 | 73.8 | 129.6 | 92.3 | 99.3 | 1058.9 | 775.0 | 315.3 | 645.4 | 23.9 |
| X39 | 158.8 | 121.8 | 108.2 | 215.3 | 145.3 | 162.7 | 1915.4 | 1260.3 | 391.0 | 7.4 | 11.6 |
| X40 | 160.2 | 487.0 | 179.9 | 245.1 | 323.3 | 322.6 | 2138.4 | 1712.4 | 434.7 | 909.9 | 126.8 |
| X41 | 219.8 | 152.4 | 144.9 | 265.8 | 220.6 | 234.8 | 2577.6 | 1801.5 | 590.5 | 703.2 | 224.9 |
| X42 | 115.2 | 99.1 | 113.6 | 221.6 | 177.8 | 186.4 | 1775.1 | 1314.5 | 432.8 | 416.7 | 40.2 |
| X43 | 294.4 | 69.5 | 0.0 | 270.9 | 132.0 | 140.9 | 1950.4 | 1120.0 | 355.2 | 949.8 | 409.7 |
| X44 | 155.1 | 139.1 | 140.8 | 313.1 | 258.7 | 240.3 | 3027.8 | 2091.3 | 588.8 | 14.0 | 6.6 |
| X45 | 182.9 | 119.6 | 114.2 | 214.5 | 181.2 | 194.8 | 1822.4 | 1369.1 | 441.4 | 441.5 | 82.9 |
| X46 | 95.0 | 64.9 | 106.3 | 194.3 | 175.5 | 167.2 | 2299.4 | 1591.6 | 421.1 | 580.4 | 144.5 |
| X47 | 69.0 | 57.6 | 124.4 | 234.4 | 235.5 | 231.5 | 2544.6 | 1966.8 | 444.8 | 842.1 | 97.2 |
| X48 | 128.4 | 219.4 | 194.5 | 352.1 | 369.6 | 353.1 | 3740.9 | 2732.2 | 557.3 | 3.2 | 55.1 |
| X49 | 56.7 | 35.5 | 57.4 | 91.2 | 82.8 | 85.2 | 577.0 | 826.0 | 287.6 | 283.7 | 18.4 |
| X50 | 47.7 | 60.5 | 62.1 | 152.3 | 75.9 | 82.0 | 691.1 | 420.7 | 417.3 | 169.0 | 36.8 |
| X51 | 62.8 | 81.2 | 125.7 | 281.1 | 243.8 | 228.8 | 2900.4 | 1997.8 | 432.8 | 4.9 | 10.8 |
| X52 | 31.1 | 75.9 | 105.1 | 207.2 | 219.6 | 189.9 | 2335.3 | 1857.1 | 403.1 | 4.7 | 168.3 |
| X53 | 205.7 | 146.1 | 142.5 | 284.0 | 261.3 | 236.1 | 2464.7 | 1710.6 | 457.4 | 58.8 | 13.5 |
| X54 | 86.1 | 144.3 | 192.3 | 307.9 | 324.7 | 342.0 | 2605.1 | 2100.9 | 500.0 | 4.0 | 309.3 |
| X55 | 178.7 | 117.6 | 85.4 | 181.7 | 133.8 | 140.0 | 1788.5 | 1062.6 | 395.6 | 336.0 | 75.5 |
| X56 | 102.1 | 141.3 | 93.5 | 124.1 | 169.3 | 183.2 | 1358.3 | 916.1 | 371.1 | 352.1 | 19.8 |
| X57 | 140.0 | 99.5 | 90.9 | 184.6 | 141.2 | 156.6 | 1682.9 | 1224.3 | 413.5 | 417.2 | 32.2 |
| X58 | 288.9 | 194.9 | 174.9 | 361.7 | 302.5 | 280.0 | 2878.6 | 2092.7 | 495.6 | 4.6 | 22.4 |
| X59 | 208.1 | 132.9 | 124.7 | 275.4 | 190.8 | 217.1 | 2240.6 | 1628.3 | 383.0 | 0.0 | 22.0 |
| X60 | 144.4 | 47.8 | 23.9 | 160.5 | 104.5 | 117.2 | 1798.8 | 804.2 | 296.2 | 421.9 | 946.3 |
| X61 | 144.8 | 95.3 | 80.6 | 118.0 | 155.9 | 158.4 | 1591.9 | 1237.8 | 555.7 | 745.3 | 90.5 |
| X62 | 70.6 | 74.5 | 43.2 | 189.1 | 147.9 | 158.2 | 1897.4 | 879.5 | 335.2 | 477.5 | 917.7 |
| X63 | 197.9 | 174.6 | 148.0 | 243.6 | 237.7 | 222.6 | 2852.5 | 2247.4 | 522.3 | 812.9 | 339.2 |
| X64 | 71.2 | 50.6 | 72.6 | 126.0 | 110.4 | 131.9 | 700.9 | 552.3 | 416.7 | 751.0 | 56.7 |
| X65 | 50.2 | 30.7 | 52.2 | 94.9 | 72.5 | 73.8 | 641.5 | 517.0 | 375.2 | 679.0 | 56.4 |
| X66 | 68.4 | 111.9 | 130.2 | 246.0 | 198.2 | 252.6 | 1697.5 | 1230.1 | 396.9 | 740.3 | 81.6 |
| X67 | 79.8 | 107.0 | 109.7 | 230.9 | 191.2 | 199.9 | 1588.8 | 1251.7 | 393.0 | 29.1 | 60.8 |
| X68 | 58.7 | 39.9 | 75.5 | 142.4 | 96.7 | 93.1 | 984.4 | 771.7 | 453.1 | 441.9 | 44.5 |
