
Seed Treatment with Cold Plasma and Electromagnetic Field: Changes in Antioxidant Capacity of Seedlings in Different Picea abies (L.) H. Karst Half-Sib Families

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Emergence of P. abies Seedlings
2.2. Height of P. abies Seedlings
2.3. Changes of Activity of Antioxidant Enzymes in Needles of P. abies
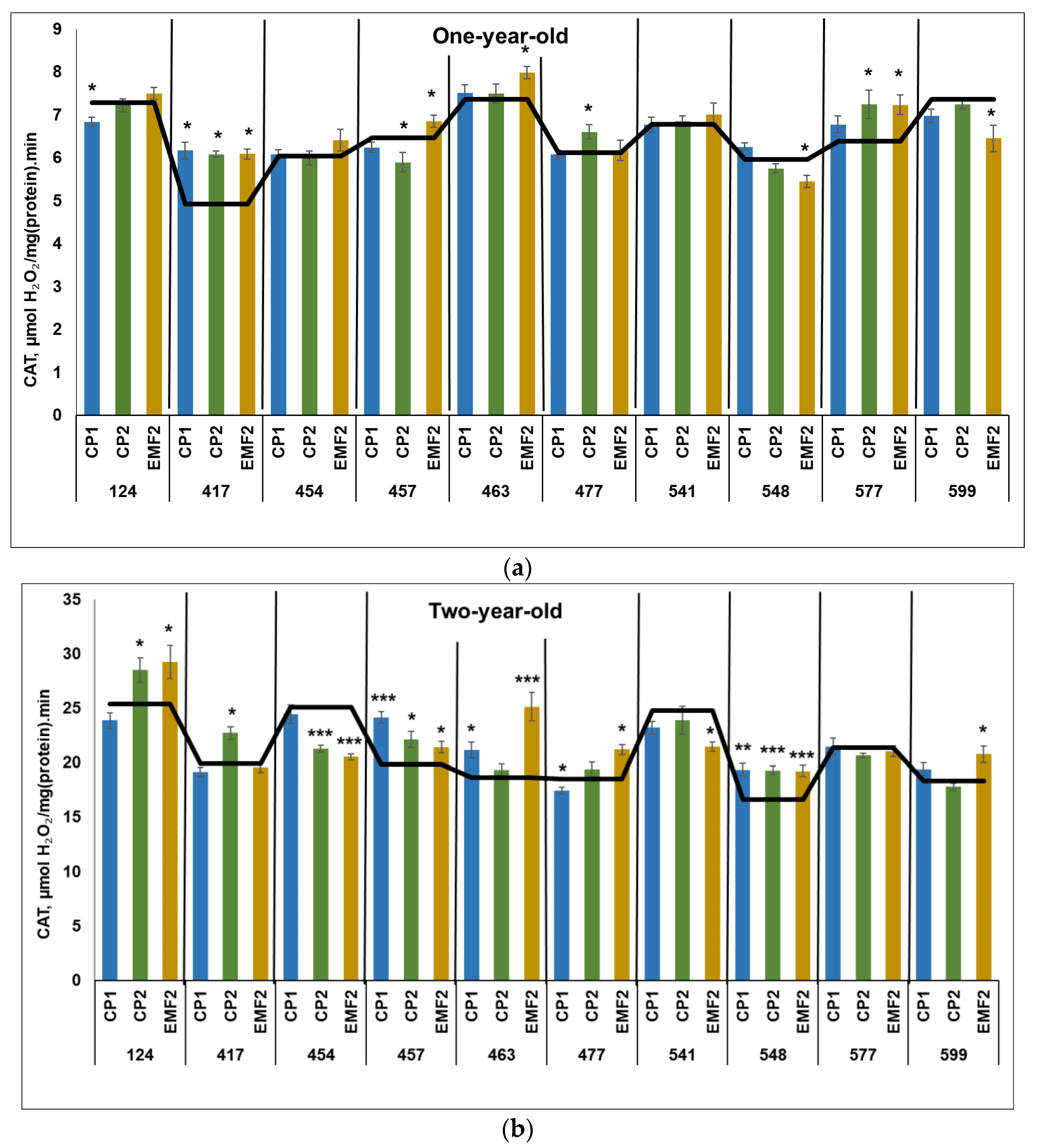
2.3.1. Catalase (CAT)
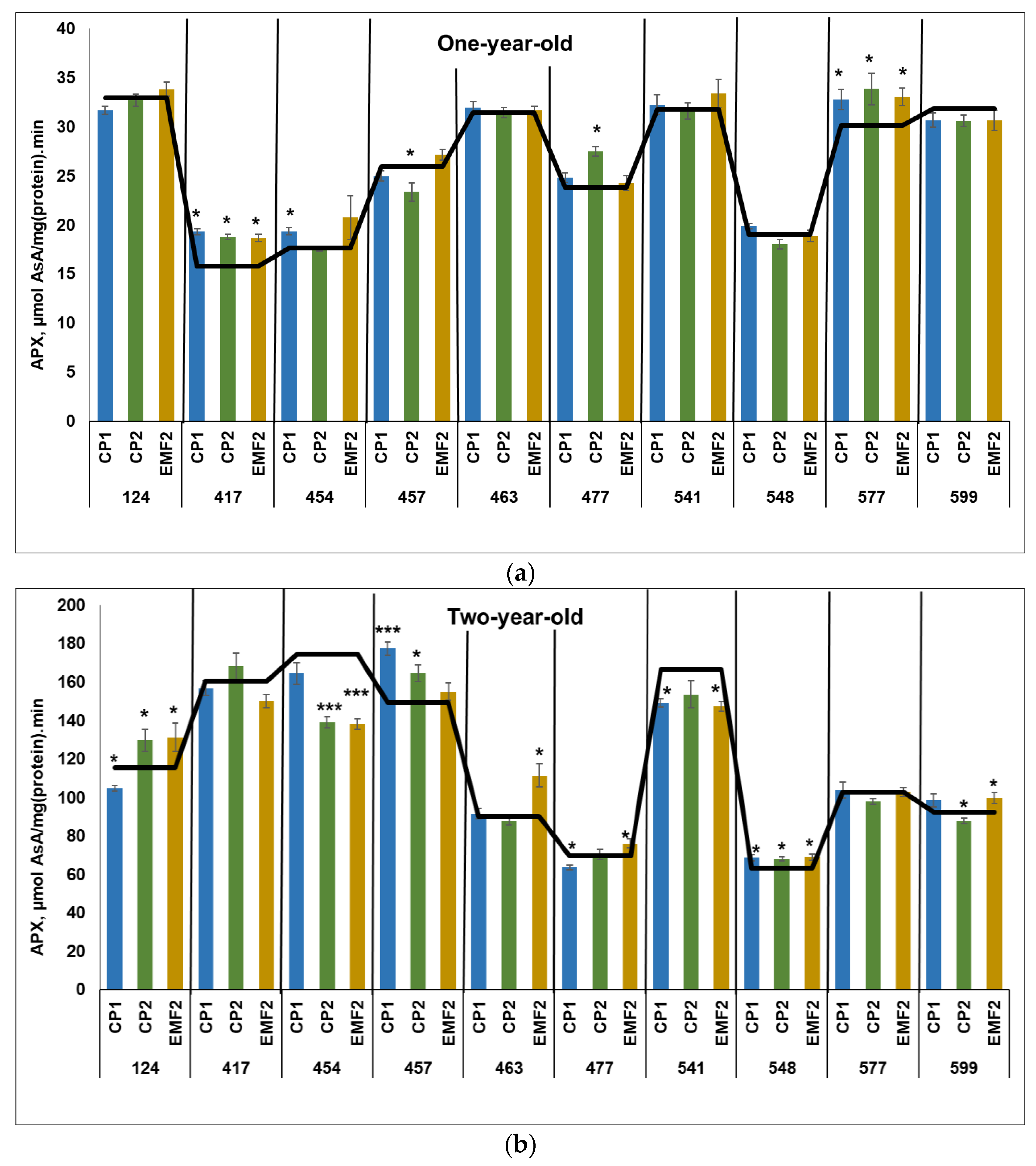
2.3.2. Ascorbate Peroxidase (APX)
2.3.3. Peroxidase (POX)

2.3.4. Glutathione Reductase (GR)

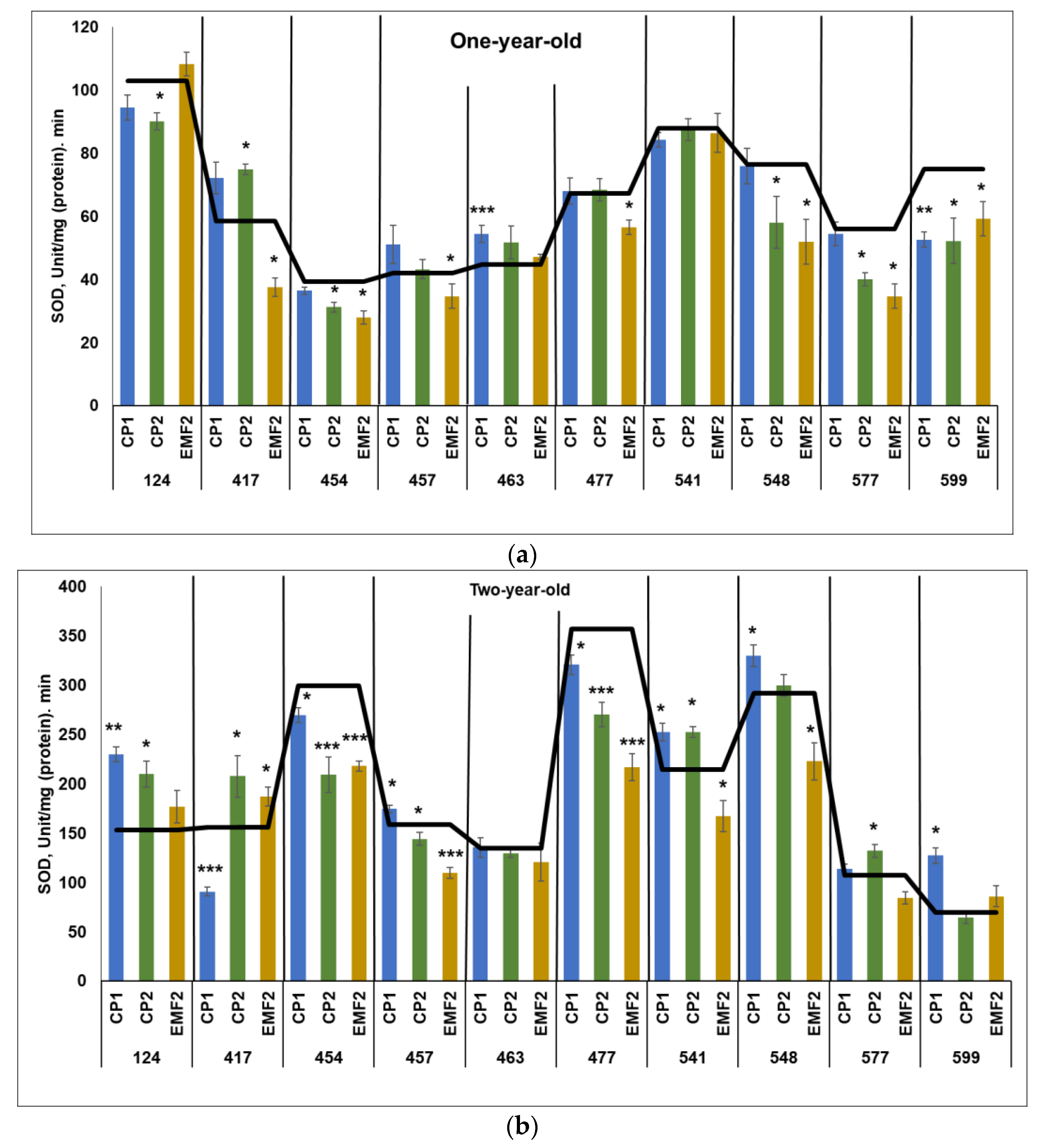
2.3.5. Superoxide Dismutase (SOD)
2.4. Changes of Antioxidant Activity in P. abies Needles

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Half-Sib Family | Radical Scavenging Activity (µmol/g) ± SE One-Year-Old | |||||||
| DPPH Assay | ABTS Assay | |||||||
| C | CP1 | CP2 | EMF2 | C | CP1 | CP2 | EMF2 | |
| 124 | 78.0 ± 5.7 | 81.3 ± 8.5 | 75.8 ± 10.6 | 76.3 ± 6.6 | 591.4 ± 24.3 | 574.6 ± 22.6 | 629.7 ± 18.0 | 606.8 ± 14.8 |
| 417 | 86.9 ± 9.2 | 72.8 ± 11.7 | 51.3 ± 7.1 * | 90.5 ± 8.8 | 790.9 ± 21.9 | 734.5 ± 14.6 * | 642.9 ± 20.6 *** | 805.6 ± 33.8 |
| 454 | 78.0 ± 7.6 | 63.3 ± 6.8 | 61.9 ± 13.6 | 69.7 ± 7.7 | 471.7 ± 12.2 | 517.4 ± 18.6 * | 586.2 ± 30.4 ** | 639.0 ± 22.4 *** |
| 457 | 72.8 ± 6.3 | 88.4 ± 8.2 | 85.7 ± 6.3 | 80.4 ± 7.3 | 610.9 ± 26.9 | 641.0 ± 6.1 | 616.2 ± 17.9 | 549.4 ± 21.4 |
| 463 | 72.6 ± 6.6 | 75.8 ± 12.2 | 93.0 ± 7.6 * | 92.3 ± 6.0 * | 539.0 ± 10.6 | 635.6 ± 11.2 *** | 622.0 ± 13.3 *** | 592.4 ± 10.0 * |
| 477 | 71.1 ± 10.7 | 71.5 ± 6.1 | 65.2 ± 8.9 | 70.8 ± 8.2 | 475.6 ± 20.1 | 462.2 ± 18.2 | 534.4 ± 33.5 | 579.8 ± 10.7 *** |
| 541 | 68.2 ± 8.8 | 48.7 ± 6.7 | 68.8 ± 5.8 | 65.0 ± 6.4 | 712.8 ± 81.9 | 573.9 ± 17.6 | 695.6 ± 12.2 | 613.3 ± 12.7 |
| 548 | 83.9 ± 4.7 | 64.7 ± 6.4 * | 75.1 ± 5.3 | 78.5 ± 6.0 | 559.9 ± 12.9 | 549.5 ± 22.7 | 616.5 ± 14.2 * | 628.2 ± 19.3 * |
| 577 | 80.8 ± 7.3 | 104.2 ± 6.6 * | 95.0 ± 7.2 | 89.0 ± 9.6 | 614.7 ± 17.2 | 576.3 ± 16.0 | 600.0 ± 12.1 | 596.0 ± 20.5 |
| 599 | 73.4 ± 12.0 | 81.0 ± 10.7 | 89.5 ± 6.1 | 83.2 ± 7.6 | 733.1 ± 26.1 | 803.8 ± 30.0 | 718.1 ± 18.6 | 662.3 ± 22.8 * |
| (b) | ||||||||
| Half-Sib Family | Radical Scavenging Activity (µmol/g) ± SE Two-Year-Old | |||||||
| DPPH Assay | ABTS Assay | |||||||
| C | CP1 | CP2 | EMF2 | C | CP1 | CP2 | EMF2 | |
| 124 | 587.8 ± 13.3 | 633.9 ± 9.7 * | 560.6 ± 10.6 | 601.7 ± 20.8 | 1418.3 ± 24.8 | 1397.7 ± 29.1 | 1206.2 ± 27.6 *** | 1243.1 ± 25.0 *** |
| 417 | 650.6 ± 10.7 | 599.9 ± 14.9 * | 614.6 ± 9.8 * | 681.4 ± 14.8 | 1402.0 ± 20.9 | 1322.1 ± 24.0 * | 1324.0 ± 17.1 * | 1468.6 ± 31.4 |
| 454 | 563.4 ± 19.5 | 579.3 ± 11.6 | 559.6 ± 9.0 | 542.8 ± 14.2 | 1117.1 ± 14.3 | 1199.0 ± 35.6 * | 1099.5 ± 17.6 | 1094.6 ± 23.4 |
| 457 | 559.2 ± 16.7 | 616.4 ± 18.1 * | 712.2 ± 19.1 *** | 660.5 ± 10.8 *** | 1283.1 ± 22.6 | 1364.1 ± 28.4 * | 1510.3 ± 40.5 *** | 1375.8 ± 22.9 * |
| 463 | 584.6 ± 13.4 | 569.8 ± 15.7 | 613.1 ± 16.3 | 566.9 ± 16.6 | 1053.4 ± 17.2 | 1122.9 ± 21.4 * | 1230.4 ± 32.7 *** | 996.8 ± 20.9 * |
| 477 | 617.6 ± 12.2 | 668.0 ± 20.1 * | 594.7 ± 12.2 | 774.0 ± 43.6 ** | 1339.7 ± 23.2 | 1366.1 ± 27.5 | 1169.3 ± 36.3 | 1557.1 ± 83.8 |
| 541 | 581.9 ± 13.4 | 605.1 ± 14.8 | 537.9 ± 10.0 * | 559.7 ± 10.4 | 1159.1 ± 24.5 | 1140.1 ± 19.1 | 1203.1 ± 26.6 | 1375.6 ± 43.3 *** |
| 548 | 586.7 ± 15.1 | 537.5 ± 14.4 * | 546.2 ± 12.8 * | 573.7 ± 20.3 | 1125.3 ± 42.5 | 1097.4 ± 27.2 | 1056.2 ± 39.0 | 1079.3 ± 37.4 |
