Abstract
In the context of climate change, methods to improve the resistance of coniferous trees to biotic and abiotic stress are in great demand. The common plant response to exposure to vastly different stressors is the generation of reactive oxygen species (ROS) followed by activation of the defensive antioxidant system. We aimed to evaluate whether seed treatment with physical stressors can activate the activity of antioxidant enzymes and radical scavenging activity in young Picea abies (L.) H. Karst seedlings. For this, we applied seed treatment with cold plasma (CP) and electromagnetic field (EMF) and compared the response in ten different half-sib families of Norway spruce. The impact of the treatments with CP (1 min—CP1; 2 min—CP2) and EMF (2 min) on one-year-old and two-year-old P. abies seedlings was determined by the emergence rate, parameters of growth, and spectrophotometric assessment of antioxidant capacity (enzyme activity; DPPH and ABTS scavenging) in needles. The results indicated that the impact of seed treatment is strongly dependent on the genetic family. In the 577 half-sib family, the activity of antioxidant enzymes catalase (CAT), ascorbate peroxidase (APX), peroxidase (POX), and glutathione reductase (GR) increased after EMF-treatment in one-year-old seedlings, while similar effects in 477 half-sib family were induced by CP2 treatment. In two-year-old seedlings, CP1-treatment increased CAT, APX, POX, GR, SOD, DPPH, and ABTS activity in the 457 half-sib family. However, no significant impact of the treatment with CP1 was determined in one-year-old seedlings in this family. The application of novel technologies and the consideration of the combinatory impact of genetic and physical factors could have the potential to improve the accumulation of compounds that play an essential role in the defense mechanisms of P. abies. Nevertheless, for different resistance and responses to stressors of plants, their genetic properties play an essential role. A comprehensive analysis of interactions among the stress factors (CP and EMF), genetic properties, and changes induced in the antioxidant system can be of importance both for the practical application of seed treatment in forestry and for understanding fundamental adaptation mechanisms in conifers.
1. Introduction
Conifers are a highly resilient life form and one of the most successful organisms thanks to their evolutionary adaptations [1]. Even though, their long-life cycle and immobility increase their vulnerability to repeated abiotic and biotic stresses that affect conifer health [2,3,4]. Recent studies have examined the effects of biological control agents that can be used to control the expansion of conifer pests and pathogens or to increase stress resistance [5,6,7,8,9,10]. To increase the biologically active compounds in plant tissue, seed priming with physical stressors can be applied [11,12,13].
The application of low-temperature plasma (CP) and electromagnetic field (EMF) has a high potential in the forestry field, without a negative impact on the environment [14,15,16,17,18]. The advantage of plasma, in contrast to other stressors, is its capacity to generate active energy-containing species [13,19]. CP is comprised of charged particles, such as free electrons and ions, as well as neutral activated species, including gas molecules, free radicals, metastable particles, and produced UV photons [13,20]. EMF is a non-ionizing radiation, characterized by energy and momentum conveyed through alternating magnetic and electric fields [11]. It was recently demonstrated that seed priming with CP and EMF, besides positive effects on germination and seedling growth [13,21,22,23,24,25,26,27], also can enhance the synthesis of biologically active compounds and photosynthetic pigments in coniferous and deciduous tree species [13,18,21]. However, there is still a knowledge gap in the effects of seed treatments with CP and EMF on the antioxidant capacity in tissues of trees.
ROS are generated during metabolic processes serving essential signaling functions in living organisms [28,29,30,31]. In plants, the primary sources of ROS are the electron transport chains in chloroplasts and mitochondria, along with photorespiration in peroxisomes [31,32,33,34]. ROS impact on plants can vary depending on its concentration, the level of stress severity, the cellular energy status, and the antioxidant capacity [31,35].
The antioxidant capacity contributes to a plant’s survival when the stress levels surpass the natural defense mechanisms [36,37,38]. The plants’ ability to counteract oxidative stress relies on the effectiveness of their antioxidant system consisting of antioxidant enzymes and numerous molecular ROS scavengers like ascorbate, glutathione, etc. Superoxide dismutase (SOD; EC:1.15.1.1), catalase (CAT; EC:1.11.1.6), ascorbate peroxidase (APX; EC:1.11.1.11), peroxidase (POX; EC:1.11.1.5), and glutathione reductase (GR; EC 1.8.1.7) are the most abundant antioxidant enzymes in plants [37,39]. SOD is one of the major enzymes scavenging stress-generated free radicals, via converting superoxide •O2− to H2O2 and O2 [38]. CAT is engaged in the scavenging of H2O2 produced during photorespiration [38]. APX functions both as a scavenger of H2O2 and as a sensor for redox changes within plant cells [40]. POX operates in the extracellular space to eliminate H2O2 [38]. GR is nicotinamide adenine dinucleotide phosphate (NADPH) oxidoreductase, an essential component of the ascorbate-glutathione pathway catalyzing the reduction in oxidized glutathione (GSSG) to its reduced monomeric counterpart (GSH) [38]. The assessment of the antioxidant capacity in tissues provides important information for the evaluation of the physiological capacity of the tree defense system.
On the other side, genetic inter-species differences among plants can have a significant impact on response to different stressors and plant defense. Previous studies revealed that based on different half-sib families (genotypes), coniferous trees, similar to deciduous trees, can synthesize different amounts of secondary metabolites, sugars, photosynthetic pigments, and the capacity of antioxidant system [41,42,43,44]. In conifers, the relationship between these biologically active compounds and even slight changes in their synthesis can predetermine whether the tree is under stress [18]. Therefore, the selection of P. abies genotypes based on the metabolic pathways in needles by comparing changes induced by seed priming with CP and EMF in the content of biologically active compounds and antioxidants in seedlings might have a potential for producing forest stands with the enhanced resistance both to biotic and abiotic stress. However, large variation between the genotypes after such stressors makes it complicated to apply procedures, and more studies are needed.
The main purpose of this study was to assess the impact of pre-sowing seed treatment with CP and EMF on the antioxidant capacity in the needles of different P. abies half-sib families, aiming to estimate how the antioxidant defense mechanisms are affected. We hypothesized that: (1) seed treatment with physical stressors has an impact on antioxidant capacity in one-year-old and two-year-old P. abies seedlings; (2) induced changes in antioxidant capacity depend on the treatment method and duration; (3) the response to the treatment is different among the studied half-sib families.
2. Results
2.1. Emergence of P. abies Seedlings
The emergence percentage of control P. abies seedlings varied substantially (from 36% to 88%) among half-sib families. The emergence percentage in the control and CP-treated groups was the lowest in the 599 half-sib family and the highest in the 417 half-sib family (Table 1). Large variation was also observed in CP1-, CP2- and EMF2-treated groups (from 36% to 79%, from 39% to 85%, and from 31% to 81%, respectively).

Table 1.
Picea abies emergence percentage (%) in different half-sib families. C—control; CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks indicate the statistical significance of the difference (Student’s test) between the treated group (CP1, CP2, and EMF2) and the control (C) in each half-sib family (* p < 0.05; ** p < 0.01; *** p < 0.001).
Seed treatment with CP1 did not affect seedling emergence in eight of the tested half-sib families; however, in 417 and 454 half-sib families negative effect was observed (reduced emergence by 9% and 30%, respectively). The CP2 treatment significantly increased emergence in 477 half-sib families (by 35%). However, longer treatment with CP has a negative impact on seedling emergence in 454, 541, and 577 half-sib families (reduced it by 24%, 17%, and 17%, respectively). Seed treatment with EMF resulted in a higher emergence percentage of seedlings in one family (463) compared to the control (by 21%); meanwhile, the EMF reduced emergence in the 417 and 454 half-sib families by 18% and 17%, respectively.
2.2. Height of P. abies Seedlings
The height of the above-ground part of the seedlings was measured to assess the growth of seedlings during both the first and second vegetation seasons. The highest height among the control one-year-old seedlings was observed in the 454 and 541 half-sib families (Table 2). Meanwhile, the highest height in the control two-year-old spruce seedlings was in the 463 half-sib family (Table 2).
Table 2.
Height of one-year-old and two-year-old seedlings in different Picea abies half-sib families. C—control; CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks indicate the statistical significance of the difference (Student’s test) between the treated group (CP1, CP2, and EMF2) and the control (C) in each half-sib family (* p < 0.05; ** p < 0.01; *** p < 0.001).
CP1 effect on height of one-year-old seedlings in P. abies half-sib families was diverse. Four half-sib families (417, 457, 477, and 548) exhibited an increase in height by 16-19%, respectively, while seedlings from 463 families were 57% higher compared to the control (Table 3). However, two families (454 and 577) had reduced height by more than 10% due to seed treatment with the CP1. Meanwhile, a longer seed treatment (CP2) resulted in increased height in four half-sib families (457, 463, 548, and 599) by 18%, 19%, 30%, and 10%, respectively. A negative CP2 effect on height was observed in the same half-sib families as for CP1 with a reduction of 16% and 38%, respectively. EMF-treatment increased height by 13% in one half-sib family only (457) but reduced height in 417, 454, 541, and 577 half-sib families by 14%, 11%, 10%, and 33%, respectively.
Three half-sib families (417, 457, and 541) from CP1 treated groups exhibited an increase in two-years-old seedling height by 25%, 13%, and 11%, respectively. However, a longer seed treatment with CP and EMF did not have a significant positive impact on height. On the contrary, a negative effect of CP2 and EMF2 on height was observed in three half-sib families (CP2 reduced growth of 463 and 577 half-sib families; EMF2—541 half-sib family seedlings) with the height lower by more than 10%.
2.3. Changes of Activity of Antioxidant Enzymes in Needles of P. abies
The results of the measurement of antioxidant enzyme (CAT, APX, POX, GR, and SOD) activities in P. abies needles are presented in Figure 1, Figure 2, Figure 3, Figure 4 and Figure 5.
2.3.1. Catalase (CAT)
The obtained results revealed that CAT activity was three times higher in two-year-old seedlings compared to younger ones, regardless of the half-sib family and the treatment. The highest CAT activity in the control samples in one-year-old seedlings was in the 124, 463, and 599 half-sib families (Figure 1a). Meanwhile, the highest CAT activity in the control samples in two-year-old seedlings was determined in the 124, 454, and 541 half-sib families (Figure 1b).
In one-year-old seedlings, CP1-treatment increased CAT activity in one half-sib family (417) by 27% (Figure 1a). However, the CAT activity in the 124 half-sib family was reduced by 7%. Meanwhile, a longer seed treatment (CP2) resulted in increased CAT activity in three half-sib families (417, 477, and 577) by 27%, 8%, and 13%, respectively, compared to control. However, in the 457 half-sib family CAT activity was reduced by 9% after CP2-treatment. Seed treatment with EMF resulted in enhanced CAT activity in four half-sib families (417, 457, 463, and 577) by 27%, 5%, 8%, and 13%, respectively. The negative effect on CAT activity after seed treatment with EMF was determined in the 548 and 599 half-sib families (the activity was lower by 8% and 12%, respectively). In one family (417), CAT activity was higher after seed treatment with all physical stressors.
