
The Genetic Diversity of the Macrophyte Ceratophyllum demersum in Backwaters Reflects Differences in the Hydrological Connectivity and Water Flow Rate of Habitats

Abstract
1. Introduction
2. Materials and Methods
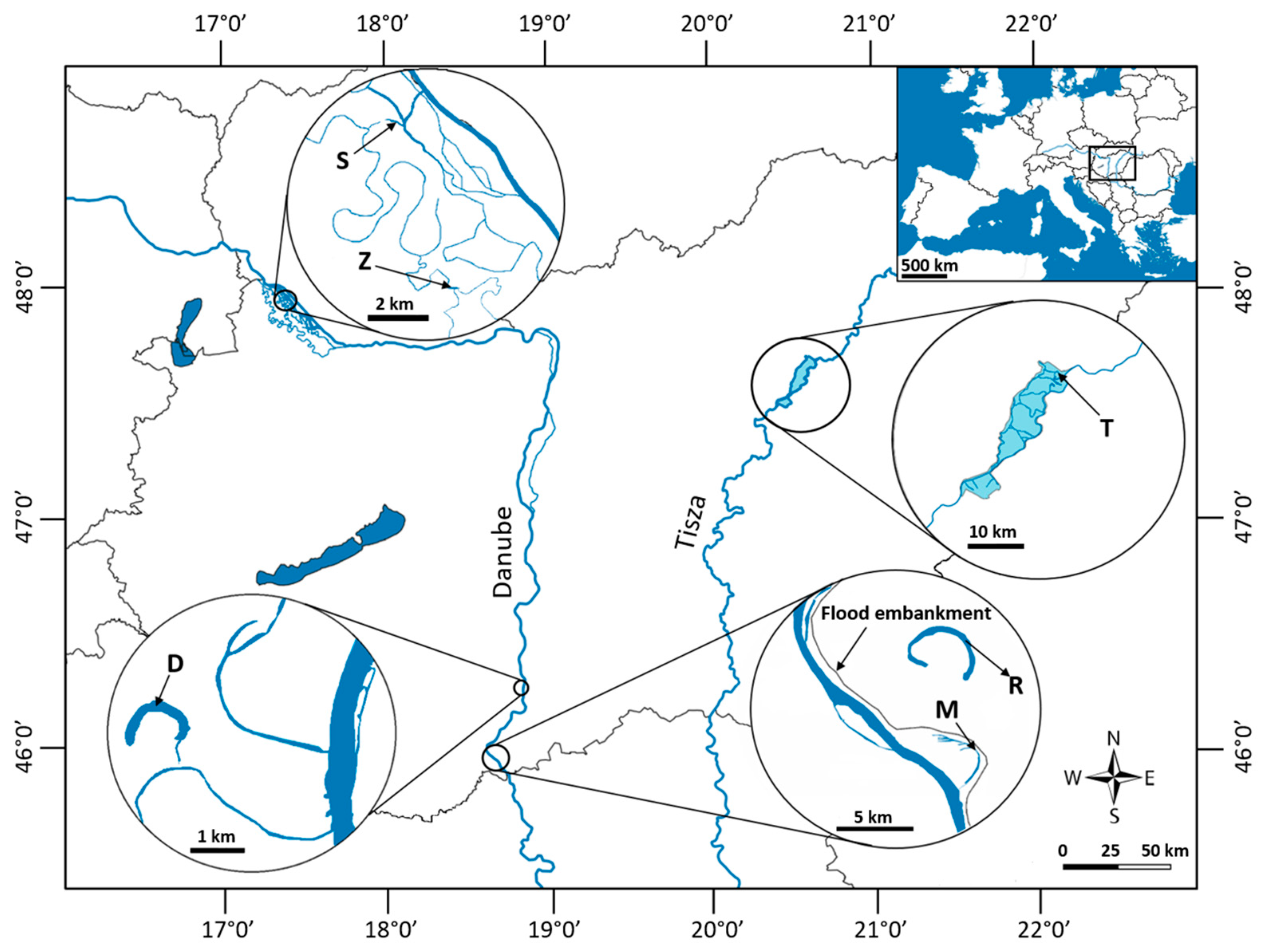
2.1. Study Sites and Sampling
2.2. Laboratory Work