| 577 | 557.7 ± 13.3 | 551.2 ± 12.6 | 626.1 ± 20.8 * | 469.4 ± 13.2 *** | 1058.4 ± 13.5 | 1035.4 ± 26.0 | 1064.5 ± 28.6 | 796.1 ± 17.1 *** |
| 599 | 662.0 ± 16.1 | 607.5 ± 17.0 * | 678.3 ± 11.7 | 660.3 ± 17.9 | 1409.9 ± 35.7 | 1266.1 ± 33.1 ** | 1308.5 ± 22.9 * | 1367.4 ± 19.7 |
2.5. Heatmap, Correlation, and Principal Component Analysis (PCA) between Antioxidant Enzymes and Antioxidant Activity

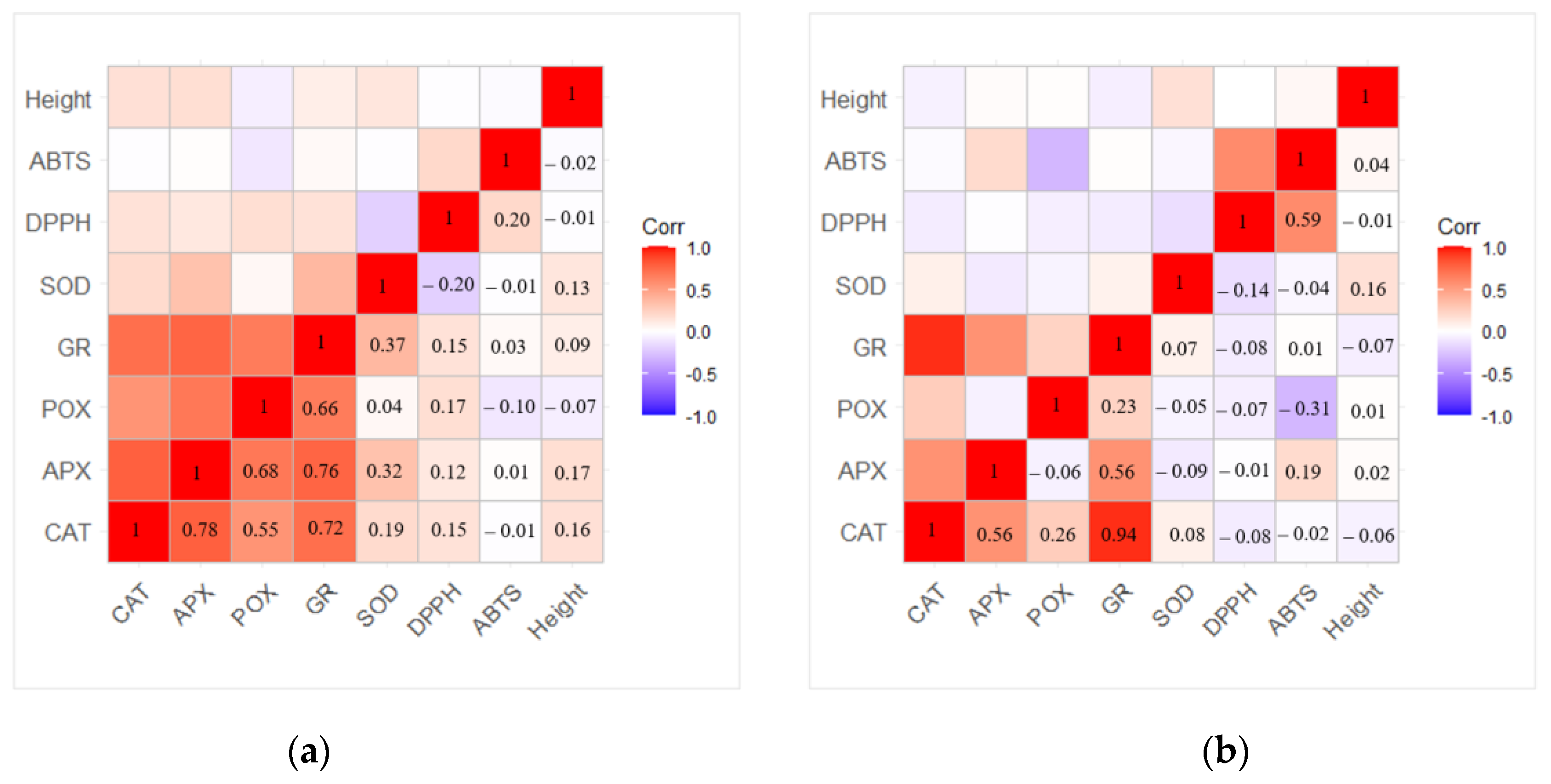

3. Discussion
4. Materials and Methods
4.1. Seed Collection and Treatment with Cold Plasma (CP) and Electromagnetic Field (EMF)
4.2. Seedling Cultivation

4.3. Seedling Emergence and Height
4.4. Sample Collection
4.5. Sample Preparation for Antioxidant Enzymes Analysis
4.6. Protein Measurement and Kinetic Assays of Antioxidant Enzymes: CAT, APX, POX, GR, and SOD
4.7. Sample Preparation for Antioxidant Activity Analysis
4.8. Antioxidant Activity Test: DPPH and ABTS Radical Scavenging
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mageroy, M.H.; Nagy, N.E.; Steffenrem, A.; Krokene, P.; Hietala, A.M. Conifer defences against pathogens and pests—Mechanisms, breeding, and management. Curr. For. Rep. 2023, 9, 429–443. [Google Scholar] [CrossRef]
- Graziosi, I.; Tembo, M.; Kuate, J.; Muchugi, A. Pests and diseases of trees in Africa: A growing continental emergency. Plants People Planet 2020, 2, 14–28. [Google Scholar] [CrossRef]
- Schuldt, B.; Buras, A.; Arend, M.; Vitasse, Y.; Beierkuhnlein, C.; Damm, A.; Gharun, M.; Grams, T.E.E.; Hauck, M.; Hajek, P.; et al. A first assessment of the impact of the extreme 2018 summer drought on Central European forests. Basic Appl. Ecol. 2020, 45, 86–103. [Google Scholar] [CrossRef]
- Teshome, D.T.; Zharare, G.E.; Naidoo, S. The threat of the combined effect of biotic and abiotic stress factors in forestry under a changing climate. Front. Plant Sci. 2020, 11, 601009. [Google Scholar] [CrossRef]
- Liebhold, A.M. Forest pest management in a changing world. Int. J. Pest. Manag. 2012, 58, 289–295. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Brockerhoff, E.G.; Wingfield, B.D.; Slippers, B. Planted forest health: The need for a global strategy. Science 2015, 349, 832–836. [Google Scholar] [CrossRef]
- Ramsfield, T.D.; Bentz, B.J.; Faccoli, M.; Jactel, H.; Brockerhoff, E.G. Forest health in a changing world: Effects of globalization and climate change on forest insect and pathogen impacts. Forestry 2016, 89, 245–252. [Google Scholar] [CrossRef]
- Hurley, B.P.; Slippers, B.; Sathyapala, S.; Wingfield, M.J. Challenges to planted forest health in developing economies. Biol. Invasions 2017, 19, 3273–3285. [Google Scholar] [CrossRef]
- Čėsna, V.; Čėsnienė, I.; Sirgedaitė-Šėžienė, V.; Marčiulynienė, D. Changes in Biologically Active Compounds in Pinus sylvestris Needles after Lymantria monacha Outbreaks and Treatment with Foray 76B. Plants 2024, 13, 328. [Google Scholar] [CrossRef]
- Čėsna, V.; Gedminas, A.; Lynikienė, J.; Marčiulynienė, D. Insect Diversity in Pinus sylvestris Forest Stands Damaged by Lymantria monacha. Insects 2024, 15, 200. [Google Scholar] [CrossRef]
- Kaur, S.; Vian, A.; Chandel, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Sensitivity of plants to high frequency electromagnetic radiation: Cellular mechanisms and morphological changes. Rev. Environ. Sci. Bio. 2021, 20, 55–74. [Google Scholar] [CrossRef]