| X69 | 108.5 | 83.4 | 104.6 | 141.4 | 189.0 | 205.1 | 1142.8 | 826.1 | 445.2 | 880.9 | 87.1 |
| X70 | 192.6 | 140.2 | 131.2 | 241.7 | 204.4 | 181.2 | 2371.7 | 1864.3 | 522.6 | 644.5 | 274.8 |
| X71 | 56.5 | 98.3 | 117.5 | 247.0 | 164.6 | 180.8 | 1946.7 | 1445.5 | 397.8 | 430.4 | 111.2 |
| X72 | 126.1 | 99.5 | 125.6 | 225.0 | 199.1 | 169.3 | 2506.5 | 1956.7 | 399.7 | 678.4 | 290.9 |
| X73 | 158.3 | 115.1 | 139.3 | 242.5 | 230.0 | 202.6 | 2704.7 | 2067.6 | 521.9 | 609.1 | 363.4 |
| X74 | 80.3 | 58.6 | 90.3 | 135.5 | 128.1 | 140.7 | 1006.4 | 753.9 | 474.5 | 712.5 | 60.9 |
| X75 | 86.5 | 56.0 | 93.0 | 182.7 | 162.7 | 158.6 | 1846.3 | 1371.7 | 398.0 | 602.9 | 97.5 |
| X76 | 18.1 | 25.8 | 54.9 | 119.8 | 76.1 | 83.6 | 951.2 | 638.7 | 351.5 | 487.3 | 34.4 |
| X77 | 182.2 | 146.0 | 116.6 | 177.4 | 189.8 | 179.1 | 1665.1 | 1102.1 | 439.4 | 529.2 | 47.5 |
| X78 | 60.4 | 54.4 | 106.5 | 208.5 | 160.8 | 170.4 | 2031.3 | 1534.4 | 525.8 | 518.4 | 94.7 |
| X79 | 79.7 | 136.3 | 119.6 | 25.6 | 137.7 | 156.4 | 1486.1 | 981.1 | 407.5 | 426.6 | 0.0 |
| X80 | 115.4 | 146.6 | 146.6 | 40.3 | 148.6 | 152.9 | 1193.3 | 807.8 | 363.8 | 336.6 | 12.2 |
| No. | SE(I) | SE(II) | SE(III) | SE(IV) | SE(V) | NO | SE(I) | SE(II) | SE(III) | SE(IV) | SE(V) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| X1 | 290.2 | 419.7 | 768 | 823.1 | 337.1 | X41 | 200.3 | 358 | 874.6 | 1121.2 | 823.1 |
| X2 | 319.9 | 584.9 | 1156.7 | 1578.9 | 770 | X42 | 134.8 | 295.9 | 731 | 1018 | 835.1 |
| X3 | 240.1 | 302.7 | 637.2 | 1319.8 | 910 | X43 | 21.2 | 53 | 80.5 | 33.7 | 9.9 |
| X4 | 397.4 | 893.5 | 146.5 | 64.5 | 25.8 | X44 | 75.3 | 329.5 | 703.8 | 1141.8 | 918.8 |
| X5 | 237.8 | 403.6 | 747.3 | 1368.6 | 583.2 | X45 | 134.7 | 293.9 | 778.7 | 1094.3 | 970.5 |
| X6 | 186.4 | 280.5 | 726.4 | 1184.2 | 660.5 | X46 | 130.5 | 245.4 | 622.6 | 1128.3 | 1373.2 |
| X7 | 132.1 | 208.3 | 586.8 | 802.1 | 241.3 | X47 | 249.3 | 415.6 | 967.8 | 1390.1 | 1245.1 |
| X8 | 134 | 217.9 | 418.9 | 499.5 | 224.9 | X48 | 145.1 | 346.9 | 757.7 | 997.5 | 1193.2 |
| X9 | 164 | 301.1 | 778.9 | 1206.7 | 923.6 | X49 | 156.3 | 311.5 | 731 | 1047.6 | 778.7 |
| X10 | 111.5 | 234.7 | 572.5 | 718.3 | 266.7 | X50 | 0 | 73.1 | 121.2 | 29.2 | 32.9 |
| X11 | 150.4 | 292.8 | 723.2 | 962.8 | 537.9 | X51 | 135.8 | 283.3 | 781.3 | 1141.3 | 1371.7 |
| X12 | 134.7 | 228.3 | 568.8 | 869.6 | 433.7 | X52 | 302.9 | 635.1 | 166.1 | 76.7 | 15.9 |
| X13 | 163.5 | 282.6 | 629.5 | 790.2 | 288.2 | X53 | 156.4 | 233 | 469.9 | 744.1 | 1024.3 |
| X14 | 172 | 345.8 | 809.3 | 1678.7 | 528 | X54 | 273.4 | 657.8 | 83.5 | 63.7 | 28.2 |
| X15 | 122.3 | 199.9 | 388.3 | 480.4 | 246.1 | X55 | 108.5 | 168.3 | 384.8 | 584.9 | 675.5 |
| X16 | 150.6 | 169.7 | 366 | 344.4 | 272.9 | X56 | 176.6 | 181.4 | 350 | 508 | 525 |
| X17 | 141.8 | 287.9 | 556.1 | 621.1 | 281.4 | X57 | 83.2 | 191.2 | 573.4 | 1035.5 | 1116.4 |
| X18 | 212.2 | 287.3 | 674.7 | 815.8 | 267.8 | X58 | 153.2 | 308.3 | 795.8 | 1236 | 966 |