In two-year-old seedlings, seed treatment with CP1 increased CAT activity in three half-sib families (457, 463, and 548) by 22%, 14%, and 16%, respectively but reduced by 6% in the 477 half-sib family (Figure 1b). CP2 treatment increased enzyme activity in four half-sib families (124, 417, 457, and 548) by 12%, 14%, 12%, and 16%, respectively. However, in the 454 half-sib family CAT activity was reduced by 15% after CP2-treatment. Meanwhile, EMF-treatment increased CAT activity in six half-sib families –124, 457, 477, 548, and 599 by 8–16%, while in 463 families—by 35%. The negative effect on CAT activity after seed treatment with EMF was observed in the 454 and 541 half-sib families (the activity was lower by 20%). In two half-sib families (457 and 548), CAT activity was higher after seed treatment with all physical stressors, compared to control.
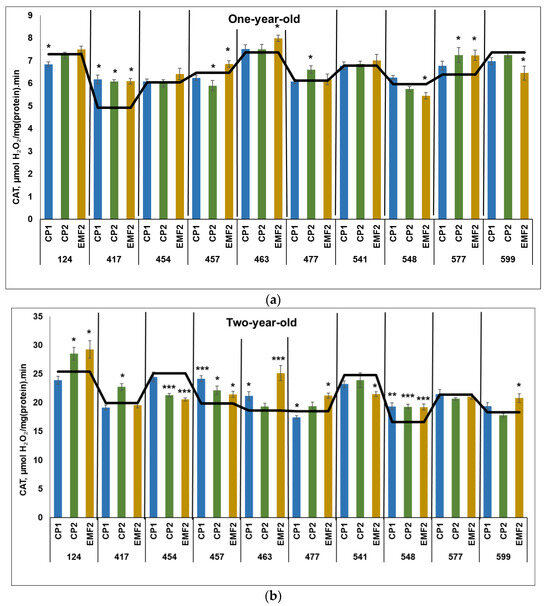
Figure 1.
CAT activity in needles of (a) one-year-old and (b) two-year-old seedlings of different half-sib families of Picea abies (N = 360; 3 biological replicates × 3 technical replicates for each sample). The black line in the graphs shows the mean values of control; CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks indicate the statistical significance of the difference (Student’s test) between the treated group (CP1, CP2, and EMF2) and the control (C) for each half-sib family (* p < 0.05; ** p < 0.01; *** p < 0.001).
Figure 1.
CAT activity in needles of (a) one-year-old and (b) two-year-old seedlings of different half-sib families of Picea abies (N = 360; 3 biological replicates × 3 technical replicates for each sample). The black line in the graphs shows the mean values of control; CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks indicate the statistical significance of the difference (Student’s test) between the treated group (CP1, CP2, and EMF2) and the control (C) for each half-sib family (* p < 0.05; ** p < 0.01; *** p < 0.001).
2.3.2. Ascorbate Peroxidase (APX)
The results showed that APX activity was higher in two-year-old seedlings, compared to one-year-old ones, regardless of the half-sib family and treatment. The highest APX activity in the control samples in one-year-old seedlings was determined in the 124, 463, and 541 half-sib families (Figure 2a). Meanwhile, the highest APX activity in the control samples in two-year-old seedlings was observed in the 454 half-sib family (Figure 2b).
In one-year-old seedlings, CP1-treatment increased APX activity in three half-sib families (417, 454, 577) by 22%, 9%, and 9%, respectively (Figure 2a). CP2 treatment resulted in increased APX activity also in three half-sib families (417, 477, and 577) by 19%, 16%, and 13%. However, the APX activity in the 457 half-sib family was reduced by 10%. Meanwhile, EMF-treatment increased APX activity in two half-sib families (417 and 577) by 19% and 10%, respectively. In two half-sib families (417 and 577), APX activity was higher after seed treatment with all physical stressors, compared to control.
Two-years-old seedlings from 457 and 548 half-sib families showed increased APX activity by 19% and 9%, respectively, due to seed treatment with CP1 (Figure 2b). However, the APX activity in the 124 and 477 half-sib families was reduced (less than 10%). Meanwhile, CP2 treatment increased APX activity in three half-sib families (124, 457, and 548) by 12%, 10%, and 9%, respectively. However, in the 454 and 599 half-sib families APX activity was reduced by 20% and 5%, respectively. Seed treatment with EMF increased APX activity in five half-sib families (124, 463, 477, 548, and 599) by 12%, 24%, 9%, 9%, and 8%, respectively. On the contrary, a negative effect of EMF treatment on APX activity was observed in 454 and 541 half-sib families (the APX activity was reduced by 21% and 12%, respectively). In 548 half-sib families only APX activity was higher after seed treatment with all physical stressors, compared to control. Both CP1 and CP2 treatments increased APX activity in the 457 half-sib family.
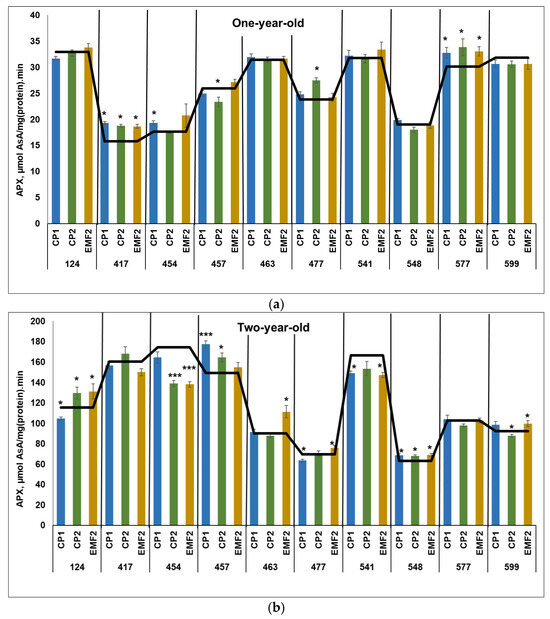
Figure 2.
APX activity in needles of (a) one-year-old and (b) two-year-old seedlings of different half-sib families of Picea abies (N = 360; 3 biological replicates × 3 technical replicates for each sample). The black line in the graphs shows the mean values of control; CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks mean the statistical significance of the difference (Student’s test) between the treated group (CP1, CP2, and EMF2) and the control (C) in each half-sib family (* p < 0.05; *** p < 0.001).
Figure 2.
APX activity in needles of (a) one-year-old and (b) two-year-old seedlings of different half-sib families of Picea abies (N = 360; 3 biological replicates × 3 technical replicates for each sample). The black line in the graphs shows the mean values of control; CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks mean the statistical significance of the difference (Student’s test) between the treated group (CP1, CP2, and EMF2) and the control (C) in each half-sib family (* p < 0.05; *** p < 0.001).
2.3.3. Peroxidase (POX)
The results indicated that POX activity was higher in two-year-old seedlings compared to younger ones, regardless of the half-sib family and treatment. The highest POX activity in the control samples in one-year-old seedlings was determined in the 457 and 477 half-sib families (Figure 3a). Meanwhile, the highest POX activity in the control samples in two-year-old seedlings was ascertained in the 577 half-sib family (Figure 3b).
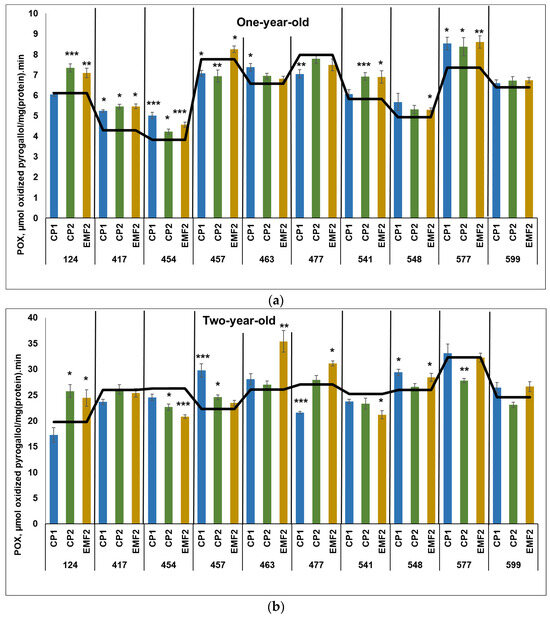
Figure 3.
POX activity in needles of (a) one-year-old and (b) two-year-old seedlings of different half-sib families of Picea abies (N = 360; 3 biological replicates × 3 technical replicates for each sample). The black line in the graphs shows the mean values of control. CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks indicate the statistical significance of the difference between (Student’s test) the treated group (CP1, CP2, and EMF2) and the control (C) in each half-sib family (* p < 0.05; ** p < 0.01; *** p < 0.001).
Figure 3.
POX activity in needles of (a) one-year-old and (b) two-year-old seedlings of different half-sib families of Picea abies (N = 360; 3 biological replicates × 3 technical replicates for each sample). The black line in the graphs shows the mean values of control. CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks indicate the statistical significance of the difference between (Student’s test) the treated group (CP1, CP2, and EMF2) and the control (C) in each half-sib family (* p < 0.05; ** p < 0.01; *** p < 0.001).
In one-year-old seedlings, CP1 treatment increased POX activity in four half-sib families (417, 454, 463, and 577) by 21, 32, 12%, and 15%, respectively (Figure 3a). However, in two families (457 and 477) CP1 reduced POX activity (by 9% and 12%, respectively). Meanwhile, CP2 increased POX activity in five half-sib families (124, 417, 454, 541, and 577) by 20%, 28%, 11%, 19% and 14%, respectively. On the contrary, seed treatment with CP2 reduced POX activity in the 457 half-sib family only (by 12%). Seed treatment with EMF increased POX activity in seven half-sib families (124, 417, 454, 457, 541, 548, and 577) by 16%, 28%, 21%, 5%, 19%, 8%, and 16%, respectively. In three half-sib families (417, 454, and 577), POX activity was higher after seed treatment with all physical stressors, compared to the control.
CP1 treatment increased POX activity in two half-sib families (457 and 548) by 33% and 14%, respectively, in two-year-old seedlings but reduced it by 21% in the 477 half-sib family (Figure 3b). Seed treatment with CP2 led to increased POX activity in the seedling needles from the two half-sib families (124 and 457) by 30% and 10%, respectively. In the two half-sib families (454 and 577) POX activity was reduced by 14% after CP2-treatment. Meanwhile, EMF-treatment increased POX activity in four half-sib families (124, 463, 477, and 548) by 23%, 36%, 15%, and 7%, respectively. However, a negative effect of EMF on POX activity was observed in 454 and 541 half-sib families (POX activity reduced by 21% and 16%, respectively).
2.3.4. Glutathione Reductase (GR)
The results demonstrated that the activity of GR was higher more than 2.5 times in two-year-old seedlings compared to younger ones, regardless of the half-sib family and the treatment. The highest GR activity in the control samples in one-year-old seedlings was determined in the 548 half-sib family (Figure 4a). Meanwhile, the highest GR activity in the control samples in two-year-old seedlings was observed in the 454 half-sib family (Figure 4b).
One-year-old seedlings from 454, 463, and 577 half-sib families showed increased GR activity (by 7%, 8%, and 11%, respectively) due to seed treatment with CP1 (Figure 4a). However, CP1 treatment was negatively affected by 18% GR activity in the 548 half-sib family. CP2 treatment resulted in increased GR activity in two half-sib families (124 and 477) by 8% and 5%, respectively. However, GR activity in 548 half-sib families was reduced by 18%. Meanwhile, seed treatment with EMF increased GR activity in three half-sib families 124, 457, and 577 by 13%, 8%, and 8%, respectively. On the contrary, EMF reduced GR activity in the 548 half-sib family by 18%.