- Bera, K.; Dutta, P.; Sadhukhan, S. Seed priming with non-ionizing physical agents: Plant responses and underlying physiological mechanisms. Plant Cell Rep. 2022, 41, 53–73. [Google Scholar] [CrossRef]
- Mildaziene, V.; Ivankov, A.; Sera, B.; Baniulis, D. Biochemical and physiological plant processes affected by seed treatment with non-thermal plasma. Plants 2022, 11, 856. [Google Scholar] [CrossRef]
- Weltmann, K.-D.; Kolb, J.; Holub, M.; Uhrlandt, D.; Simek, M.; Ostrikov, K.; Hamaguchi, S.; Cvelbar, U.; Černák, M.; Locke, B.; et al. The future for plasma science and technology. Plasma Process. Polym. 2018, 16, 1800118. [Google Scholar] [CrossRef]
- Foroughbakhch Pournavab, R.; Bacópulos Mejía, E.; Benavides Mendoza, A.; Salas Cruz, L.R.; Ngangyo Heya, M. Ultraviolet radiation effect on seed germination and seedling growth of common species from northeastern mexico. Agronomy 2019, 9, 269. [Google Scholar] [CrossRef]
- Attri, P.; Ishikawa, K.; Okumura, T.; Koga, K.; Shiratani, M. Plasma agriculture from laboratory to farm: A review. Processes 2020, 8, 1002. [Google Scholar] [CrossRef]
- Sirgedaitė-Šėžienė, V.; Lučinskaitė, I.; Mildažienė, V.; Ivankov, A.; Koga, K.; Shiratani, M.; Laužikė, K.; Baliuckas, V. Changes in content of bioactive compounds and antioxidant activity induced in needles of different half-sib families of norway spruce (Picea abies (L.) H. Karst) by seed treatment with cold plasma. Antioxidants 2022, 11, 1558. [Google Scholar] [CrossRef] [PubMed]
- Čėsnienė, I.; Miškelytė, D.; Novickij, V.; Mildažienė, V.; Sirgedaitė-Šėžienė, V. Seed treatment with electromagnetic field induces different effects on emergence, growth and profiles of biochemical compounds in seven half-sib families of silver birch. Plants 2023, 12, 3048. [Google Scholar] [CrossRef] [PubMed]
- Stranak, V.; Tichy, M.; Kriha, V.; Scholtz, V.; Sera, B.; Spatenka, P. Technological applications of surfatron produced discharge. J. Optoelectron. Adv. Mater. 2007, 9, 852–857. [Google Scholar]
- Leti, L.I.; Gerber, I.C.; Mihaila, I.; Galan, P.M.; Strajeru, S.; Petrescu, D.E.; Cimpeanu, M.M.; Topala, I.; Gorgan, D.L. The modulatory effects of non-thermal plasma on seed’s morphology, germination and genetics—A review. Plants 2022, 11, 2181. [Google Scholar] [CrossRef] [PubMed]
- Sirgedaitė-Šėžienė, V.; Mildažienė, V.; Žemaitis, P.; Ivankov, A.; Koga, K.; Shiratani, M.; Baliuckas, V. Long-term response of Norway spruce to seed treatment with cold plasma: Dependence of the effects on the genotype. Plasma Process. Polym. 2021, 18, 2000159. [Google Scholar] [CrossRef]
- Pawełek, A.; Wyszkowska, J.; Cecchetti, D.; Dinka, M.D.; Przybylski, K.; Szmidt-Jaworska, A. The physiological and biochemical response of field bean (Vicia faba L. (partim)) to electromagnetic field exposure is influenced by seed age, light conditions, and growth media. Agronomy 2022, 12, 2161. [Google Scholar] [CrossRef]
- Maffei, M.E. Magnetic field effects on plant growth, development, and evolution. Front. Plant Sci. 2014, 5, 445. [Google Scholar] [CrossRef]
- Pietruszewski, S.; Martínez, E. Magnetic field as a method of improving the quality of sowing material: A review. Int. Agrophys 2015, 29, 377–389. [Google Scholar] [CrossRef]
- Randeniya, L.K.; de Groot, G. Non-thermal plasma treatment of agricultural seeds for stimulation of germination, removal of surface contamination and other benefits: A review. Plasma Process. Polym. 2015, 12, 608–623. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A.; Dobránszki, J. Magnetic fields: How is plant growth and development impacted? Protoplasma 2016, 253, 231–248. [Google Scholar] [CrossRef]