| X19 | 207.7 | 316.3 | 555.5 | 586.5 | 255.8 | X59 | 268.6 | 782.1 | 132.1 | 94.1 | 54.3 |
| X20 | 130.1 | 258.7 | 544.8 | 688.6 | 287.3 | X60 | 143.3 | 286 | 609.6 | 811 | 690.7 |
| X21 | 224.2 | 368.3 | 655.9 | 714 | 336 | X61 | 251.8 | 326 | 717.7 | 909.8 | 628.8 |
| X22 | 188.5 | 364.3 | 717.5 | 849.2 | 159.1 | X62 | 192.5 | 327 | 733.9 | 922.8 | 769.4 |
| X23 | 286.2 | 603.1 | 115.9 | 91.9 | 22.2 | X63 | 156.6 | 320.9 | 822.5 | 1133.2 | 800.3 |
| X24 | 411.1 | 990.3 | 175.9 | 72 | 17.1 | X64 | 180.3 | 271.8 | 712 | 826.9 | 328.9 |
| X25 | 248.7 | 643 | 119.5 | 45.1 | 14 | X65 | 175 | 283.7 | 569.7 | 648.3 | 438.4 |
| X26 | 158.6 | 356.6 | 881.7 | 1304.9 | 687.4 | X66 | 278.6 | 657.3 | 378.5 | 420.8 | 289.4 |
| X27 | 50.3 | 252.8 | 627.1 | 941.9 | 694.1 | X67 | 227.2 | 467.3 | 966.1 | 914.8 | 292.7 |
| X28 | 208.1 | 622.2 | 83.2 | 50 | 13.7 | X68 | 171.6 | 318.2 | 588.6 | 689.7 | 324.6 |
| X29 | 130.6 | 307.3 | 802.4 | 1079 | 1245.2 | X69 | 224.9 | 352 | 679.1 | 750.2 | 338.1 |
| X30 | 184.8 | 475.5 | 396.3 | 476.1 | 263.8 | X70 | 188.3 | 456.2 | 828.6 | 1190.4 | 751.2 |
| X31 | 140.5 | 375.6 | 537.4 | 843.6 | 430.6 | X71 | 276.2 | 657.2 | 182.5 | 75.2 | 29.9 |
| X32 | 152.1 | 264 | 544.8 | 784.2 | 753 | X72 | 96.3 | 250.9 | 421.1 | 626.8 | 455.3 |
| X33 | 204 | 311.6 | 578.3 | 738.5 | 360.7 | X73 | 203 | 417.3 | 953 | 1131 | 754.9 |
| X34 | 141 | 281.3 | 559.7 | 748.8 | 571.3 | X74 | 222.3 | 391.3 | 718.5 | 746.9 | 361.6 |
| X35 | 123.4 | 135.7 | 317.5 | 546 | 473.1 | X75 | 205.4 | 463.9 | 1049.6 | 1450.3 | 765.2 |
| X36 | 149.9 | 254.8 | 145.9 | 72.2 | 35.3 | X76 | 155.6 | 246.2 | 584 | 963.9 | 619.7 |
| X37 | 128.1 | 191.3 | 690.1 | 829.9 | 229.5 | X77 | 145.1 | 211.1 | 492 | 651.2 | 511.3 |
| X38 | 91.3 | 276.1 | 844.2 | 1483.9 | 654.1 | X78 | 203.3 | 387.8 | 886.2 | 1232.7 | 814.1 |
| X39 | 89.7 | 184.3 | 563.6 | 900.6 | 1234.7 | X79 | 59.5 | 138.4 | 358.7 | 460.2 | 314.8 |
| X40 | 213.2 | 301.1 | 748.3 | 1072.4 | 977.4 | X80 | 202.5 | 237.8 | 447.2 | 593.3 | 462.6 |
References
- Fillinger, S.; Elad, Y. Botrytis-the Fungus, the Pathogen and Its Management in Agricultural Systems, Ripening of Tomato Fruit and Susceptibility to Botrytis Cinerea; Springer International Publishing: Cham, Switzerland, 2016; pp. 387–412. [Google Scholar]
- Bui, T.T.; Wright, S.A.; Falk, A.B.; Vanwalleghem, T.; Van, H.W.; Hertog, M.L.; Keulemans, J.; Davey, M.W. Botrytis cinerea differentially induces postharvest antioxidant responses in ‘Braeburn’ and ‘Golden Delicious’ apple fruit. J. Sci. Food Agric. 2019, 99, 5662–5670. [Google Scholar] [CrossRef]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Kan, L.J.A.L.V. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef]
- Gian, P.R.; Joseph, L.; Smilanick, E.F.; Samir, D. Integrated management of postharvest gray mold on fruit crops. Postharvest Biol. Technol. 2016, 113, 69–76. [Google Scholar] [CrossRef]