In two-year-old seedlings, CP1-treatment increased GR activity in half-sib families of 457, 548, and 599 by 25%, 9%, and 8%, respectively, but slightly reduced (by less than 10%) GR activity in the 477 and 541 half-sib families (Figure 4b). CP2 increased GR activity in three half-sib families (124, 457, and 548) by 15%, 11%, and 7%, respectively, and reduced it in the 454 half-sib family GR activity by 21%. Meanwhile, EMF-treatment increased GR activity in five half-sib families (124, 463, 477, 548, and 599) by 17%, 28%, 1%, 8%, and 12%, respectively. The negative effect EMF-treatment on GR activity was determined in the 454 and 541 half-sib families (by 21% and 13%, respectively). All physical stressors used for seed treatments induced an increase in GR activity compared to control only in the needles of the 548 half-sib family. Seed treatment with both CP1 and CP2 caused higher GR activity in the 457 half-sib family.
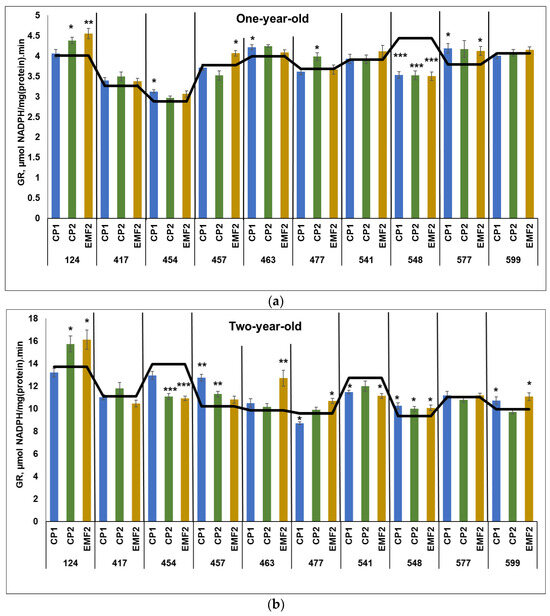
Figure 4.
GR activity in needles of (a) one-year-old and (b) two-year-old seedlings of different half-sib families of Picea abies (N = 360; 3 biological replicates × 3 technical replicates for each sample). The black line in the graphs shows the mean values of control; CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks indicate the statistical significance of the difference (Student’s test) between the treated group (CP1, CP2, and EMF2) and the control (C) in each half-sib family (* p < 0.05; ** p < 0.01; *** p < 0.001).
Figure 4.
GR activity in needles of (a) one-year-old and (b) two-year-old seedlings of different half-sib families of Picea abies (N = 360; 3 biological replicates × 3 technical replicates for each sample). The black line in the graphs shows the mean values of control; CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks indicate the statistical significance of the difference (Student’s test) between the treated group (CP1, CP2, and EMF2) and the control (C) in each half-sib family (* p < 0.05; ** p < 0.01; *** p < 0.001).
2.3.5. Superoxide Dismutase (SOD)
SOD activity was higher in two-year-old seedlings compared to one-year-old ones, regardless of the half-sib family and the treatment. The highest SOD activity in the control groups of one-year-old seedlings was detected in the 124 half-sib family (Figure 5a), and in the control samples in two-year-old seedlings—in the 477 half-sib family (Figure 5b).
CP-treatment increased SOD activity in one half-sib family (463 after CP1-treatment and 417 after CP2-treatment) by 22% and 28%, respectively in one-year-old seedlings (Figure 5a). However, the SOD activity in the 599 half-sib family was reduced by 30%. In addition, CP2 treatment caused a negative effect in most families—compared to the control, SOD activity was reduced by 13%, 21%, 24%, 28%, and 21% in 124, 454, 548, 577, and 599 half-sib families, respectively. Moreover, EMF-treatment reduced SOD activity in seven half-sib families (417, 454, 457, 477, 548, 577, and 599) by 36%, 29%, 17%, 16%, 32%, 38%, and 21%, respectively.
When SOD activity was assessed in two-year-old seedlings, a more significant impact of seed treatments was found as compared with one-year-old seedlings (Figure 5b). Seed treatment with CP1 enhanced SOD activity in five half-sib families (124, 457, 541, 548, and 599) by 50%, 10%, 18, 13%, and 83%, respectively, although SOD activity in the 417, 454, and 477 half-sib families was reduced by 10%. CP2 increased SOD activity in four half-sib families (124, 417, 541, and 577) by 37%, 33%, 18%, and 13%, respectively, but reduced it in 454, 457, and 477 half-sib families (the activity reduced by 30%, 9%, and 24%, respectively). Meanwhile, EMF-treatment increased SOD activity in the 417 half-sib families by 20%. The negative effect of EMF on SOD activity was observed in the 454, 457, 477, 541, and 548 half-sib families (the activity was lower by 27%, 31%, 39%, 22%, and 24%, respectively). In two half-sib families (124 and 541) SOD activity was increased by both CP1 and CP2 treatments.
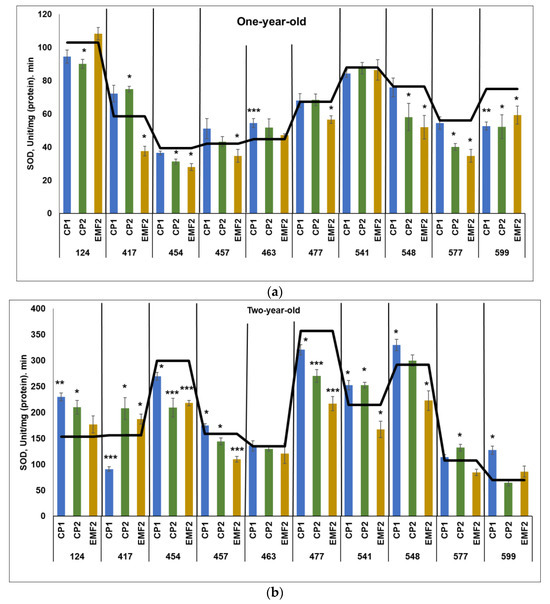
Figure 5.
SOD activity in needles of (a) one-year-old and (b) two-year-old seedlings of different half-sib families of Picea abies (N = 360; 3 biological replicates × 3 technical replicates for each sample). The black line in the graphs shows the mean values of control; CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks mean the statistical significance of the difference (Student’s test) between the treated group (CP1, CP2, and EMF2) and the control (C) in each half-sib family (* p < 0.05; ** p < 0.01; *** p < 0.001).
Figure 5.
SOD activity in needles of (a) one-year-old and (b) two-year-old seedlings of different half-sib families of Picea abies (N = 360; 3 biological replicates × 3 technical replicates for each sample). The black line in the graphs shows the mean values of control; CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks mean the statistical significance of the difference (Student’s test) between the treated group (CP1, CP2, and EMF2) and the control (C) in each half-sib family (* p < 0.05; ** p < 0.01; *** p < 0.001).
2.4. Changes of Antioxidant Activity in P. abies Needles
The results of the conducted measurements of antioxidant activity (DPPH and ABTS) in P. abies needles are presented in Table 3.
Table 3.
Radical scavenging activity in needles of (a) one-year-old and (b) two-year-old seedlings of different half-sib families of Picea abies. C—control; CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks indicate the statistical significance of the difference (Student’s test) between the treated group (CP1, CP2, and EMF2) and the control (C) in each half-sib family (* p < 0.05; ** p < 0.01; *** p < 0.001).
Table 3.
Radical scavenging activity in needles of (a) one-year-old and (b) two-year-old seedlings of different half-sib families of Picea abies. C—control; CP1—seed treatment with cold plasma 1 min; CP2—seed treatment with cold plasma 2 min; EMF2—seed treatment with electromagnetic field 2 min. The asterisks indicate the statistical significance of the difference (Student’s test) between the treated group (CP1, CP2, and EMF2) and the control (C) in each half-sib family (* p < 0.05; ** p < 0.01; *** p < 0.001).
| (a) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Half-Sib Family | Radical Scavenging Activity (µmol/g) ± SE One-Year-Old | |||||||
| DPPH Assay | ABTS Assay | |||||||
| C | CP1 | CP2 | EMF2 | C | CP1 | CP2 | EMF2 | |
| 124 | 78.0 ± 5.7 | 81.3 ± 8.5 | 75.8 ± 10.6 | 76.3 ± 6.6 | 591.4 ± 24.3 | 574.6 ± 22.6 | 629.7 ± 18.0 | 606.8 ± 14.8 |
| 417 | 86.9 ± 9.2 | 72.8 ± 11.7 | 51.3 ± 7.1 * | 90.5 ± 8.8 | 790.9 ± 21.9 | 734.5 ± 14.6 * | 642.9 ± 20.6 *** | 805.6 ± 33.8 |
| 454 | 78.0 ± 7.6 | 63.3 ± 6.8 | 61.9 ± 13.6 | 69.7 ± 7.7 | 471.7 ± 12.2 | 517.4 ± 18.6 * | 586.2 ± 30.4 ** | 639.0 ± 22.4 *** |
| 457 | 72.8 ± 6.3 | 88.4 ± 8.2 | 85.7 ± 6.3 | 80.4 ± 7.3 | 610.9 ± 26.9 | 641.0 ± 6.1 | 616.2 ± 17.9 | 549.4 ± 21.4 |
| 463 | 72.6 ± 6.6 | 75.8 ± 12.2 | 93.0 ± 7.6 * | 92.3 ± 6.0 * | 539.0 ± 10.6 | 635.6 ± 11.2 *** | 622.0 ± 13.3 *** | 592.4 ± 10.0 * |
| 477 | 71.1 ± 10.7 | 71.5 ± 6.1 | 65.2 ± 8.9 | 70.8 ± 8.2 | 475.6 ± 20.1 | 462.2 ± 18.2 | 534.4 ± 33.5 | 579.8 ± 10.7 *** |
| 541 | 68.2 ± 8.8 | 48.7 ± 6.7 | 68.8 ± 5.8 | 65.0 ± 6.4 | 712.8 ± 81.9 | 573.9 ± 17.6 | 695.6 ± 12.2 | 613.3 ± 12.7 |
| 548 | 83.9 ± 4.7 | 64.7 ± 6.4 * | 75.1 ± 5.3 | 78.5 ± 6.0 | 559.9 ± 12.9 | 549.5 ± 22.7 | 616.5 ± 14.2 * | 628.2 ± 19.3 * |
| 577 | 80.8 ± 7.3 | 104.2 ± 6.6 * | 95.0 ± 7.2 | 89.0 ± 9.6 | 614.7 ± 17.2 | 576.3 ± 16.0 | 600.0 ± 12.1 | 596.0 ± 20.5 |
| 599 | 73.4 ± 12.0 | 81.0 ± 10.7 | 89.5 ± 6.1 | 83.2 ± 7.6 | 733.1 ± 26.1 | 803.8 ± 30.0 | 718.1 ± 18.6 | 662.3 ± 22.8 * |
| (b) | ||||||||
| Half-Sib Family | Radical Scavenging Activity (µmol/g) ± SE Two-Year-Old | |||||||
| DPPH Assay | ABTS Assay | |||||||
| C | CP1 | CP2 | EMF2 | C | CP1 | CP2 | EMF2 | |