- Degutytė-Fomins, L.; Paužaitė, G.; Žūkienė, R.; Mildažienė, V.; Koga, K.; Shiratani, M. Relationship between cold plasma treatment-induced changes in radish seed germination and phytohormone balance. Jpn. J. Appl. Phys. 2020, 59, SH1001. [Google Scholar] [CrossRef]
- Beniušytė, E.; Čėsnienė, I.; Sirgedaitė-Šėžienė, V.; Vaitiekūnaitė, D. Genotype-dependent jasmonic acid effect on Pinus sylvestris L. Growth and induced systemic resistance indicators. Plants 2023, 12, 255. [Google Scholar] [CrossRef]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef]
- Dvořák, P.; Krasylenko, Y.; Zeiner, A.; Šamaj, J.; Takáč, T. Signaling toward reactive oxygen species-scavenging enzymes in plants. Front. Plant Sci. 2021, 11, 618835. [Google Scholar] [CrossRef] [PubMed]
- Pospíšil, P. Production of reactive oxygen species by photosystem ii as a response to light and temperature stress. Front. Plant Sci. 2016, 7, 1950. [Google Scholar] [CrossRef] [PubMed]
- Del Río, L.A.; López-Huertas, E. ROS generation in peroxisomes and its role in cell signaling. Plant Cell Physiol. 2016, 57, 1364–1376. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Serrano, M.; L’Haridon, F.; Tjamos, S.E.; Metraux, J.P. Reactive oxygen species and plant resistance to fungal pathogens. Phytochemistry 2015, 112, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Eltayeb, A.E.; Kawano, N.; Badawi, G.H.; Kaminaka, H.; Sanekata, T.; Shibahara, T.; Inanaga, S.; Tanaka, K. Overexpression of monodehydroascorbate reductase in transgenic tobacco confers enhanced tolerance to ozone, salt and polyethylene glycol stresses. Planta 2007, 225, 1255–1264. [Google Scholar] [CrossRef] [PubMed]
- Arena, C.; Vitale, L.; Bianchi, A.R.; Mistretta, C.; Vitale, E.; Parisi, C.; Guerriero, G.; Magliulo, V.; De Maio, A. The ageing process affects the antioxidant defences and the poly (adpribosyl) ation activity in Cistus incanus L. Leaves. Antioxidants 2019, 8, 528. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; Mandzhieva, S. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Fimognari, L.; Dölker, R.; Kaselyte, G.; Jensen, C.; Akhtar, S.; Großkinsky, D.; Roitsch, T. Simple semi-high throughput determination of activity signatures of key antioxidant enzymes for physiological phenotyping. Plant Methods 2020, 16, 42. [Google Scholar] [CrossRef]
- Caverzan, A.; Passaia, G.; Barcellos Rosa, S.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef]
- Thor, M.; Stenlid, J. Heterobasidion annosum infection of Picea abies following manual or mechanized stump treatment. Scand. J. For. Res. 2005, 20, 154–164. [Google Scholar] [CrossRef]
- Oliva, J.; Bendz-Hellgren, M.; Stenlid, J. Spread of Heterobasidion annosum s.s. and Heterobasidion parviporum in Picea abies 15 years after stump inoculation. FEMS Microb. Ecol. 2011, 75, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Marčiulynas, A.; Sirgedaitė-Šėžienė, V.; Žemaitis, P.; Baliuckas, V. The resistance of scots pine (Pinus sylvestris L.) Half-sib families to Heterobasidion annosum. Forests 2019, 10, 287. [Google Scholar] [CrossRef]
- Striganavičiūtė, G.; Žiauka, J.; Sirgedaitė-Šėžienė, V.; Vaitiekūnaitė, D. Priming of resistance-related phenolics: A study of plant-associated bacteria and Hymenoscyphus fraxineus. Microorganisms 2021, 9, 2504. [Google Scholar] [CrossRef]