- Li, H.; Chen, Y.; Zhang, Z.Q.; Li, B.Q.; Qin, G.Z.; Tian, S.P. Pathogenic mechanisms and control strategies of Botrytis cinerea causing post-harvest decay in fruits and vegetables. Food Qual. Saf. 2018, 2, 111–119. [Google Scholar] [CrossRef]
- Leroch, M.; Plesken, C.; Weber, R.W.S.; Kauff, F.; Scalliet, G.; Hahn, M. Gray mold populations in german strawberry fields are resistant to multiple fungicides and dominated by a novel clade closely related to Botrytis cinerea. Appl. Environ. Microb. 2013, 79, 159–167. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Zhang, J.; Wang, X.D.; Yang, L.; Jiang, D.H.; Li, G.Q.; Hsiang, T.; Zhuang, W.Y. Morphological and phylogenetic identification of Botrytis sinoviticola, a novel cryptic species causing gray mold disease of table grapes (Vitis vinifera) in China. Mycologia 2013, 106, 43–56. [Google Scholar] [CrossRef]
- Johnston, P.R.; Hoksbergen, K.; Park, D.; Beever, R.E. Genetic diversity of Botrytis in New Zealand vineyards and the significance of its seasonal and regional variation. Plant Pathol. 2014, 63, 888–898. [Google Scholar] [CrossRef]
- Saito, S.; Margosan, D.; Michailides, T.J.; Xiao, C.L. Botrytis California, a new cryptic species in the B. cinerea species complex causing gray mold in blueberries and table grapes in California. Mycologia 2016, 108, 330–343. [Google Scholar] [CrossRef]
- Garfinkel, A.R.; Lorenzini, M.; Zapparoli, G.; Chastagner, G.A. Botrytis euroamericana, a new species from peony and grape in North America and Europe. Mycologia 2017, 109, 495–507. [Google Scholar] [CrossRef]
- Shao, W.; Zhao, Y.; Ma, Z. Advances in Understanding Fungicide Resistance in Botrytis cinerea in China. Phytopathology 2021, 111, 455–463. [Google Scholar] [CrossRef]
- Xu, H.X.; Zheng, X.S.; Tian, J.; Lai, F.X.; He, J.C.; Lu, Z.X. Advances in the development and application of control technologies for insect pest management in paddy fields in China. J. Plant Prot. 2017, 44, 925–939. [Google Scholar]
- Cordova-Albores, L.C.; Rios, M.Y.; Barrera-Necha, L.L.; Bautista-Baños, S. Chemical compounds of a native Jatropha curcas seed oil from Mexico and their antifungal effect on Fusarium oxysporum f. sp. gladioli. Ind. Crops Prod. 2014, 62, 166–172. [Google Scholar] [CrossRef]
- Khaledi, N.; Taheri, P.; Tarighi, S. Antifungal activity of various essential oils against Rhizoctonia solani and Macrophomina phaseolina as major bean pathogens. J. Appl. Microbiol. 2015, 118, 704–717. [Google Scholar] [CrossRef]
- Jasso de Rodríguez, D.; Trejo-González, F.A.; Rodríguez-Garcia, R.; Díaz-Jimenez, M.L.V.; Sáenz-Galindo, A.; Hernández-Castillo, F.D.; Villarreal-Quintanilla, J.A.; Peña-Ramos, F.M. Antifungal activity in vitro of Rhus muelleri against Fusarium oxysporum f. sp. Lycopersici. Ind. Crops Prod. 2015, 75, 150–158. [Google Scholar] [CrossRef]
- Jasso de Rodríguez, D.J.; Hernández-Castillo, D.; Rodríguez-García, R.; Angulo-Sánchez, J.L. Antifungal activity in vitro of Aloe vera pulp and liquid fraction against plant pathogenic fungi. Ind. Crops Prod. 2015, 21, 81–87. [Google Scholar] [CrossRef]