| 124 | 587.8 ± 13.3 | 633.9 ± 9.7 * | 560.6 ± 10.6 | 601.7 ± 20.8 | 1418.3 ± 24.8 | 1397.7 ± 29.1 | 1206.2 ± 27.6 *** | 1243.1 ± 25.0 *** |
| 417 | 650.6 ± 10.7 | 599.9 ± 14.9 * | 614.6 ± 9.8 * | 681.4 ± 14.8 | 1402.0 ± 20.9 | 1322.1 ± 24.0 * | 1324.0 ± 17.1 * | 1468.6 ± 31.4 |
| 454 | 563.4 ± 19.5 | 579.3 ± 11.6 | 559.6 ± 9.0 | 542.8 ± 14.2 | 1117.1 ± 14.3 | 1199.0 ± 35.6 * | 1099.5 ± 17.6 | 1094.6 ± 23.4 |
| 457 | 559.2 ± 16.7 | 616.4 ± 18.1 * | 712.2 ± 19.1 *** | 660.5 ± 10.8 *** | 1283.1 ± 22.6 | 1364.1 ± 28.4 * | 1510.3 ± 40.5 *** | 1375.8 ± 22.9 * |
| 463 | 584.6 ± 13.4 | 569.8 ± 15.7 | 613.1 ± 16.3 | 566.9 ± 16.6 | 1053.4 ± 17.2 | 1122.9 ± 21.4 * | 1230.4 ± 32.7 *** | 996.8 ± 20.9 * |
| 477 | 617.6 ± 12.2 | 668.0 ± 20.1 * | 594.7 ± 12.2 | 774.0 ± 43.6 ** | 1339.7 ± 23.2 | 1366.1 ± 27.5 | 1169.3 ± 36.3 | 1557.1 ± 83.8 |
| 541 | 581.9 ± 13.4 | 605.1 ± 14.8 | 537.9 ± 10.0 * | 559.7 ± 10.4 | 1159.1 ± 24.5 | 1140.1 ± 19.1 | 1203.1 ± 26.6 | 1375.6 ± 43.3 *** |
| 548 | 586.7 ± 15.1 | 537.5 ± 14.4 * | 546.2 ± 12.8 * | 573.7 ± 20.3 | 1125.3 ± 42.5 | 1097.4 ± 27.2 | 1056.2 ± 39.0 | 1079.3 ± 37.4 |
| 577 | 557.7 ± 13.3 | 551.2 ± 12.6 | 626.1 ± 20.8 * | 469.4 ± 13.2 *** | 1058.4 ± 13.5 | 1035.4 ± 26.0 | 1064.5 ± 28.6 | 796.1 ± 17.1 *** |
| 599 | 662.0 ± 16.1 | 607.5 ± 17.0 * | 678.3 ± 11.7 | 660.3 ± 17.9 | 1409.9 ± 35.7 | 1266.1 ± 33.1 ** | 1308.5 ± 22.9 * | 1367.4 ± 19.7 |
Radical scavenging activity in the different half-sib families of P. abies needles was determined using two tests: DPPH (2,2-diphenyl-1-picryl-hydrazylhydrate) and ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) assays. The difference in antioxidant activity in needles between one-year-old and two-year-old seedlings in the same experimental group was statistically significant differences (p < 0.05) in all half-sib families. The antioxidant activity was higher eight times (DPPH) and two times (ABTS) in two-year-old seedlings compared to one-year-old seedlings (Table 3). In the control samples, the highest antioxidant activity in one-year-old seedlings (86.93 µmol/g (DPPH) and 790.86 µmol/g (ABTS), respectively) was observed in the 417 half-sib family, while the other two families exhibited the highest values in two-year-old seedlings 599 (661.96 µmol/g (DPPH) and 124 (1418.29 µmol/g (ABTS) half-sib families (Table 3). The antioxidant activity in groups of seedlings growing from treated seeds was also dependent on the half-sib family, and different families exhibited diverse responses to the used physical stressors and treatment duration.
Seed treatment with CP1 increased antioxidant activity (DPPH assay) only in the 577 half-sib family (activity was higher by 29%) but reduced the antioxidant activity in the 548 half-sib family by 23% in one-year-old seedlings (Table 3a). However, CP2 treatment increased activity (by 28%) only in the 463 half-sib family, and a similar trend was observed after seed treatment with EMF. Compared to DPPH, different changes were revealed using ABTS assay. In one-year-old seedlings, seed treatment with CP1 increased antioxidant activity in two half-sib families (454 and 463 by less than 20%) and reduced it in 417 half-sib families. Meanwhile, CP2-treatment increased antioxidant activity in three half-sib families 454, 463, and 548 by 24%, 15%, and 10%, respectively. The activity was reduced in 417 half-sib families, as in the case with CP1 treatment. EMF-treatment increased antioxidant activity in the 454, 463, 477 and 548 half-sib families (by 35%, 10%, 22%, and 12%, respectively), but reduced it by less than 10% in the 599 half-sib family. In two families (454 and 463) antioxidant activity (using the ABTS method) was higher after seed treatment with all protocols.
The differences in ABTS activity in one half-sib family (541) increased by 36% in the CP1-treated group, compared to the control (Table 3a). In addition, the differences in ABTS activity in the 599 half-sib family were reduced by 35% in the CP1-treated group, compared to the control. The differences in ABTS activity in the 457 half-sib family increased by 35% in the CP2-treated group, compared to the control. Meanwhile, the differences in ABTS activity in three half-sib families (124, 454, and 477) reduced by 48%, 49%, and 63%, respectively in the CP2-treated group, compared to the control. On the other hand, two half-sib families (457 and 541) increased in differences of ABTS activity by 40% and 62%, respectively in the EMF-treated group, compared to the control. However, the differences in ABTS activity in three half-sib families (124, 454, and 577) were reduced by 35%, 66%, and 39%, respectively compared to the control groups.
Seed treatment with CP1 resulted in increased antioxidant activity in two-year-old seedlings (using DPPH assay) in three half-sib families (124, 457, and 477) by 8%, 10%, and 8%, respectively (Table 3b). However, seed treatment with CP1 reduced antioxidant activity in 417, 548, and 599 half-sib families by 8%. Meanwhile, CP2-treatment increased antioxidant activity in two half-sib families (457 and 577 by 27% and 12%, respectively), but reduced antioxidant activity by less than 10% in three half-sib families (417, 541, and 548). EMF-treatment resulted in higher antioxidant activity in two half-sib families (457 and 477 by 18% and 25%, respectively) and reduced antioxidant activity in the 577 half-sib family by 19%. In one family (457) antioxidant activity (using the DPPH method) was higher after seed treatment with all physical stressors.
It was observed using ABTS assay, that antioxidant activity was higher (by less than 10%) in the 454, 457, and 463 half-sib families (Table 3b). However, CP1 treatment reduced antioxidant activity in 417 and 599 half-sib families (by less than 10%). Seed treatment with CP2 caused an increase in antioxidant activity in the 457 and 463 half-sib families by 17%. However, in the 124, 417, and 599 half-sib families activity was reduced (by less than 15%). EMF-treatment increased antioxidant activity in 457 and 541 half-sib families by 7% and 17%, respectively, and reduced it in three half-sib families (124, 463, and 577 by 12%, 5%, and 25%, respectively).
2.5. Heatmap, Correlation, and Principal Component Analysis (PCA) between Antioxidant Enzymes and Antioxidant Activity
MANOVA analysis showed that the age of seedlings, different half-sib families, and the treatment had a significant influence on P. abies antioxidant system (enzymes, DPPH, and ABTS) with the largest impact by an age of seedlings (Wilks lambda—0.02, p < 0.001).
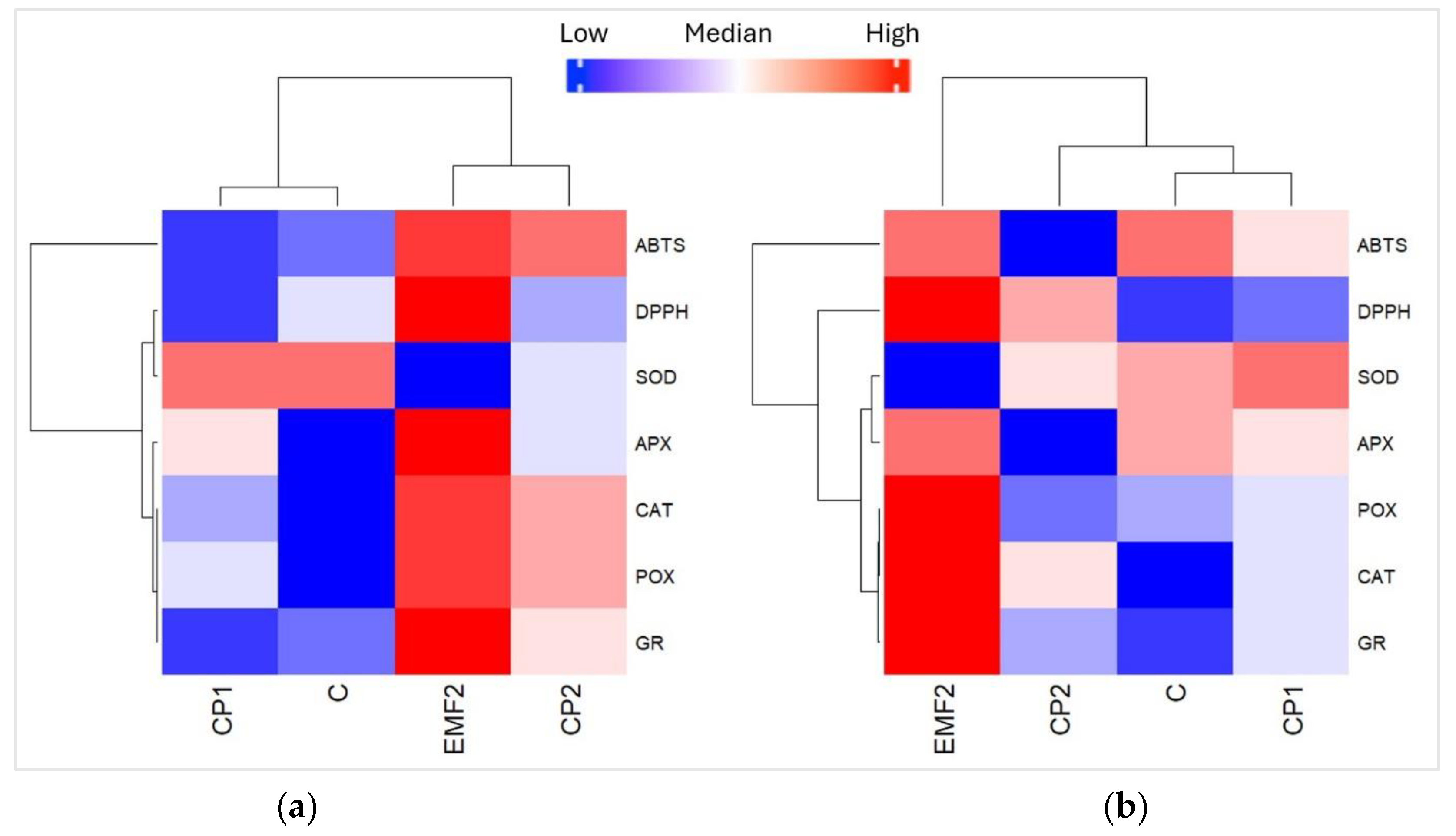
Heatmaps display the variation in the content of enzymes (CAT, APX, POX, GR, and SOD) and antioxidant activity (ABTS and DPPH) in the needles of one-year-old (Figure 6a) and two-year-old (Figure 6b) seedlings in groups growing from the seeds treated with physical stressors (CP1, CP2, and EMF). Blue colors indicate that the value of the analyzed variable was below the mean level, while red colors indicate values higher than the mean. For one-year-old seedlings, results obtained for seedlings growing from untreated seeds (C) and those treated with CP1, CP2, and EMF were clustered into two groups: the first cluster contained CP1 and C samples, while the second cluster contained EMF and CP2 (Figure 7a). The analysis shows that POX activity was significantly higher (p < 0.05) in the needles from one-year-old seedlings growing from seeds treated with EMF, compared to the control group. In contrast, EMF decreased the activity of SOD. Analyzing two-year-old seedlings, the results obtained from C, CP1, CP2, and EMF2 groups were clustered into three groups, where the first cluster contained C and CP1, the second—included CP2, and the third—included EMF (Figure 6b). The EMF-induced changes in the activity of POX and SOD followed the same trend as in one-year-old seedlings. In addition, EMF increased the activity of CAT in the needles of the two-year-old seedlings.