- Fossdal, C.G.; Krokene, P.; Olsen, J.E.; Strimbeck, R.; Viejo, M.; Yakovlev, I.; Mageroy, M.H. Epigenetic stress memory in gymnosperms. Plant Physiol. 2024, 195, 1117–1133. [Google Scholar] [CrossRef]
- Li, X.; Piao, S.; Wang, K.; Wang, X.; Wang, T.; Ciais, P.; Chen, A.; Lian, X.; Peng, S.; Peñuelas, J. Temporal trade-off between gymnosperm resistance and resilience increases forest sensitivity to extreme drought. Nat. Ecol. Evol. 2020, 4, 1075–1083. [Google Scholar] [CrossRef]
- Krokene, P. Chapter 5—Conifer Defense and Resistance to Bark Beetles. In Bark Beetles; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 177–207. [Google Scholar] [CrossRef]
- Anand, A.; Surendran, V.; Sukumaran, S. Bioactive metabolites in gymnosperms. In Plant Metabolites: Methods, Applications and Prospects; Springer: Singapore, 2020; pp. 17–346. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, S.; Li, L.; Yang, T.; Dong, S.; Wei, T.; Wu, S.; Liu, Y.; Gong, Y.; Feng, X.; et al. The Cycas genome and the early evolution of seed plants. Nat. Plants 2022, 8, 389–401. [Google Scholar] [CrossRef]
- Yan, Z.; Tian, F.; Wu, C.; Yan, S.; Kou, X.; Zhou, D.; Fan, G.; Li, T.; Bai, C. Transcriptome analysis of the related defense genes in postharvest Ginkgo biloba seeds against Penicillium oxalicum. Physiol. Mol. Plant Pathol. 2023, 127, 102060. [Google Scholar] [CrossRef]
- Ivankov, A.; Naučienė, Z.; Degutytė-Fomins, L.; Žūkienė, R.; Januškaitienė, I.; Malakauskienė, A.; Jakštas, V.; Ivanauskas, L.; Romanovskaja, D.; Šlepetienė, A.; et al. Changes in agricultural performance of common buckwheat induced by seed treatment with cold plasma and electromagnetic field. Appl. Sci. 2021, 11, 4391. [Google Scholar] [CrossRef]
- Pańka, D.; Jeske, M.; Łukanowski, A.; Baturo-Cieśniewska, A.; Prus, P.; Maitah, M.; Maitah, K.; Malec, K.; Rymarz, D.; Muhire, J.; et al. Can cold plasma be used for boosting plant growth and plant protection in sustainable plant production? Agronomy 2022, 12, 841. [Google Scholar] [CrossRef]
- Holubová, Ľ.; Kyzek, S.; Ďurovcová, I.; Fabová, J.; Horváthová, E.; Ševčovičová, A.; Gálová, E. Non-Thermal Plasma—A New Green Priming Agent for Plants? Int. J. Mol. Sci. 2020, 21, 9466. [Google Scholar] [CrossRef] [PubMed]
- Pauzaite, G.; Malakauskiene, A.; Nauciene, Z.; Zukiene, R.; Filatova, I.; Lyushkevich, V.; Azarko, I.; Mildaziene, V. Changes in Norway spruce germination and growth induced by pre-sowing seed treatment with cold plasma and electromagnetic field: Short-term versus long-term effects. Plasma Process. Polym. 2018, 15, 1700068. [Google Scholar] [CrossRef]
- Šerá, B.; Šerý, M.; Zahoranová, A.; Tomeková, J. Germination improvement of three pine species (Pinus) after diffuse coplanar surface barrier discharge plasma treatment. Plasma Chem. Plasma Process. 2021, 41, 211–226. [Google Scholar] [CrossRef]
- Meiqiang, Y.; Mingjing, H.; Buzhou, M.; Tengcai, M. Stimulating effects of seed treatment by magnetized plasma on tomato growth and yield. Plasma Sci. Technol. 2005, 7, 3143. [Google Scholar] [CrossRef]
- Zukiene, R.; Nauciene, Z.; Januskaitiene, I.; Pauzaite, G.; Mildaziene, V.; Koga, K.; Shiratani, M. Dielectric barrier discharge plasma treatment-induced changes in sunflower seed germination, phytohormone balance, and seedling growth. Appl. Phys. Express 2019, 12, 126003. [Google Scholar] [CrossRef]