- Ding, Y.L.; Sun, T.J.; Ao, K.; Peng, Y.J.; Zhang, Y.X.; Li, X.; Zhang, Y.L. Opposite roles of salicylic acid receptors Npr1 and Npr3/Npr4 in transcriptional regulation of plant immunity. Cell 2018, 173, 1454–1467. [Google Scholar] [CrossRef]
- Šernaitė, L.; Rasiukevičiūtė, N.; Valiuškaitė, A. Application of Plant Extracts to Control Postharvest Gray Mold and Susceptibility of Apple fruits to B. cinerea from Different Plant Hosts. Foods 2020, 9, 1430. [Google Scholar] [CrossRef]
- Chang, X.H.; Wang, Z.J.; Chen, Z.T.; Li, N. Effect of Preharvest Salicylic Acid Treatment on Postharvest Disease of Muskmelon and Disease Resistance-related Enzyme and Defense Compounds Induction. Storage Process 2019, 19, 51–56. [Google Scholar]
- Zhang, Y.; Li, X. Salicylic scid: Biosynthesis, perception, and contributions to plant immunity. Curr. Opin. Plant Biol. 2019, 50, 29–36. [Google Scholar] [CrossRef]
- Tremblay, R.; Wang, D.; Jevnikar, A.M.; Ma, S. Tobacco, a highly efficient green bioreactor for production of therapeutic proteins. Biotechnol. Adv. 2010, 28, 214–221. [Google Scholar] [CrossRef]
- Qu, Y.F.; Xu, M.L.; Cao, J.M.; Sun, H.Q.; You, X.W.; Wang, G.P.; Jiang, C.H.; Wang, Y.Y.; Chang, A.X. Differential analysis of major glandular trichome secretions of different tobacco genotypes. Acta Tabacaria Sin. 2018, 24, 45–52. [Google Scholar] [CrossRef]
- Roberto, B.; Wernsman, E.A. Trichome type, density and distribution on the leaves of certain tobacco varieties and hybrids. Tob. Sci. 1966, 10, 157–161. [Google Scholar]
- Jassbi, A.R.; Zare, S.; Asadollahi, M.; Schuman, M.C. Ecological Roles and Biological Activities of Specialized Metabolites from the Genus Nicotiana. Chem. Rev. 2017, 117, 12227–12280. [Google Scholar] [CrossRef]
- Wang, J.; Xu, K.; Zhang, J.H.; Ren, G.W.; Yang, X.Y.; Zhang, Z.F.; Zhang, Y.Y.; Xiao, Y.; Du, Y.M. Systematic activity-oriented separation and structure-activity relationship of tobacco cembranoids. Ind. Crops Prod. 2021, 173, 114136. [Google Scholar] [CrossRef]
- Yan, N.; Du, Y.M.; Liu, X.M.; Zhang, H.B.; Liu, Y.H.; Zhang, P.; Gong, D.P.; Zhang, Z.F. Chemical structures, biosynthesis, bioactivities, biocatalysis and semisynthesis of tobacco cembranoids: An overview. Ind. Crops Prod. 2016, 83, 66–80. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, J.; Zhang, P.; Xie, S.N.; Yuan, X.L.; Hou, X.D.; Yan, N.; Fang, Y.D.; Du, Y.M. In vitro and in vivo antifungal activity and preliminary mechanism of cembratrien-diols against Botrytis cinerea. Ind. Crops Prod. 2020, 154, 112745. [Google Scholar] [CrossRef]
- Mischko, W.; Hirte, M.; Roehrer, S.; Engelhardt, H.; Mehlmer, N.; Minceva, M.; Brück, T. Modular biomanufacturing for a sustainable production of terpenoid-based insect deterrents. Green Chem. 2018, 20, 2637–2650. [Google Scholar] [CrossRef]