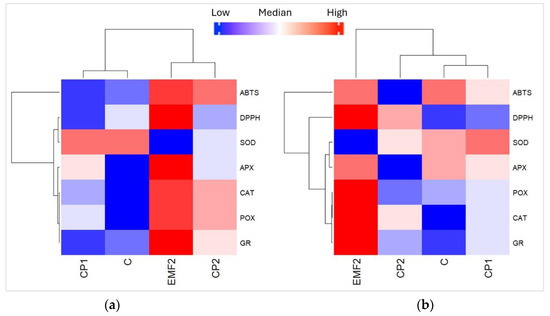
Figure 6.
Heatmap for different enzymes and antioxidant activity of needles from (a) one-year-old and (b) two-year-old Picea abies seedlings growing from the seeds treated by CP1, CP2, and EMF2. Control (C) represents needles from seedlings growing from non-treated seeds.
Figure 6.
Heatmap for different enzymes and antioxidant activity of needles from (a) one-year-old and (b) two-year-old Picea abies seedlings growing from the seeds treated by CP1, CP2, and EMF2. Control (C) represents needles from seedlings growing from non-treated seeds.
The correlation analysis revealed a strong positive dependence between activities of GR, POX, APX, and CAT antioxidant enzymes, especially between GR—CAT (r = 0.72) and GR—APX (r = 0.76) in one-year-old seedlings (Figure 7a). Meanwhile, a moderate correlation between SOD—APX and SOD—GR was obtained (r = 0.32 and r = 0.37, respectively). The strongest correlation in two-year-old seedlings was determined between GR—CAT enzymes (r = 0.94) (Figure 7b). The correlation between SOD and other enzymes was not determined. In addition, differences between changes in POX activity compared to changes in DPPH or ABTS scavenging were determined. The moderate negative correlation between ABTS—POX (r = −0.31) was observed, while the correlation between DPPH—POX (−0.07) was not observed.
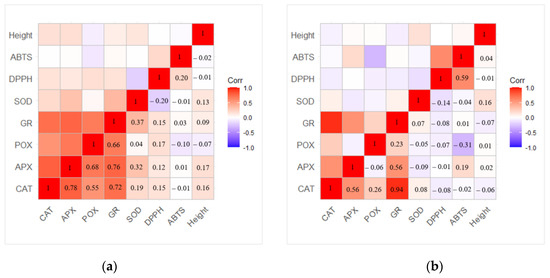
Figure 7.
Matrix of correlation between different variables in (a) one-year-old and (b) two-year-old Picea abies seedlings based on the Pearson correlation coefficient.
Figure 7.
Matrix of correlation between different variables in (a) one-year-old and (b) two-year-old Picea abies seedlings based on the Pearson correlation coefficient.
The results of PCA for data obtained on height, enzyme (CAT, APX, POX, GR, and SOD) activities, and antioxidant activity (ABTS and DPPH assays) in the needles of one-year-old and two-year-old are presented in Figure 8a and Figure 8b, respectively. The first four of the seven principal components explained 82.3% of the total variance of the parameters, while the first two—56.9% in one-year-old seedlings (Table 4). Similarly, in two-year-old seedlings, the first four principal components explained 79.5% of the total variance of the parameters, while the first two—52.5% SOD was negatively linked to PC2 in one-year-old seedlings, while ABTS and DPPH were positively linked to PC2 (Table 5). The enzymes CAT, APX, POX, and GR were grouped together and positively linked to PC1. Analyzing two-year-old seedlings, similarly to one-year-old seedlings, ABTS and DPPH were positively linked to PC2. However, POX was negatively linked to PC2 in two-year-old seedlings. Analyzing the results of PCA for data with the treatments separately, it was found that CAT and GR were grouped together and positively linked to PC1 after all the treatments in both one-year-old and two-year-old seedlings (Supplementary Figures S1 and S2). APX was grouped together with CAT and GR after all the treatments in one-year-old seedlings as well, however, in two-year-old seedlings, APX was grouped together with CAT and GR only after the treatment with CP1, the same as in the control. ABTS and DPPH were positively linked to PC2 in two-year-old seedlings after the treatments with CP1 and EMF2, the same as in the control (Supplementary Figures S1 and S2).
Table 4.
Standard deviation, proportion of variance, and cumulative proportion explained by principal components (PC1–PC7) through analysis of PCA from the data of one-year-old and two-year-old Picea abies seedlings.
Table 5.
Individual proportion of the traits to the 1st and 2nd components from the data of one-year-old and two-year-old Picea abies seedlings.
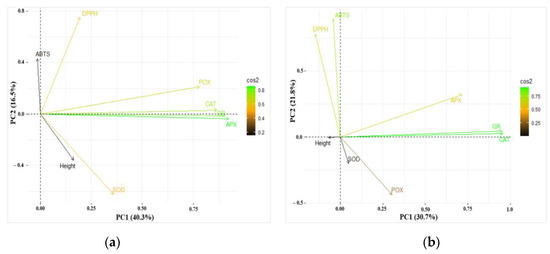
Figure 8.
Principal component analysis (PCA) displaying the first and second components (PCs) from the data of (a) one-year-old and (b) two-year-old Picea abies seedlings. The loading vectors show the correlation between different variables. Colors of vectors from black to green represent different squared cosine (Cos2) values.
Figure 8.
Principal component analysis (PCA) displaying the first and second components (PCs) from the data of (a) one-year-old and (b) two-year-old Picea abies seedlings. The loading vectors show the correlation between different variables. Colors of vectors from black to green represent different squared cosine (Cos2) values.
3. Discussion
During the long lifespans of gymnosperms, they are attacked by different pests and pathogens [45], and regularly face drought and other stressful environmental conditions [46]. To cope with potential damages induced by numerous biotic and abiotic factors, the gymnosperms have evolved many constitutive and inducible mechanisms, including the production of defensive enzymes, which act across different lifespans and types of tissues [45,47,48,49,50].
Pre-sowing plant seed treatment with cold plasma (CP) and electromagnetic field (EMF) has been proved as potential stressors eliciting complex plant response that can result in improvement of agricultural performance and stress resistance of numerous crops [51,52]. It was demonstrated that moderate stress experienced by the plasma or EMF-irradiated seed can lead to a multifaceted long-term response in the growing plant, including activated photosynthesis, secondary metabolism, and increased harvest yield. Although numerous studies have reported changes induced in the activity of the antioxidant defense system, the obtained results have been controversial [53].
Pre-sowing seed treatment with CP and EMF showed a potential to increase the resistance of forest trees as well [17,18,54,55]. However, numerous knowledge gaps related to changes induced in biochemical and physiological processes and antioxidant defense capacity in perennials remain unresolved. Thus, the purpose of this study was to compare the effects of pre-sowing treatment of P. abies seeds with CP and EMF on different half-sib families by observing changes in seedling emergence and growth of one-year-old and two-year-old seedlings, with particular attention to changes in the antioxidant system in seedling needles.
No significant effect of seed treatments with CP1, CP2, and EMF on the emergence of P. abies seedlings was observed in our study. These results confirmed earlier reported observations that the seed treatment with CP and EMF may not have a significant impact on the emergence of gymnosperm seedlings [56,57,58,59]. However, we found that pre-sowing seed treatment with physical stressors might cause a stronger impact on growth (estimated by changes in the height) of one-year-old seedlings compared to two-year-old P. abies seedlings. The treatment with CP1 and CP2 had a positive effect on 50% and 40% of half-sib families, respectively, while EMF had a negative effect on 40% of one-year-old half-sib families. Meanwhile, the significant impact of these treatments on the height of two-year-old seedlings was not determined. Competition among trees is an important driver of community structure in forests, and our results indicated that the pre-sowing seed treatments may impact tree seedling establishment and early growth [21,59].
The obtained results also demonstrated evidence that pre-sowing seed treatments with CP and EMF had the potential to cause significant changes in metabolic processes in P. abies seedlings. Catalytic proteins play an essential role in managing and stabilizing reactive oxygen species (ROS) [37]. Under environmental stress conditions, the level of ROS that causes oxidative cell damage in plants could be buffered by enzymatic antioxidants [60,61]. It was demonstrated that the accumulation of biologically active compounds in conifers depends on the tree’s age [62]. CP and EMF-induced changes in biochemical plant protection systems are age-dependent, as highlighted by higher amounts of chlorophylls and phenols in two-year-old than one-year-old P. abies seedlings [21]. The results of this study supplement such findings indicating that the activity of antioxidant enzymes is significantly higher in two-year-old than one-year-old seedlings.
One of the enzymes is superoxide dismutase (SOD), which is responsible for scavenging free radicals, particularly superoxide anion •O2−, produced during photosynthesis and other metabolic processes [38,63]. Our study showed that, unlike other enzymes, seed treatment with CP2 and EMF might cause a decrease in SOD activity in one-year-old seedlings. Meanwhile, the treatment with CP1 has the potential to increase SOD activity in two-year-old seedlings. The increase in SOD activity is primarily related to plant response to drought [64,65,66] and salinity [38,67]. Pre-sowing seed treatment with CP and EMF may have either a positive or negative effect on SOD activity, dependent on the half-sib family. Whereas SOD converts superoxide anions into H2O2, which is dangerous for plant cells, other enzymes, such as CAT, APX, POX, and GR operate in tight cooperation with SOD, since they neutralize the toxic H2O2 or restore GSH from oxidated GSSG, preventing the Haber-Weiss reaction or the Foyer–Halliwell–Asada pathway [38]. Although the reaction mechanisms of CAT/POX, APX, and GR are different, they function as elements of a single defense system against biotic and abiotic stress [38]. In our study, a strong correlation between CAT, APX, POX, and GR activities was also determined. According to other studies, water stress can increase CAT, POX, APX, and GR enzyme activity in Cucurbita pepo [68]. A similar increase in CAT and POX activities has been reported after chemical stress (various solutions of NaCl) in Berseem clover [69]. Furthermore, the application of biological treatment (jasmonic acid) can increase either CAT [70] or POX [71] activity in plants. The treatment with a magnetic field can also activate the synthesis of SOD and CAT enzymes in soybeans, indicating a strong relationship between their activities [72]. Our study provides evidence that pre-sowing seed treatment with CP and EMF can induce multiple changes in the activities of antioxidant enzymes; however, these changes were influenced by the treatment and P. abies half-sib family. For instance, in one-year-old seedlings of the 577 half-sib family, the activity of CAT, APX, POX, and GR increased after EMF-treatment, compared to the control group, while in the 477 half-sib family, CAT, APX, and GR activities increased after CP2-treatment. In two-year-old seedlings of the 548 half-sib family, the activity of CAT, APX, POX, and GR increased with the pre-sowing seed treatment with EMF. In the half-sib families 548 and 463 the activity of CAT, APX, POX, and GR had the potential to increase after pre-sowing treatment with CP1 and EMF, respectively. In the 457 half-sib family, none of the analyzed parameters (except height) changed after CP1-treatment in one-year-old seedlings.