- Švubová, R.; Kyzek, S.; Medvecká, V.; Slováková, Ľ.; Gálová, E.; Zahoranová, A. Novel insight at the effect of cold atmospheric pressure plasma on the activity of enzymes essential for the germination of pea (Pisum sativum L. cv. Prophet) seeds. Plasma Chem. Plasma Process. 2020, 40, 1221–1240. [Google Scholar] [CrossRef]
- Mildaziene, V.; Pauzaite, G.; Malakauskiene, A.; Zukiene, R.; Nauciene, Z.; Filatova, I.; Azharonok, V.; Lyushkevich, V. Response of perennial woody plants to seed treatment by electromagnetic field and low-temperature plasma. Bioelectromagnetics 2016, 37, 536–548. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxygen processing in photosynthesis: Regulation and signalling. New Phytol. 2020, 146, 359–388. [Google Scholar] [CrossRef]
- Žaltauskaitė, J.; Dikšaitytė, A.; Miškelytė, D.; Kacienė, G.; Sujetovienė, G.; Januškaitienė, I.; Juknys, R. Effects of elevated CO2 concentration and temperature on the mixed-culture grown wild mustard (Sinapis arvensis L.) response to auxin herbicide. Environ. Sci. Pollut. Res. 2023, 30, 13711–13725. [Google Scholar] [CrossRef]
- Tegischer, K.; Tausz, M.; Wieser, G.; Grill, D. Tree- and needle-age-dependent variations in antioxidants and photoprotective pigments in Norway spruce needles at the alpine timberline. Tree Physiol. 2002, 22, 591–596. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul. 2005, 46, 209–221. [Google Scholar] [CrossRef]
- Sharma, P.; Shanker Dubey, R. Modulation of nitrate reductase activity in rice seedlings under aluminium toxicity and water stress: Role of osmolytes as enzyme protectant. J. Plant Physiol. 2005, 162, 854–864. [Google Scholar] [CrossRef]
- Zlatev, Z.S.; Lidon, F.C.; Ramalho, J.C.; Yordanov, I.T. Comparison of resistance to drought of three bean cultivars. Biol. Plant 2006, 50, 389–394. [Google Scholar] [CrossRef]
- Gapińska, M.; Skłodowska, M.; Gabara, B. Effect of short- and long-term salinity on the activities of antioxidative enzymes and lipid peroxidation in tomato roots. Acta Physiol. Plant 2008, 30, 11–18. [Google Scholar] [CrossRef]
- Biareh, V.; Shekari, F.; Sayfzadeh, S.; Zakerin, H.; Hadidi, E.; Beltrão, J.G.T.; Mastinu, A. Physiological and qualitative response of Cucurbita pepo L. To salicylic acid under controlled water stress conditions. Horticulturae 2022, 8, 79. [Google Scholar] [CrossRef]
- Rouhi, H.; Aboutalebian, M.; Moosavi, S.; Karimi, F.; Karimi, F.; Saman, M.; Samadi, M. Change in several antioxidant enzymes activity of Berseem clover (Trifolium alexandrinum L.) by priming. Int. J. Agric. Sci. 2012, 2, 237–243. [Google Scholar]
- Guan, L.M.; Scandalios, J.G. Hydrogen peroxide-mediated catalase gene expression in response to wounding. Free Radic. Biol. Med. 2000, 28, 1182–1190. [Google Scholar] [CrossRef]
- Gould, N.; Reglinski, T.; Northcott, G.L.; Spiers, M.; Taylor, J.T. Physiological and biochemical responses in Pinus radiata seedlings associated with methyl jasmonate-induced resistance to Diplodia pinea. Physiol. Mol. Plant Pathol. 2009, 74, 121–128. [Google Scholar] [CrossRef]
- Çelik, Ö.; Büyükuslu, N.; Atak, C.; Rzakoulieva, A. Effects of Magnetic Field on Activity of Superoxide Dismutase and Catalase in Glycine max (L.) Merr. Roots. Pol. J. Environ. Stud. 2009, 18, 175–182. [Google Scholar]
- Ferreira-Santos, P.; Genisheva, Z.; Pereira, R.N.; Teixeira, J.A.; Rocha, C.M.R. Moderate electric fields as a potential tool for sustainable recovery of phenolic compounds from Pinus pinaster bark. ACS Sustain. Chem. Eng. 2019, 7, 8816–8826. [Google Scholar] [CrossRef]