- Yan, B.W.; Jia, H.J.; Wei, J.H.; Huang, C.J.; Huang, B.; Mao, M.B.; Wang, J.; Wang, J.; Huang, S. Extraction, purification and anti-TMV effects of α (β)-2,7,11-cembratriene-4,6-diol from tobacco leaves. Ind. Crops Prod. 2021, 174, 1141972021. [Google Scholar] [CrossRef]
- Seo, S.; Seto, H.; Koshino, H.; Yoshida, S.; Ohashi, Y. A Diterpene as an Endogenous Signal for the Activation of Defense Responses to Infection with Tobacco mosaic virus and wounding in Tobacco. Plant Cell 2003, 15, 863–873. [Google Scholar] [CrossRef]
- Carlos, C.G.; Inmaculada, V.; Philip, A.G.E.; Josep, U.; Neus, T. Suppression of Botrytis cinerea on nectrotic grapevine tissues by early season applications of natural products and biocontrol agents. Pest. Manag. Sci. 2014, 70, 595–602. [Google Scholar] [CrossRef]
- Ren, M.; Wang, Z.D.; Mu, J.M.; Liu, Y.H.; Zhang, X.W. Overview of species and distribution of tobacco germplasm resources in China. Chin. Tob. Sci. 2009, 30, 8–14. [Google Scholar]
- Yang, P.F.; Hua, T.; Huang, S.; Yang, J.; Wei, T.; Mao, D.B. Progress on Cembranoid Diterpenes in Tobacco. Huaxueshiji 2020, 42, 380–388. [Google Scholar]
- Cai, L.L.; Xie, F.W.; Liu, K.J.; Zhang, Y.; Xie, J.P. Analysis of Sucrose Esters in Oriental Tobacco by GC/MS. Tob. Sci. Technol. 2009, 42, 40–44. [Google Scholar]
- Duan, C.; Du, Y.M.; Hou, X.D.; Yan, N.; Dong, W.J.; Mao, X.X.; Zhang, Z.F. Chemical Basis of the Fungicidal Activity of Tobacco Extracts against Valsa mali. Molecules 2016, 21, 1743. [Google Scholar] [CrossRef]
- Yan, N.; Du, Y.M.; Liu, X.M.; Zhang, H.B.; Liu, Y.H.; Shi, J.H.; Xue, S.J.; Zhang, Z.F. Analyses of effects of α-cembratrien-diol on cell morphology and transcriptome of Valsa mali var. mali. Food Chem. 2017, 214, 110–118. [Google Scholar] [CrossRef]
- Han, J.F.; Zhang, Z.Y.; Liu, H.S.; Wang, X.J.; Liu, H.; Mao, G.J. Research advance in tobacco glandular trichomes and their secretion substance cembranoids. Acta Tabacaria Sin. 2013, 19, 118–124. [Google Scholar]
- Kennedy, B.S.; Nielsen, M.T.; Severson, R.F. Biorationals from Nicotiana protect cucumbers against Colletotrichum lagenarium (Pass.) ell. & halst disease development. J. Chem. Ecol. 1995, 21, 221–231. [Google Scholar] [CrossRef]
- Aqil, F.; Zahin, M.; Sayed, K.A.E. Antimicrobial, antioxidant, and antimutagenic activities of selected marine natural products and tobacco cembranoids. Drug Chem. Toxicol. 2011, 34, 167–179. [Google Scholar] [CrossRef]
- Seo, S.; Gomi, K.; Kaku, H.; Abe, H.; Seto, H.; Nakatsu, S.; Neya, M.; Kobayashi, M.; Nakaho, K.; Ichinose, Y.; et al. Identification of natural diterpenes that inhibit bacterial wilt disease in tobacco, tomato and arabidopsis. Plant Cell Physiol. 2012, 53, 1432–1444. [Google Scholar] [CrossRef]
- Zhao, L.; Li, S.R.; Zhang, H.Y.; Hao, T.Y.; Wu, D.; Wang, H. Preliminary study on enzymatic synthesis of sugar fatty acid esters and their antifungal activities. Food Ferment. Ind. 2016, 40, 32–36. [Google Scholar]