The free radical scavenging capacity in plant extracts is commonly based on DPPH and ABTS tests [73]. It was determined that seed treatment with a magnetic field increased antioxidant activity in different sunflower extracts; however, this effect was dependent on plant variety [74]. Seed treatment with bio-stimulants enhanced ROS scavenging activity in young olive trees [75]. Pre-sowing treatment of P. abies seeds with CP and EMF also promises to increase radical scavenging activity in the needles of one-year-old and two-year-old seedlings, but it depends on the half-sib family. The results showed that both CP and EMF increased radical scavenging activity (ABTS and DPPH) in the 457 half-sib family of one-year-old and two-year-old seedlings. The differences in changes in antioxidant activity (estimated by both ABTS and DPPH assays) between the treatments with CP1 and CP2 were also observed.
In conclusion, pre-sowing seed treatment with CP and EMF has the potential to induce significant changes in the activities of antioxidant enzymes and radical scavenging in one-year-old and two-year-old P. abies seedlings, without significant impact on seedling emergence and height. The impact of pre-sowing seed treatment with CP on biochemical processes depended on the duration of treatment (1 min and 2 min). The changes induced in the activity of enzymes and radical scavenging activity were specific to different half-sib families. These treatments can be promising to apply, but for practical use, hormones and plant gene expression or other factors must be analyzed. To evaluate the impact of physical stress experienced by seeds on different P. abies half-sib families, further long-term observations are needed. Furthermore, to assess the potential application of pre-sowing seed treatments involving CP and EMF, it is necessary to test experimental models measuring changes in the antioxidant system following biotic and abiotic stress, such as drought, salinity, or attacks by pests and/or pathogens.
4. Materials and Methods
4.1. Seed Collection and Treatment with Cold Plasma (CP) and Electromagnetic Field (EMF)
Ten different half-sib families of P. abies seeds were collected from a second-generation spruce seed orchard in the Trakai regional division (Lithuania) (Table 6). The collected seeds were kept in refrigeration until the treatment. Seeds were divided into four different groups: control; seed treatment with cold plasma for 1 min (CP1), seed treatment with cold plasma for 2 min (CP2), and seed treatment with electromagnetic field for 2 min (EMF2). The total amount of seeds in experiments was 6400 (10 half-sib families × 4 treatment protocols × 160 seeds per treatment).
Table 6.
Description of Picea abies seeds plantation in Lithuania.
Seed treatment with cold plasma (CP). The seeds of each P. abies half-sib family were irradiated using a low-temperature atmospheric dielectric barrier discharge (DBD) plasma device produced at Kyushy University, Fukuota, Japan [76]. The treatment was performed at Vytautas Magnus University, Kaunas, Lithuania. Seeds were placed on a glass plate in an area of homogeneous treatment (4 × 4.38 cm2) at a distance of 5 mm between the seed surface and the electrode plate; the discharge voltage was 7.0 kV, the current was 0.2 A; the power density was 3.1 W/cm2; the humidity was 45–55%; the atmospheric pressure and room temperature [17]. The duration of treatment was 1 and 2 min.
Seed treatment with electromagnetic field (EMF). The seeds of each P. abies half-sib family were exposed to radiofrequency electromagnetic field (EMF) treatment with a pulsed magnetic field generator designed at Vilnius Gediminas Technical University (VilniusTECH), Lithuania. The seeds were treated for 2 min (parameters: 100 kHz, 400 ± 50 μT oscillating magnetic field; atmospheric pressure and room temperature) [18]. The magnetic field was measured using a calibrated loop sensor (VilniusTECH, Vilnius, Lithuania), and a DPO4034 oscilloscope (Tektronix, Beaverton, OR, USA). The peak amplitude value was also confirmed using a Gaussmeter 475DSP (Lakeshore, Carson, CA, USA).
4.2. Seedling Cultivation
Treated P. abies seeds were stored in plastic bags for 4 days at 25 °C, to allow sufficient time for alterations in the levels of phytohormones within the seeds [27]. The treated and control seeds were sown in the cassettes (40-cell) with a peat substrate (SuliFlor SF2, pH of 5.5–6.5) and lightly covered with perlite. The seedlings were cultivated in a controlled-condition greenhouse (25–32 °C daytime; >10 °C at night) for two months. Then the seedlings were transferred to an open area. After a year, the seedlings were transplanted into larger pots (18 × 17 × 2 cm) filled with 1000 mL of peat substrate (Figure 9).

Figure 9.
Treated and control one-year-old Picea abies seedlings in pots.
Figure 9.
Treated and control one-year-old Picea abies seedlings in pots.

4.3. Seedling Emergence and Height
The emergence of P. abies seedlings started to count one week after the sowing and continued for 15 days. The percentage of emerged seedlings along with the standard error was calculated [18]. P. abies seedlings height measurement (cm) was completed after the vegetation period in both one-year-old and two-year-old seedlings.
4.4. Sample Collection
The samples of P. abies needles were collected (three biological replicates) both from seedlings in the control and treated groups at the end of the first and the second vegetation season. The needle samples were selected to collect from random seedlings by taking approximately 30 needles from different analyzed groups. Fresh needle samples were partitioned into equal weight and stored at −20 °C until the start of the biochemical analyses.
4.5. Sample Preparation for Antioxidant Enzymes Analysis
P. abies needle extracts were prepared for the assessments using the method described by [18]. 0.2 g needles were powdered in liquid nitrogen and transferred to 15 mL tubes, and 5 mL of extraction buffer (containing K-phosphate buffer (pH 7.8), 1% [v/v] Triton X-100, 300 mg polyvinylpolypyrrolidone (PVPP) and 5 mM ascorbate (ASC)) solution was added. The mixture with the sample was centrifuged for 1 h, 16,090× g, +4 °C (Andreas Hettich GmbH & Co. KG, Tuttlingen, Germany). The supernatant was promptly transferred to 96 microplates for assessing the overall protein content (PROT) and measuring the activities of superoxide dismutase (SOD) and catalase (CAT). Separation of the supernatant and extract for the assessment of ascorbate peroxidase (APX), peroxidase (POX), and glutathione reductase (GR) was completed using Sephadex G-25 (Column PD-10, Cytiva, Gillingham, UK) columns on ice.
4.6. Protein Measurement and Kinetic Assays of Antioxidant Enzymes: CAT, APX, POX, GR, and SOD
All kinetic assays were performed in a 96-well microplate setup (Greiner Bio-One GmbH, Frickenhausen, Germany) using a SpectroStar Nano microplate reader (BMG Labtech, Offenburg, Germany), extract volume (20 µL), and a total assay (reaction buffer) volume (170–180 µL) in each well. Evaluation of total protein content in the needle extract was performed according to Čėsnienė et al., 2023 [18] methodology. The reaction mixture consisted of Biuret reagent and Folin–Ciocalteu reagent (1:9 w/v). The absorbance was measured at a wavelength of 660 nm. As a standard it was used Bovine Serum Albumin (BSA). The protein content in the crude extract was measured in mg/mL (the total amount of mg equivalent to BSA per milliliter).
Measurement of CAT, APX, POX, GR, and SOD activity in the needle extracts was performed according to Čėsnienė et al., 2023 [18]. The absorbance was measured at a wavelength of 240 nm, 290 nm, 430 nm, 340 nm, and 550 nm, respectively. Protein concentration in the fresh needles is used for the assessment of CAT, APX, and POX activity that was expressed in µmol H2O2 mg protein−1 min−1 in the needle biomass, as described by Wu and von Tiedeman, 2002 [76] methodology. Based on the protein concentration in the fresh needle tissue, the GR activity was measured using the oxidation of NADPH reducing oxidized L-glutathione reaction, expressed in μmol GSSG mg protein−1 min−1 in the needle biomass as described by Wu and von Tiedeman, 2002 [77] methodology. SOD activity was measured using nitroblue tetrazolium (NBT) reduction, expressed in units mg protein−1 min−1 in the needle biomass, according to Dhindsa et al., 1981 [78] methodology.
4.7. Sample Preparation for Antioxidant Activity Analysis
For detection of the antioxidant activity, both DPPH (2,2-diphenyl-1-picryl-hydrazyl-hydrate) and ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) radical scavenging tests were used according to Čėsnienė et al., 2023 [18] methodology. Needle samples (0.5 g) were homogenized, and mixed with MeOH (75%), and extracts were shaken at room temperature for 24 h, using a Kuhner Shaker X electronic shaker. After a 24-h period, the needle extracts underwent purification using Rotilabo®-113A (Ø 90 mm) filter papers.
4.8. Antioxidant Activity Test: DPPH and ABTS Radical Scavenging
Free radical scavenging activity was determined by DPPH assay in MeOH extracts of P. abies needle samples as described by Ragaee et al., 2006 [79] methodology. The DPPH reaction solution was mixed with MeOH and extract and transferred to dark for 16 min. The absorbance was measured at a wavelength of 515 nm. The DPPH radical scavenging activity was calculated as antioxidant Trolox equivalents/per gram of fresh needle material [18].
Free radicals scavenging activity was determined by ABTS assay in MeOH extracts of P. abies needle samples according to Lučinskaitė et al., 2021 [80] methodology. The ABTS reaction solution was mixed with extract and transferred to dark for 10 min. The absorbance was measured at a wavelength of 734 nm. The radical scavenging activity using ABTS assays was calculated the same as DPPH (see description above).
4.9. Statistical Analysis
Statistical analysis was performed using R (Version 4.3.1) with RStudio (Version 2023.06.0) and Microsoft Excel 2010. Column charts of antioxidant enzymes with error bars represented the mean values along with the standard error (SE), while the emergence rate, seedling height, and antioxidant system (DPPH and ABTS) were presented as means in the tables (±SE). The statistically significant differences between the two groups (control and treatment: CP1, CP2, and EMF2) were calculated using Student’s t test [81]. The Benjamini and Hochberg method was used for the adjustment of p-values for multiple comparisons [82]. In order to determine whether tested factors (treatment, year, half-sib family) had a significant influence on Picea abies antioxidant system, MANOVA was performed [83]. To compute the correlation matrix of the different components of the antioxidant system, the cor function from the corrr package was used [84]. For the visualization, the ggcorplot function from the ggplot2 package was applied [85]. To visualize the outputs of the principal component analysis (PCA) of the components of the antioxidant system, the princomp, summary, fviz_eig, fviz_pca_var, and fviz_cos2 functions from the FactoMineR package were applied [86]. Numeric differences in the mean values of different antioxidant components with different colors were visualized using the heatmap function.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants13152021/s1, Figure S1: principal component analysis (PCA) displaying the first and second components (PCs) from the data of one-year-old and two-year-old Picea abies seedlings in (A) control, (B) CP1, (C) CP2 and (D) EMF2. Figure S2. Principal component analysis (PCA) displaying the first and second components (PCs) from the data of two-year-old Picea abies seedlings in (A) control, (B) CP1, (C) CP2 and (D) EMF2.