- Bukhari, S.A.; Tanveer, M.; Mustafa, G.; Zia-Ud-Den, N. Magnetic field stimulation effect on germination and antioxidant activities of presown hybrid seeds of sunflower and its seedlings. J. Food Qual. 2021, 2021, 5594183. [Google Scholar] [CrossRef]
- Graziani, G.; Cirillo, A.; Giannini, P.; Conti, S.; El-Nakhel, C.; Rouphael, Y.; Ritieni, A.; Di Vaio, C. Biostimulants improve plant growth and bioactive compounds of young olive trees under abiotic stress conditions. Agriculture 2022, 12, 227. [Google Scholar] [CrossRef]
- Sarinont, T.; Amano, T.; Attri, P.; Koga, K.; Hayashi, N.; Shiratani, M. Effects of plasma irradiation using various feeding gases on growth of Raphanus sativus L. Arch. Biochem. Biophys. 2016, 605, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; von Tiedemann, A. Impact of fungicides on active oxygen species and antioxidant enzymes in spring barley (Hordeum vulgare L.) exposed to ozone. Environ. Pollut. 2002, 116, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Ragaee, S.; Abdel-Aal, E.-S.M.; Noaman, M. Antioxidant activity and nutrient composition of selected cereals for food use. Food Chem. 2006, 98, 32–38. [Google Scholar] [CrossRef]
- Lučinskaitė, I.; Laužikė, K.; Žiauka, J.; Baliuckas, V.; Čėsna, V.; Sirgedaitė-Šėžienė, V. Assessment of biologically active compounds, organic acids and antioxidant activity in needle extracts of different Norway spruce (Picea abies (L.) H. Karst) half-sib families. Wood Sci. Technol. 2021, 55, 1221–1235. [Google Scholar] [CrossRef]
- Student. The Probable Error of a Mean. Biometrika 1908, 6, 1–25. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Keselman, H.J.; Huberty, C.J.; Lix, L.M.; Olejnik, S.; Cribbie, R.A.; Donahue, B.; Levin, J.R. Statistical practices of educational researchers: An analysis of their ANOVA, MANOVA, and ANCOVA analyses. Rev. Educ. Res. 1998, 68, 350–386. [Google Scholar] [CrossRef]
- Kuhn, M.; Jackson, S.; Cimentada, J. Corrr: Correl. R. Available online: https://github.com/tidymodels/corrr (accessed on 2 March 2023).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
| Half-Sib Family | Emergence (%) ± SE | |||
|---|---|---|---|---|
| C | CP1 | CP2 | EMF2 | |
| 124 | 68.1 ± 0.4 | 65.0 ± 0.4 | 61.3 ± 0.4 | 68.8 ± 0.4 |
| 417 | 87.5 ± 0.3 | 79.4 ± 0.3 * | 85.0 ± 0.3 | 71.9 ± 0.3 *** |
| 454 | 79.4 ± 0.3 | 55.6 ± 0.4 *** | 60.0 ± 0.4 *** | 66.3 ± 0.4 ** |
| 457 | 43.1 ± 0.4 | 52.5 ± 0.4 | 48.1 ± 0.4 | 52.5 ± 0.4 |
| 463 | 66.7 ± 0.6 | 63.9 ± 0.6 | 75.0 ± 0.5 | 80.6 ± 0.3 * |
| 477 | 46.3 ± 0.4 | 52.5 ± 0.4 | 62.5 ± 0.4 ** | 55.0 ± 0.4 |
| 541 | 69.4 ± 0.4 | 63.8 ± 0.4 | 57.5 ± 0.4 * | 59.4 ± 0.4 |
| 548 | 61.3 ± 0.4 | 67.5 ± 0.4 | 61.9 ± 0.4 | 63.2 ± 0.4 |
| 577 | 65.6 ± 0.4 | 58.8 ± 0.4 | 54.4 ± 0.4 * | 60.0 ± 0.4 |
| 599 | 35.6 ± 0.4 | 35.6 ± 0.4 | 39.4 ± 0.4 | 31.3 ± 0.4 |
| Half-Sib Family | Height (cm) | |||||||
|---|---|---|---|---|---|---|---|---|
| One-Year-Old | Two-Year-Old | |||||||
| C | CP1 | CP2 | EMF2 | C | CP1 | CP2 | EMF2 | |
| 124 | 7.6 ± 0.2 | 7.6 ± 0.3 | 7.3 ± 0.2 | 7.0 ± 0.3 | 12.5 ± 0.6 | 13.1 ± 0.9 | 13.0 ± 0.5 | 11.9 ± 0.5 |
| 417 | 5.9 ± 0.2 | 6.9 ± 0.2 ** | 6.2 ± 0.2 | 5.1 ± 0.2 ** | 10.6 ± 0.5 | 13.3 ± 0.5 *** | 10.4 ± 0.9 | 9.9 ± 0.7 |
| 454 | 8.7 ± 0.3 | 7.5 ± 0.4 * | 7.3 ± 0.2 *** | 7.7 ± 0.2 * | 15.4 ± 0.7 | 14.0 ± 0.7 | 14.4 ± 0.5 | 13.7 ± 0.6 |
| 457 | 6.7 ± 0.3 | 7.9 ± 0.2 *** | 7.9 ± 0.2 *** | 7.6 ± 0.3 * | 15.0 ± 0.8 | 17.0 ± 0.9 * | 15.5 ± 0.6 | 15.2 ± 0.8 |