- Zhao, L.; Zhang, H.Y.; Hao, T.Y.; Li, S.R. In vitro antibacterial activities and mechanism of sugar fatty acid esters against five food-related bacteria. Food Chem. 2015, 187, 370–377. [Google Scholar] [CrossRef] [PubMed]
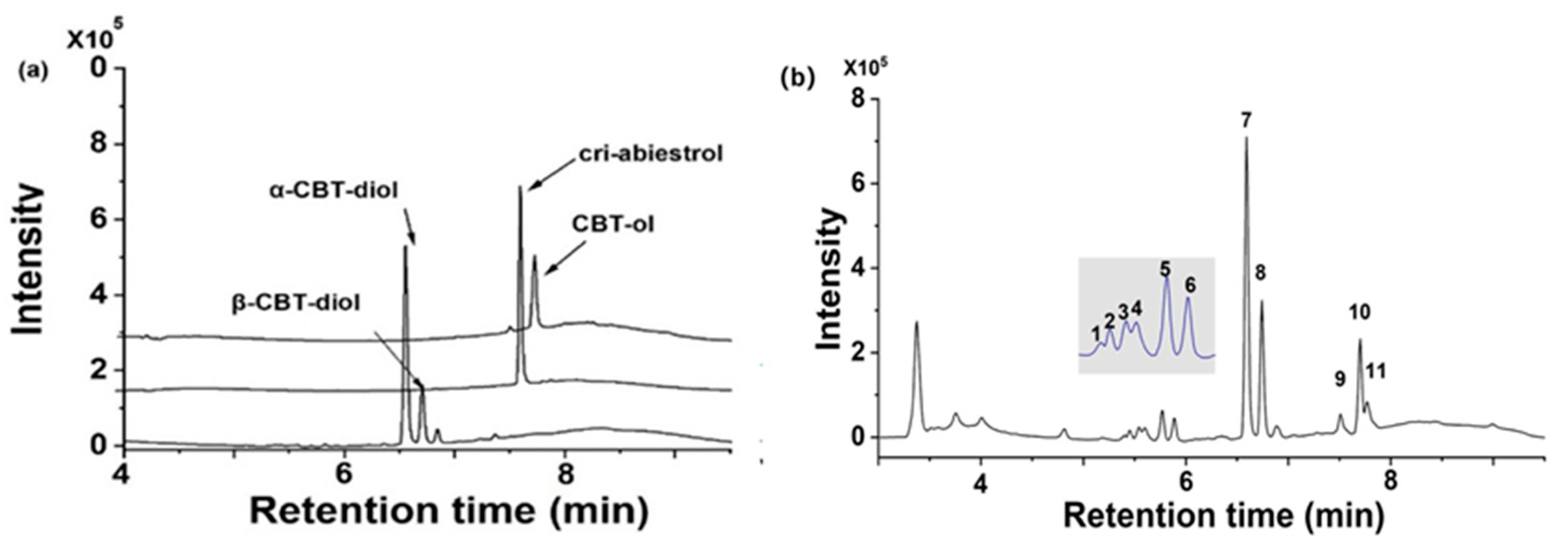
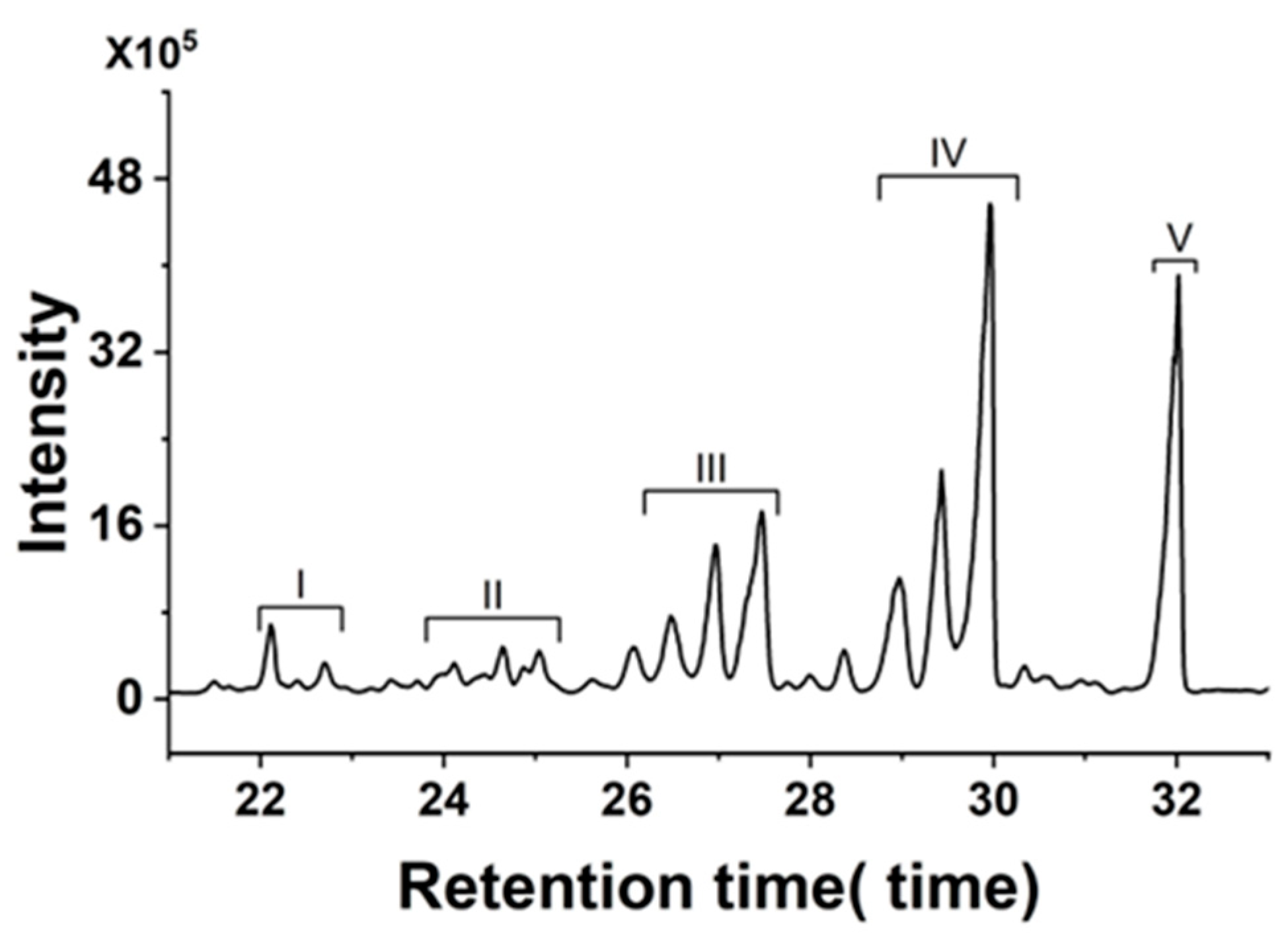



| Peak | Formula | Compound | tR (min) | Error (ppm) | Score | m/z |
|---|---|---|---|---|---|---|
| Peak1 | C20H34O4 | T1 | 4.3 min | −1.4 | 100 | 361.2341, 303.2304, 287.2356 |
| Peak2 | C20H34O4 | T2 | 4.4 min | −1.6 | 100 | 361.2340, 303.2310, 287.2366 |
| Peak3 | C20H34O4 | T3 | 4.5 min | 1.8 | 84.46 | 361.2396, 303.2420, 287.2361 |
| Peak4 | C22H40O5 | T4 | 4.7 min | −1.9 | 89.67 | 407.2768, 349.2734, 331.2226, 286.2242 |
| Peak5 | C20H34O3 | T5 | 4.9 min | −2.4 | 89.67 | 345.2492, 305.2476, 287.2361, 269.2351 |
| Peak6 | C20H32O3 | T6 | 5.1 min | 1.8 | 100 | 343.2238, 303.2311, 285.2209 |
| Peak7 | C20H34O2 | T7 | 6.6 min | 0.6 | 100 | 329.2453, 289.2523, 271.2417 |
| Peak8 | C20H34O2 | T8 | 6.9 min | −0.2 | 100 | 329.2451, 289.2522, 271.2417 |
| Peak9 | C20H32O | T9 | 8.4 min | 1.2 | 100 | 311.2345, 271.2414 |
| Peak10 | C20H34O | T10 | 8.6 min | 1.4 | 100 | 313.2446, 273.2521, 255.241 |
| Peak11 | C20H34O | T11 | 8.8 min | 1.3 | 100 | 313.2502, 273.2537 |
| Matrix | nLV | R2X | Q2 | RMSECV |
|---|---|---|---|---|
| 80 × 16 | 4 | 0.742 | 0.501 | 0.1892 |
| 60 × 16 | 4 | 0.762 | 0.593 | 0.1796 |
| 40 × 16 | 3 | 0.775 | 0.639 | 0.1877 |
| 30 × 16 | 2 | 0.861 | 0.747 | 0.1336 |
| 20 × 16 | 2 | 0.881 | 0.643 | 0.1511 |
| No. | Name | Type | Source | No. | Name | Type | Source |
|---|---|---|---|---|---|---|---|
| X1 | Havana-1 | cigar | introduced | X41 | Tangfeng | sun-cured | local |
| X2 | Havana 1 | cigar | introduced | X42 | Jiangyouyan | sun-cured | local |
| X3 | Beinhart 1000-1 | cigar | introduced | X43 | Wushanxiaolanyan | sun-cured | local |
| X4 | Criollo Salteno 11 | cigar | introduced | X44 | Tiebanqing | sun-cured | local |
| X5 | S-2 | cigar | introduced | X45 | Meitanshaiyan | sun-cured | local |
| X6 | Havana IIc | cigar | introduced | X46 | Hefengheiyan | sun-cured | local |
| X7 | Zrenjanin | cigar | introduced | X47 | Liufengmaobayan | sun-cured | local |
| X8 | Yinnixuejiabaopi | cigar | introduced | X48 | Jianpingpiaobayan | sun-cured | local |
| X9 | 112–117 | cigar | introduced | X49 | Dayeziqingyan | sun-cured | local |
| X10 | Bad Geudertheimer Landsorte | cigar | introduced | X50 | Lichuanmaoyan | sun-cured | local |
| X11 | Begej | cigar | introduced | X51 | Zhushandaliuzi | sun-cured | local |
| X12 | Connecticut Broad Leaf | cigar | introduced | X52 | Fengjiedamaoyan | sun-cured | local |
| X13 | Connecticut Shade | cigar | introduced | X53 | Shai9118 | sun-cured | breeding |
| X14 | E 18 | cigar | introduced | X54 | Shifangpipaliu | sun-cured | local |
| X15 | Geudetthelmex | cigar | introduced | X55 | Shiyan1 | sun-cured | local |