Author Contributions
Conceptualization: I.Č., V.Č., V.S.-Š., and V.M.; Methodology: I.Č., V.S.-Š., and D.M.; Software: I.Č. and V.Č.; Validation: I.Č., V.Č., V.S.-Š., and D.M.; Formal analysis: I.Č. and V.Č.; Investigation: I.Č., V.S.-Š., V.Č., and D.M.; Resources: V.S.-Š., D.M., V.N., and V.M.; Data Curation: I.Č., V.Č., D.M., and V.M.; Writing—Original Draft: I.Č.; Writing—Review and Editing: I.Č., V.Č., V.S.-Š., V.M., V.N., and D.M.; Visualization: I.Č. and V.Č.; Supervision: I.Č. and V.S.-Š. All authors have read and agreed to the published version of the manuscript.
Funding
This research did not receive any specific funding.
Data Availability Statement
Data will be made available on request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mageroy, M.H.; Nagy, N.E.; Steffenrem, A.; Krokene, P.; Hietala, A.M. Conifer defences against pathogens and pests—Mechanisms, breeding, and management. Curr. For. Rep. 2023, 9, 429–443. [Google Scholar] [CrossRef]
- Graziosi, I.; Tembo, M.; Kuate, J.; Muchugi, A. Pests and diseases of trees in Africa: A growing continental emergency. Plants People Planet 2020, 2, 14–28. [Google Scholar] [CrossRef]
- Schuldt, B.; Buras, A.; Arend, M.; Vitasse, Y.; Beierkuhnlein, C.; Damm, A.; Gharun, M.; Grams, T.E.E.; Hauck, M.; Hajek, P.; et al. A first assessment of the impact of the extreme 2018 summer drought on Central European forests. Basic Appl. Ecol. 2020, 45, 86–103. [Google Scholar] [CrossRef]
- Teshome, D.T.; Zharare, G.E.; Naidoo, S. The threat of the combined effect of biotic and abiotic stress factors in forestry under a changing climate. Front. Plant Sci. 2020, 11, 601009. [Google Scholar] [CrossRef]
- Liebhold, A.M. Forest pest management in a changing world. Int. J. Pest. Manag. 2012, 58, 289–295. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Brockerhoff, E.G.; Wingfield, B.D.; Slippers, B. Planted forest health: The need for a global strategy. Science 2015, 349, 832–836. [Google Scholar] [CrossRef]
- Ramsfield, T.D.; Bentz, B.J.; Faccoli, M.; Jactel, H.; Brockerhoff, E.G. Forest health in a changing world: Effects of globalization and climate change on forest insect and pathogen impacts. Forestry 2016, 89, 245–252. [Google Scholar] [CrossRef]
- Hurley, B.P.; Slippers, B.; Sathyapala, S.; Wingfield, M.J. Challenges to planted forest health in developing economies. Biol. Invasions 2017, 19, 3273–3285. [Google Scholar] [CrossRef]
- Čėsna, V.; Čėsnienė, I.; Sirgedaitė-Šėžienė, V.; Marčiulynienė, D. Changes in Biologically Active Compounds in Pinus sylvestris Needles after Lymantria monacha Outbreaks and Treatment with Foray 76B. Plants 2024, 13, 328. [Google Scholar] [CrossRef]
- Čėsna, V.; Gedminas, A.; Lynikienė, J.; Marčiulynienė, D. Insect Diversity in Pinus sylvestris Forest Stands Damaged by Lymantria monacha. Insects 2024, 15, 200. [Google Scholar] [CrossRef]
- Kaur, S.; Vian, A.; Chandel, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Sensitivity of plants to high frequency electromagnetic radiation: Cellular mechanisms and morphological changes. Rev. Environ. Sci. Bio. 2021, 20, 55–74. [Google Scholar] [CrossRef]
- Bera, K.; Dutta, P.; Sadhukhan, S. Seed priming with non-ionizing physical agents: Plant responses and underlying physiological mechanisms. Plant Cell Rep. 2022, 41, 53–73. [Google Scholar] [CrossRef]
- Mildaziene, V.; Ivankov, A.; Sera, B.; Baniulis, D. Biochemical and physiological plant processes affected by seed treatment with non-thermal plasma. Plants 2022, 11, 856. [Google Scholar] [CrossRef]
- Weltmann, K.-D.; Kolb, J.; Holub, M.; Uhrlandt, D.; Simek, M.; Ostrikov, K.; Hamaguchi, S.; Cvelbar, U.; Černák, M.; Locke, B.; et al. The future for plasma science and technology. Plasma Process. Polym. 2018, 16, 1800118. [Google Scholar] [CrossRef]
- Foroughbakhch Pournavab, R.; Bacópulos Mejía, E.; Benavides Mendoza, A.; Salas Cruz, L.R.; Ngangyo Heya, M. Ultraviolet radiation effect on seed germination and seedling growth of common species from northeastern mexico. Agronomy 2019, 9, 269. [Google Scholar] [CrossRef]
- Attri, P.; Ishikawa, K.; Okumura, T.; Koga, K.; Shiratani, M. Plasma agriculture from laboratory to farm: A review. Processes 2020, 8, 1002. [Google Scholar] [CrossRef]
- Sirgedaitė-Šėžienė, V.; Lučinskaitė, I.; Mildažienė, V.; Ivankov, A.; Koga, K.; Shiratani, M.; Laužikė, K.; Baliuckas, V. Changes in content of bioactive compounds and antioxidant activity induced in needles of different half-sib families of norway spruce (Picea abies (L.) H. Karst) by seed treatment with cold plasma. Antioxidants 2022, 11, 1558. [Google Scholar] [CrossRef] [PubMed]
- Čėsnienė, I.; Miškelytė, D.; Novickij, V.; Mildažienė, V.; Sirgedaitė-Šėžienė, V. Seed treatment with electromagnetic field induces different effects on emergence, growth and profiles of biochemical compounds in seven half-sib families of silver birch. Plants 2023, 12, 3048. [Google Scholar] [CrossRef] [PubMed]
- Stranak, V.; Tichy, M.; Kriha, V.; Scholtz, V.; Sera, B.; Spatenka, P. Technological applications of surfatron produced discharge. J. Optoelectron. Adv. Mater. 2007, 9, 852–857. [Google Scholar]
- Leti, L.I.; Gerber, I.C.; Mihaila, I.; Galan, P.M.; Strajeru, S.; Petrescu, D.E.; Cimpeanu, M.M.; Topala, I.; Gorgan, D.L. The modulatory effects of non-thermal plasma on seed’s morphology, germination and genetics—A review. Plants 2022, 11, 2181. [Google Scholar] [CrossRef] [PubMed]
- Sirgedaitė-Šėžienė, V.; Mildažienė, V.; Žemaitis, P.; Ivankov, A.; Koga, K.; Shiratani, M.; Baliuckas, V. Long-term response of Norway spruce to seed treatment with cold plasma: Dependence of the effects on the genotype. Plasma Process. Polym. 2021, 18, 2000159. [Google Scholar] [CrossRef]
- Pawełek, A.; Wyszkowska, J.; Cecchetti, D.; Dinka, M.D.; Przybylski, K.; Szmidt-Jaworska, A. The physiological and biochemical response of field bean (Vicia faba L. (partim)) to electromagnetic field exposure is influenced by seed age, light conditions, and growth media. Agronomy 2022, 12, 2161. [Google Scholar] [CrossRef]
- Maffei, M.E. Magnetic field effects on plant growth, development, and evolution. Front. Plant Sci. 2014, 5, 445. [Google Scholar] [CrossRef]
- Pietruszewski, S.; Martínez, E. Magnetic field as a method of improving the quality of sowing material: A review. Int. Agrophys 2015, 29, 377–389. [Google Scholar] [CrossRef]
- Randeniya, L.K.; de Groot, G. Non-thermal plasma treatment of agricultural seeds for stimulation of germination, removal of surface contamination and other benefits: A review. Plasma Process. Polym. 2015, 12, 608–623. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A.; Dobránszki, J. Magnetic fields: How is plant growth and development impacted? Protoplasma 2016, 253, 231–248. [Google Scholar] [CrossRef]
- Degutytė-Fomins, L.; Paužaitė, G.; Žūkienė, R.; Mildažienė, V.; Koga, K.; Shiratani, M. Relationship between cold plasma treatment-induced changes in radish seed germination and phytohormone balance. Jpn. J. Appl. Phys. 2020, 59, SH1001. [Google Scholar] [CrossRef]
- Beniušytė, E.; Čėsnienė, I.; Sirgedaitė-Šėžienė, V.; Vaitiekūnaitė, D. Genotype-dependent jasmonic acid effect on Pinus sylvestris L. Growth and induced systemic resistance indicators. Plants 2023, 12, 255. [Google Scholar] [CrossRef]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef]
- Dvořák, P.; Krasylenko, Y.; Zeiner, A.; Šamaj, J.; Takáč, T. Signaling toward reactive oxygen species-scavenging enzymes in plants. Front. Plant Sci. 2021, 11, 618835. [Google Scholar] [CrossRef] [PubMed]
- Pospíšil, P. Production of reactive oxygen species by photosystem ii as a response to light and temperature stress. Front. Plant Sci. 2016, 7, 1950. [Google Scholar] [CrossRef] [PubMed]
- Del Río, L.A.; López-Huertas, E. ROS generation in peroxisomes and its role in cell signaling. Plant Cell Physiol. 2016, 57, 1364–1376. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Serrano, M.; L’Haridon, F.; Tjamos, S.E.; Metraux, J.P. Reactive oxygen species and plant resistance to fungal pathogens. Phytochemistry 2015, 112, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Eltayeb, A.E.; Kawano, N.; Badawi, G.H.; Kaminaka, H.; Sanekata, T.; Shibahara, T.; Inanaga, S.; Tanaka, K. Overexpression of monodehydroascorbate reductase in transgenic tobacco confers enhanced tolerance to ozone, salt and polyethylene glycol stresses. Planta 2007, 225, 1255–1264. [Google Scholar] [CrossRef] [PubMed]
- Arena, C.; Vitale, L.; Bianchi, A.R.; Mistretta, C.; Vitale, E.; Parisi, C.; Guerriero, G.; Magliulo, V.; De Maio, A. The ageing process affects the antioxidant defences and the poly (adpribosyl) ation activity in Cistus incanus L. Leaves. Antioxidants 2019, 8, 528. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; Mandzhieva, S. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Fimognari, L.; Dölker, R.; Kaselyte, G.; Jensen, C.; Akhtar, S.; Großkinsky, D.; Roitsch, T. Simple semi-high throughput determination of activity signatures of key antioxidant enzymes for physiological phenotyping. Plant Methods 2020, 16, 42. [Google Scholar] [CrossRef]
- Caverzan, A.; Passaia, G.; Barcellos Rosa, S.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef]
- Thor, M.; Stenlid, J. Heterobasidion annosum infection of Picea abies following manual or mechanized stump treatment. Scand. J. For. Res. 2005, 20, 154–164. [Google Scholar] [CrossRef]