| 463 | 5.3 ± 0.3 | 8.3 ± 0.4 *** | 6.3 ± 0.3 * | 5.3 ± 0.2 | 17.1 ± 1.5 | 16.6 ± 1.2 | 13.5 ± 0.9 * | 15.2 ± 1.3 |
| 477 | 5.8 ± 0.3 | 6.9 ± 0.3 * | 6.2 ± 0.3 | 5.6 ± 0.3 | 14.6 ± 0.8 | 16.5 ± 0.9 | 13.7 ± 1.2 | 13.9 ± 0.9 |
| 541 | 8.8 ± 0.3 | 8.8 ± 0.3 | 8.1 ± 0.3 | 7.9 ± 0.3 * | 14.1 ± 0.4 | 15.7 ± 0.6 * | 13.8 ± 0.9 | 12.7 ± 0.6 * |
| 548 | 6.7 ± 0.2 | 7.9 ± 0.2 *** | 8.7 ± 0.2 *** | 6.8 ± 0.2 | 15.0 ± 0.8 | 15.8 ± 0.9 | 15.7 ± 0.5 | 14.0 ± 0.9 |
| 577 | 7.4 ± 0.3 | 6.3 ± 0.2 ** | 4.6 ± 0.2 *** | 5.1 ± 0.2 *** | 12.3 ± 0.6 | 12.7 ± 0.6 | 10.4 ± 0.8 * | 13.0 ± 0.7 |
| 599 | 7.8 ± 0.3 | 8.3 ± 0.3 | 8.6 ± 0.3 * | 8.5 ± 0.5 | 12.3 ± 0.9 | 10.1 ± 0.9 | 11.0 ± 0.7 | 12.0 ± 0.6 |
| Importance of Components (One-Year-Old) | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 |
|---|---|---|---|---|---|---|---|---|
| Standard deviation | 1.794 | 1.149 | 1.047 | 0.964 | 0.799 | 0.622 | 0.485 | 0.389 |
| Proportion of Variance | 0.403 | 0.165 | 0.137 | 0.117 | 0.080 | 0.048 | 0.0295 | 0.019 |
| Cumulative Proportion | 0.403 | 0.569 | 0.706 | 0.823 | 0.903 | 0.951 | 0.981 | 1.000 |
| Importance of Components (Two-Year-Old) | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 |
| Standard deviation | 1.565 | 1.318 | 1.084 | 0.989 | 0.921 | 0.632 | 0.573 | 0.246 |
| Proportion of Variance | 0.307 | 0.218 | 0.147 | 0.122 | 0.106 | 0.050 | 0.041 | 0.008 |
| Cumulative Proportion | 0.307 | 0.525 | 0.672 | 0.795 | 0.901 | 0.951 | 0.992 | 1.000 |
| Trait | One-Year-Old | Two-Year-Old | ||
|---|---|---|---|---|
| 1st Component | 2nd Component | 1st Component | 2nd Component | |
| CAT | 0.48201 | 0.02427 | 0.61008 | 0.01850 |
| APX | 0.51520 | −0.03179 | 0.45603 | 0.24341 |
| POX | 0.43443 | 0.18383 | 0.19429 | −0.33059 |
| GR | 0.50361 | −0.01278 | 0.60793 | 0.03309 |
| SOD | 0.19564 | −0.54432 | 0.03128 | −0.15240 |
| DPPH | 0.10827 | 0.65602 | −0.09389 | 0.59030 |
| ABTS | −0.00743 | 0.37482 | −0.02679 | 0.67699 |
| HEIGHT | 0.09022 | −0.31195 | −0.04462 | −0.00342 |
| Plantation Code | Regional Division of State Forestry | Forestry Sector | Geographical Position | Block | Site | Area (ha) | Year of Enrolment | |
|---|---|---|---|---|---|---|---|---|
| P. abies | 08ESP031 | Trakai | Būda | 54°54′38.7″ N 24°18′01.2″ E | 216 | 16, 17 | 5.32 | 2002 |
| Half-sib families: 124, 417, 454, 457, 463, 477, 541, 548, 577, 599 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čėsnienė, I.; Čėsna, V.; Miškelytė, D.; Novickij, V.; Mildažienė, V.; Sirgedaitė-Šėžienė, V. Seed Treatment with Cold Plasma and Electromagnetic Field: Changes in Antioxidant Capacity of Seedlings in Different Picea abies (L.) H. Karst Half-Sib Families. Plants 2024, 13, 2021. https://doi.org/10.3390/plants13152021
Čėsnienė I, Čėsna V, Miškelytė D, Novickij V, Mildažienė V, Sirgedaitė-Šėžienė V. Seed Treatment with Cold Plasma and Electromagnetic Field: Changes in Antioxidant Capacity of Seedlings in Different Picea abies (L.) H. Karst Half-Sib Families. Plants. 2024; 13(15):2021. https://doi.org/10.3390/plants13152021
Chicago/Turabian StyleČėsnienė, Ieva, Vytautas Čėsna, Diana Miškelytė, Vitalij Novickij, Vida Mildažienė, and Vaida Sirgedaitė-Šėžienė. 2024. "Seed Treatment with Cold Plasma and Electromagnetic Field: Changes in Antioxidant Capacity of Seedlings in Different Picea abies (L.) H. Karst Half-Sib Families" Plants 13, no. 15: 2021. https://doi.org/10.3390/plants13152021