| X16 | Hanica | cigar | introduced | X56 | Kuiliu | sun-cured | local |
| X17 | Havana 211 | cigar | introduced | X57 | Bamaoliu | sun-cured | local |
| X18 | Havana 510 | cigar | introduced | X58 | Mianzhushaiyan | sun-cured | local |
| X19 | Manila | cigar | introduced | X59 | Bashan1 | sun-cured | local |
| X20 | Havana | cigar | introduced | X60 | Chongzhoushaiyan2 | sun-cured | local |
| X21 | Tuerqixueji | cigar | introduced | X61 | Quanyan | sun-cured | local |
| X22 | Conn Shade | cigar | introduced | X62 | Zhouyan | sun-cured | local |
| X23 | Dexue 1 | cigar | introduced | X63 | Xuejia5 | sun-cured | introduced |
| X24 | Dexue2 | cigar | introduced | X64 | New Havana IIc | cigar | introduced |
| X25 | Dexue 3 | cigar | introduced | X65 | Comstock Spanish | cigar | introduced |
| X26 | Habana92 | cigar | introduced | X66 | Mont Calme Brun | cigar | introduced |
| X27 | Cubra-Brazil | cigar | introduced | X67 | Trapesond 288 | cigar | introduced |
| X28 | Kangzhoukuoye | cigar | introduced | X68 | CA0705 | cigar | breeding |
| X29 | OLOR | cigar | introduced | X69 | CA0709 | cigar | breeding |
| X30 | Duominijiachangxin | cigar | introduced | X70 | Nilajiaguachangxin | cigar | introduced |
| X31 | Duominijiaduanxin | cigar | introduced | X71 | Besuki | cigar | introduced |
| X32 | MFPP | cigar | introduced | X72 | Ha20 | cigar | introduced |
| X33 | MFZS | cigar | introduced | X73 | Criollo | cigar | introduced |
| X34 | CP2011 | cigar | introduced | X74 | Ha12 | cigar | introduced |
| X35 | MSCA | cigar | introduced | X75 | Ha19 | cigar | introduced |
| X36 | CS0708 | cigar | breeding | X76 | Jilindabaihua | sun-cured | local |
| X37 | Shangzhiyiduohua | air-cured | local | X77 | Mulengdaqingjin | sun-cured | local |
| X38 | Shandongdaye | sun-cured | local | X78 | Juanyeshaiyan | aromatic | local |
| X39 | Xinbinxiaotuanye | sun-cured | local | X79 | Huangmaoyan | sun-cured | local |
| X40 | Liaoduoye | sun-cured | local | X80 | Baimaoyan | sun-cured | local |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Wang, J.; Du, Y.; Yan, N.; Han, X.; Zhang, J.; Dou, Y.; Liu, Y. Application and Evaluation of the Antifungal Activities of Glandular Trichome Secretions from Air/Sun-Cured Tobacco Germplasms against Botrytis cinerea. Plants 2024, 13, 1997. https://doi.org/10.3390/plants13141997
Liu J, Wang J, Du Y, Yan N, Han X, Zhang J, Dou Y, Liu Y. Application and Evaluation of the Antifungal Activities of Glandular Trichome Secretions from Air/Sun-Cured Tobacco Germplasms against Botrytis cinerea. Plants. 2024; 13(14):1997. https://doi.org/10.3390/plants13141997
Chicago/Turabian StyleLiu, Jing, Jiao Wang, Yongmei Du, Ning Yan, Xiao Han, Jianhui Zhang, Yuqing Dou, and Yanhua Liu. 2024. "Application and Evaluation of the Antifungal Activities of Glandular Trichome Secretions from Air/Sun-Cured Tobacco Germplasms against Botrytis cinerea" Plants 13, no. 14: 1997. https://doi.org/10.3390/plants13141997