- Oliva, J.; Bendz-Hellgren, M.; Stenlid, J. Spread of Heterobasidion annosum s.s. and Heterobasidion parviporum in Picea abies 15 years after stump inoculation. FEMS Microb. Ecol. 2011, 75, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Marčiulynas, A.; Sirgedaitė-Šėžienė, V.; Žemaitis, P.; Baliuckas, V. The resistance of scots pine (Pinus sylvestris L.) Half-sib families to Heterobasidion annosum. Forests 2019, 10, 287. [Google Scholar] [CrossRef]
- Striganavičiūtė, G.; Žiauka, J.; Sirgedaitė-Šėžienė, V.; Vaitiekūnaitė, D. Priming of resistance-related phenolics: A study of plant-associated bacteria and Hymenoscyphus fraxineus. Microorganisms 2021, 9, 2504. [Google Scholar] [CrossRef]
- Fossdal, C.G.; Krokene, P.; Olsen, J.E.; Strimbeck, R.; Viejo, M.; Yakovlev, I.; Mageroy, M.H. Epigenetic stress memory in gymnosperms. Plant Physiol. 2024, 195, 1117–1133. [Google Scholar] [CrossRef]
- Li, X.; Piao, S.; Wang, K.; Wang, X.; Wang, T.; Ciais, P.; Chen, A.; Lian, X.; Peng, S.; Peñuelas, J. Temporal trade-off between gymnosperm resistance and resilience increases forest sensitivity to extreme drought. Nat. Ecol. Evol. 2020, 4, 1075–1083. [Google Scholar] [CrossRef]
- Krokene, P. Chapter 5—Conifer Defense and Resistance to Bark Beetles. In Bark Beetles; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 177–207. [Google Scholar] [CrossRef]
- Anand, A.; Surendran, V.; Sukumaran, S. Bioactive metabolites in gymnosperms. In Plant Metabolites: Methods, Applications and Prospects; Springer: Singapore, 2020; pp. 17–346. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, S.; Li, L.; Yang, T.; Dong, S.; Wei, T.; Wu, S.; Liu, Y.; Gong, Y.; Feng, X.; et al. The Cycas genome and the early evolution of seed plants. Nat. Plants 2022, 8, 389–401. [Google Scholar] [CrossRef]
- Yan, Z.; Tian, F.; Wu, C.; Yan, S.; Kou, X.; Zhou, D.; Fan, G.; Li, T.; Bai, C. Transcriptome analysis of the related defense genes in postharvest Ginkgo biloba seeds against Penicillium oxalicum. Physiol. Mol. Plant Pathol. 2023, 127, 102060. [Google Scholar] [CrossRef]
- Ivankov, A.; Naučienė, Z.; Degutytė-Fomins, L.; Žūkienė, R.; Januškaitienė, I.; Malakauskienė, A.; Jakštas, V.; Ivanauskas, L.; Romanovskaja, D.; Šlepetienė, A.; et al. Changes in agricultural performance of common buckwheat induced by seed treatment with cold plasma and electromagnetic field. Appl. Sci. 2021, 11, 4391. [Google Scholar] [CrossRef]
- Pańka, D.; Jeske, M.; Łukanowski, A.; Baturo-Cieśniewska, A.; Prus, P.; Maitah, M.; Maitah, K.; Malec, K.; Rymarz, D.; Muhire, J.; et al. Can cold plasma be used for boosting plant growth and plant protection in sustainable plant production? Agronomy 2022, 12, 841. [Google Scholar] [CrossRef]
- Holubová, Ľ.; Kyzek, S.; Ďurovcová, I.; Fabová, J.; Horváthová, E.; Ševčovičová, A.; Gálová, E. Non-Thermal Plasma—A New Green Priming Agent for Plants? Int. J. Mol. Sci. 2020, 21, 9466. [Google Scholar] [CrossRef] [PubMed]
- Pauzaite, G.; Malakauskiene, A.; Nauciene, Z.; Zukiene, R.; Filatova, I.; Lyushkevich, V.; Azarko, I.; Mildaziene, V. Changes in Norway spruce germination and growth induced by pre-sowing seed treatment with cold plasma and electromagnetic field: Short-term versus long-term effects. Plasma Process. Polym. 2018, 15, 1700068. [Google Scholar] [CrossRef]
- Šerá, B.; Šerý, M.; Zahoranová, A.; Tomeková, J. Germination improvement of three pine species (Pinus) after diffuse coplanar surface barrier discharge plasma treatment. Plasma Chem. Plasma Process. 2021, 41, 211–226. [Google Scholar] [CrossRef]
- Meiqiang, Y.; Mingjing, H.; Buzhou, M.; Tengcai, M. Stimulating effects of seed treatment by magnetized plasma on tomato growth and yield. Plasma Sci. Technol. 2005, 7, 3143. [Google Scholar] [CrossRef]
- Zukiene, R.; Nauciene, Z.; Januskaitiene, I.; Pauzaite, G.; Mildaziene, V.; Koga, K.; Shiratani, M. Dielectric barrier discharge plasma treatment-induced changes in sunflower seed germination, phytohormone balance, and seedling growth. Appl. Phys. Express 2019, 12, 126003. [Google Scholar] [CrossRef]
- Švubová, R.; Kyzek, S.; Medvecká, V.; Slováková, Ľ.; Gálová, E.; Zahoranová, A. Novel insight at the effect of cold atmospheric pressure plasma on the activity of enzymes essential for the germination of pea (Pisum sativum L. cv. Prophet) seeds. Plasma Chem. Plasma Process. 2020, 40, 1221–1240. [Google Scholar] [CrossRef]
- Mildaziene, V.; Pauzaite, G.; Malakauskiene, A.; Zukiene, R.; Nauciene, Z.; Filatova, I.; Azharonok, V.; Lyushkevich, V. Response of perennial woody plants to seed treatment by electromagnetic field and low-temperature plasma. Bioelectromagnetics 2016, 37, 536–548. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxygen processing in photosynthesis: Regulation and signalling. New Phytol. 2020, 146, 359–388. [Google Scholar] [CrossRef]
- Žaltauskaitė, J.; Dikšaitytė, A.; Miškelytė, D.; Kacienė, G.; Sujetovienė, G.; Januškaitienė, I.; Juknys, R. Effects of elevated CO2 concentration and temperature on the mixed-culture grown wild mustard (Sinapis arvensis L.) response to auxin herbicide. Environ. Sci. Pollut. Res. 2023, 30, 13711–13725. [Google Scholar] [CrossRef]
- Tegischer, K.; Tausz, M.; Wieser, G.; Grill, D. Tree- and needle-age-dependent variations in antioxidants and photoprotective pigments in Norway spruce needles at the alpine timberline. Tree Physiol. 2002, 22, 591–596. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul. 2005, 46, 209–221. [Google Scholar] [CrossRef]
- Sharma, P.; Shanker Dubey, R. Modulation of nitrate reductase activity in rice seedlings under aluminium toxicity and water stress: Role of osmolytes as enzyme protectant. J. Plant Physiol. 2005, 162, 854–864. [Google Scholar] [CrossRef]
- Zlatev, Z.S.; Lidon, F.C.; Ramalho, J.C.; Yordanov, I.T. Comparison of resistance to drought of three bean cultivars. Biol. Plant 2006, 50, 389–394. [Google Scholar] [CrossRef]
- Gapińska, M.; Skłodowska, M.; Gabara, B. Effect of short- and long-term salinity on the activities of antioxidative enzymes and lipid peroxidation in tomato roots. Acta Physiol. Plant 2008, 30, 11–18. [Google Scholar] [CrossRef]
- Biareh, V.; Shekari, F.; Sayfzadeh, S.; Zakerin, H.; Hadidi, E.; Beltrão, J.G.T.; Mastinu, A. Physiological and qualitative response of Cucurbita pepo L. To salicylic acid under controlled water stress conditions. Horticulturae 2022, 8, 79. [Google Scholar] [CrossRef]
- Rouhi, H.; Aboutalebian, M.; Moosavi, S.; Karimi, F.; Karimi, F.; Saman, M.; Samadi, M. Change in several antioxidant enzymes activity of Berseem clover (Trifolium alexandrinum L.) by priming. Int. J. Agric. Sci. 2012, 2, 237–243. [Google Scholar]
- Guan, L.M.; Scandalios, J.G. Hydrogen peroxide-mediated catalase gene expression in response to wounding. Free Radic. Biol. Med. 2000, 28, 1182–1190. [Google Scholar] [CrossRef]
- Gould, N.; Reglinski, T.; Northcott, G.L.; Spiers, M.; Taylor, J.T. Physiological and biochemical responses in Pinus radiata seedlings associated with methyl jasmonate-induced resistance to Diplodia pinea. Physiol. Mol. Plant Pathol. 2009, 74, 121–128. [Google Scholar] [CrossRef]
- Çelik, Ö.; Büyükuslu, N.; Atak, C.; Rzakoulieva, A. Effects of Magnetic Field on Activity of Superoxide Dismutase and Catalase in Glycine max (L.) Merr. Roots. Pol. J. Environ. Stud. 2009, 18, 175–182. [Google Scholar]
- Ferreira-Santos, P.; Genisheva, Z.; Pereira, R.N.; Teixeira, J.A.; Rocha, C.M.R. Moderate electric fields as a potential tool for sustainable recovery of phenolic compounds from Pinus pinaster bark. ACS Sustain. Chem. Eng. 2019, 7, 8816–8826. [Google Scholar] [CrossRef]
- Bukhari, S.A.; Tanveer, M.; Mustafa, G.; Zia-Ud-Den, N. Magnetic field stimulation effect on germination and antioxidant activities of presown hybrid seeds of sunflower and its seedlings. J. Food Qual. 2021, 2021, 5594183. [Google Scholar] [CrossRef]
- Graziani, G.; Cirillo, A.; Giannini, P.; Conti, S.; El-Nakhel, C.; Rouphael, Y.; Ritieni, A.; Di Vaio, C. Biostimulants improve plant growth and bioactive compounds of young olive trees under abiotic stress conditions. Agriculture 2022, 12, 227. [Google Scholar] [CrossRef]
- Sarinont, T.; Amano, T.; Attri, P.; Koga, K.; Hayashi, N.; Shiratani, M. Effects of plasma irradiation using various feeding gases on growth of Raphanus sativus L. Arch. Biochem. Biophys. 2016, 605, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; von Tiedemann, A. Impact of fungicides on active oxygen species and antioxidant enzymes in spring barley (Hordeum vulgare L.) exposed to ozone. Environ. Pollut. 2002, 116, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Ragaee, S.; Abdel-Aal, E.-S.M.; Noaman, M. Antioxidant activity and nutrient composition of selected cereals for food use. Food Chem. 2006, 98, 32–38. [Google Scholar] [CrossRef]
- Lučinskaitė, I.; Laužikė, K.; Žiauka, J.; Baliuckas, V.; Čėsna, V.; Sirgedaitė-Šėžienė, V. Assessment of biologically active compounds, organic acids and antioxidant activity in needle extracts of different Norway spruce (Picea abies (L.) H. Karst) half-sib families. Wood Sci. Technol. 2021, 55, 1221–1235. [Google Scholar] [CrossRef]
- Student. The Probable Error of a Mean. Biometrika 1908, 6, 1–25. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Keselman, H.J.; Huberty, C.J.; Lix, L.M.; Olejnik, S.; Cribbie, R.A.; Donahue, B.; Levin, J.R. Statistical practices of educational researchers: An analysis of their ANOVA, MANOVA, and ANCOVA analyses. Rev. Educ. Res. 1998, 68, 350–386. [Google Scholar] [CrossRef]
- Kuhn, M.; Jackson, S.; Cimentada, J. Corrr: Correl. R. Available online: https://github.com/tidymodels/corrr (accessed on 2 March 2023).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).