2.3. Statistical Analyses
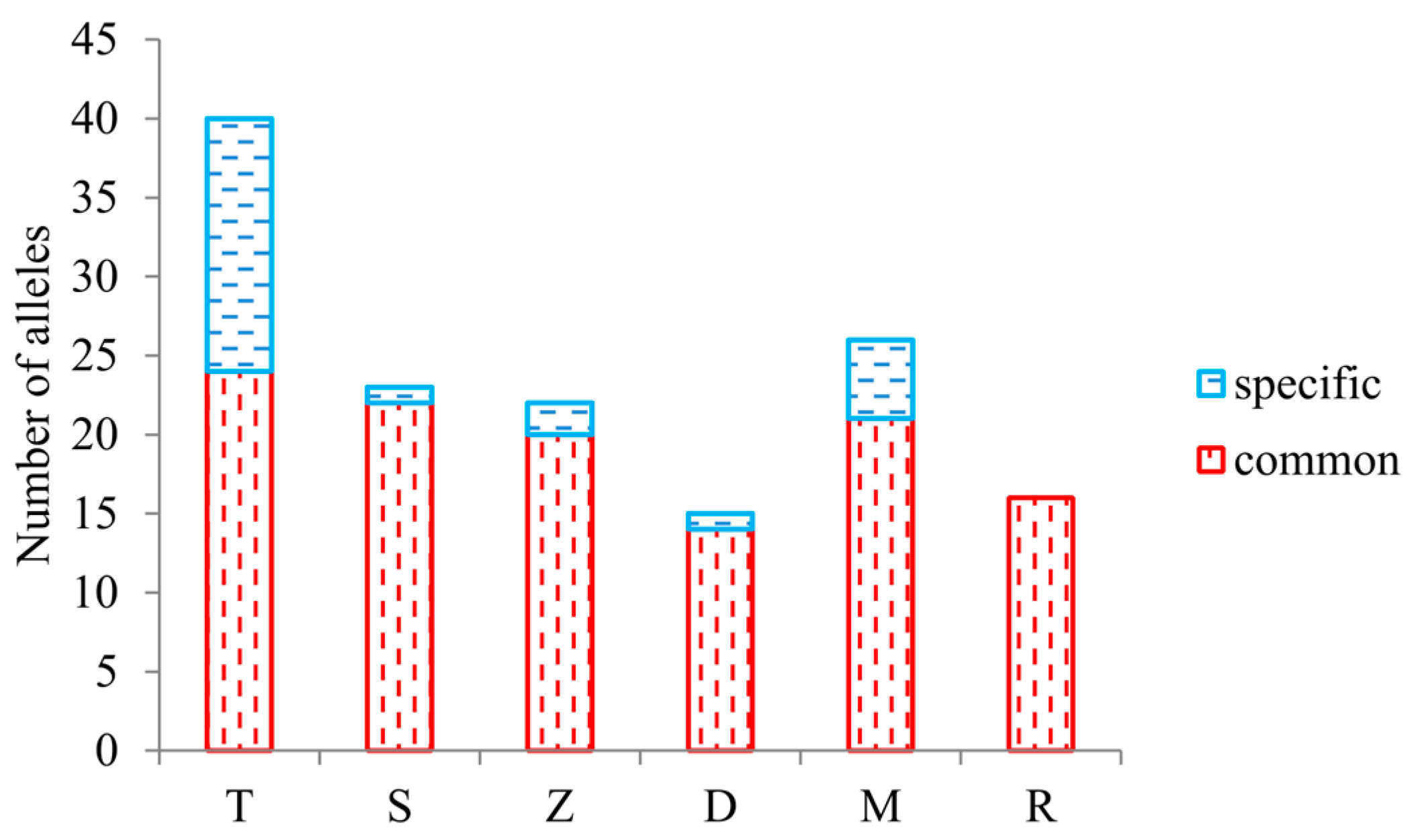
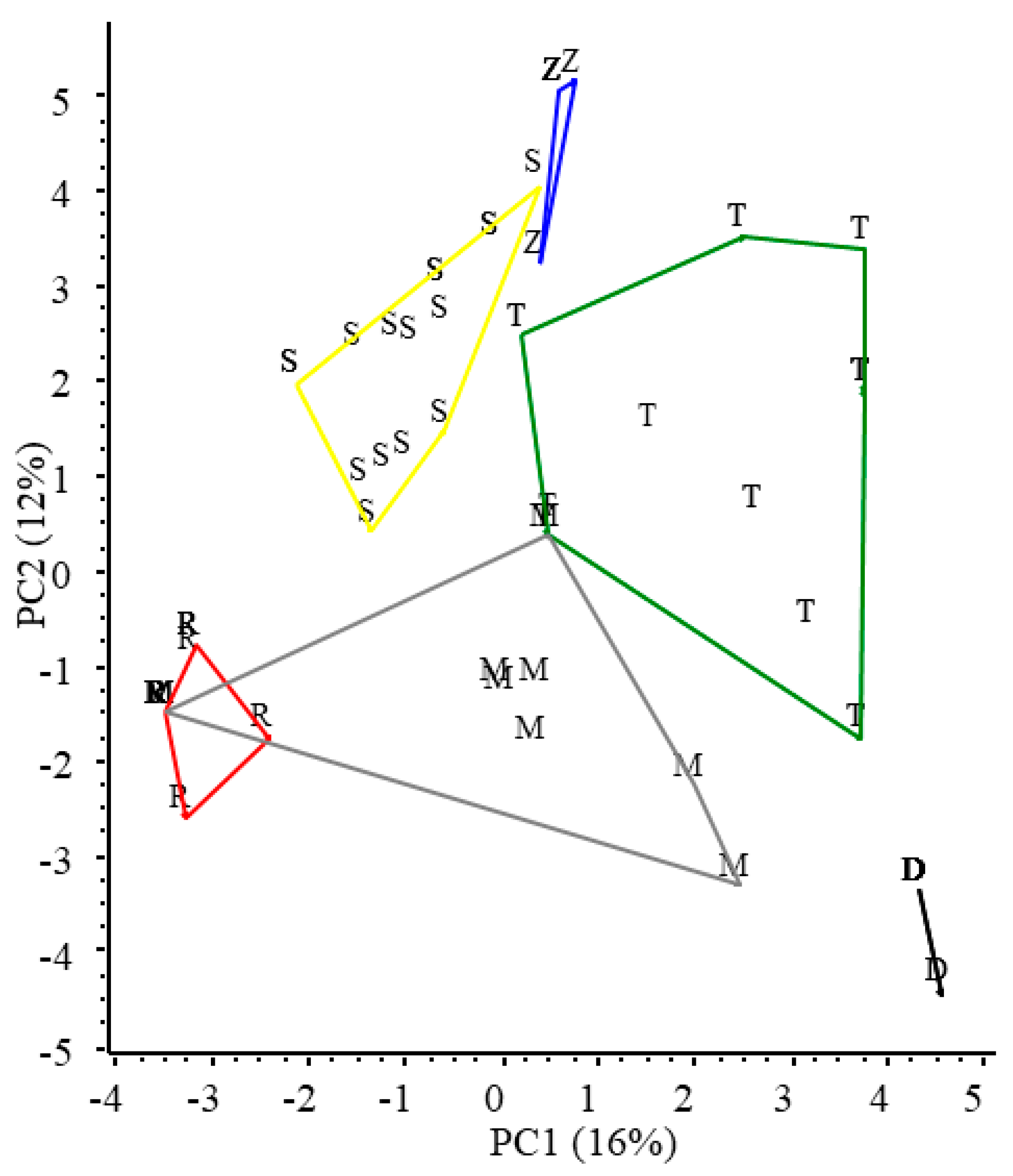
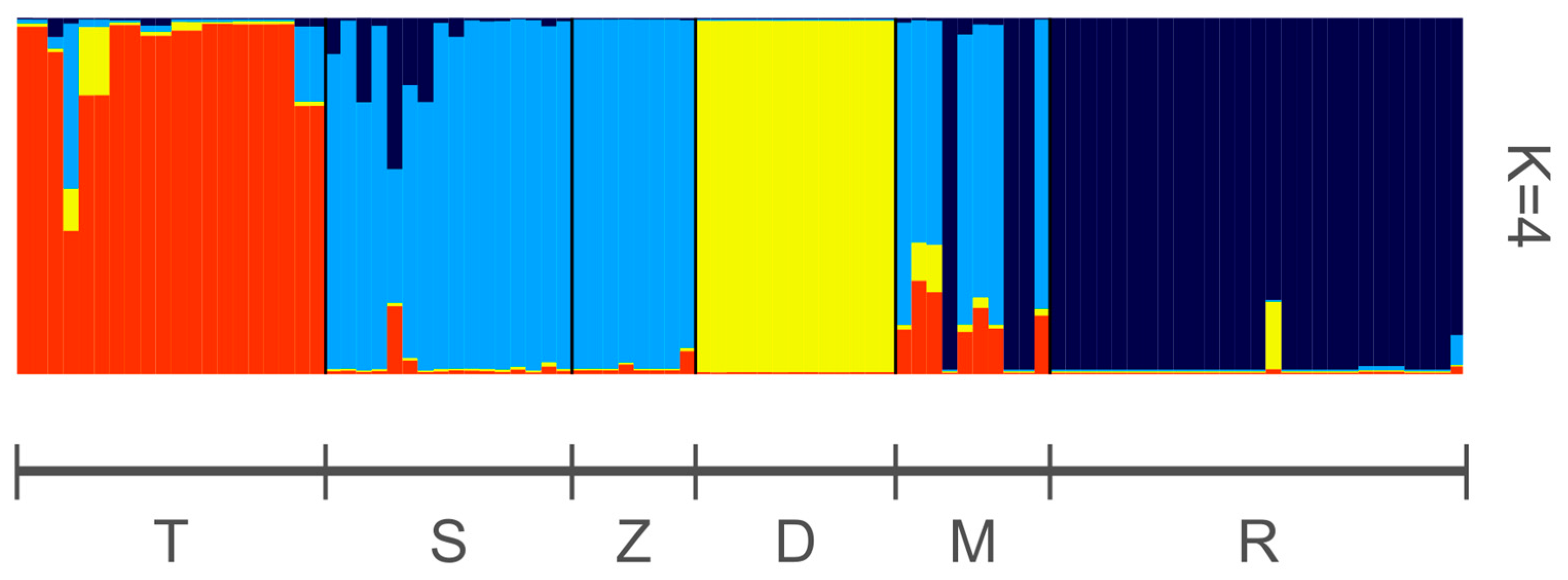
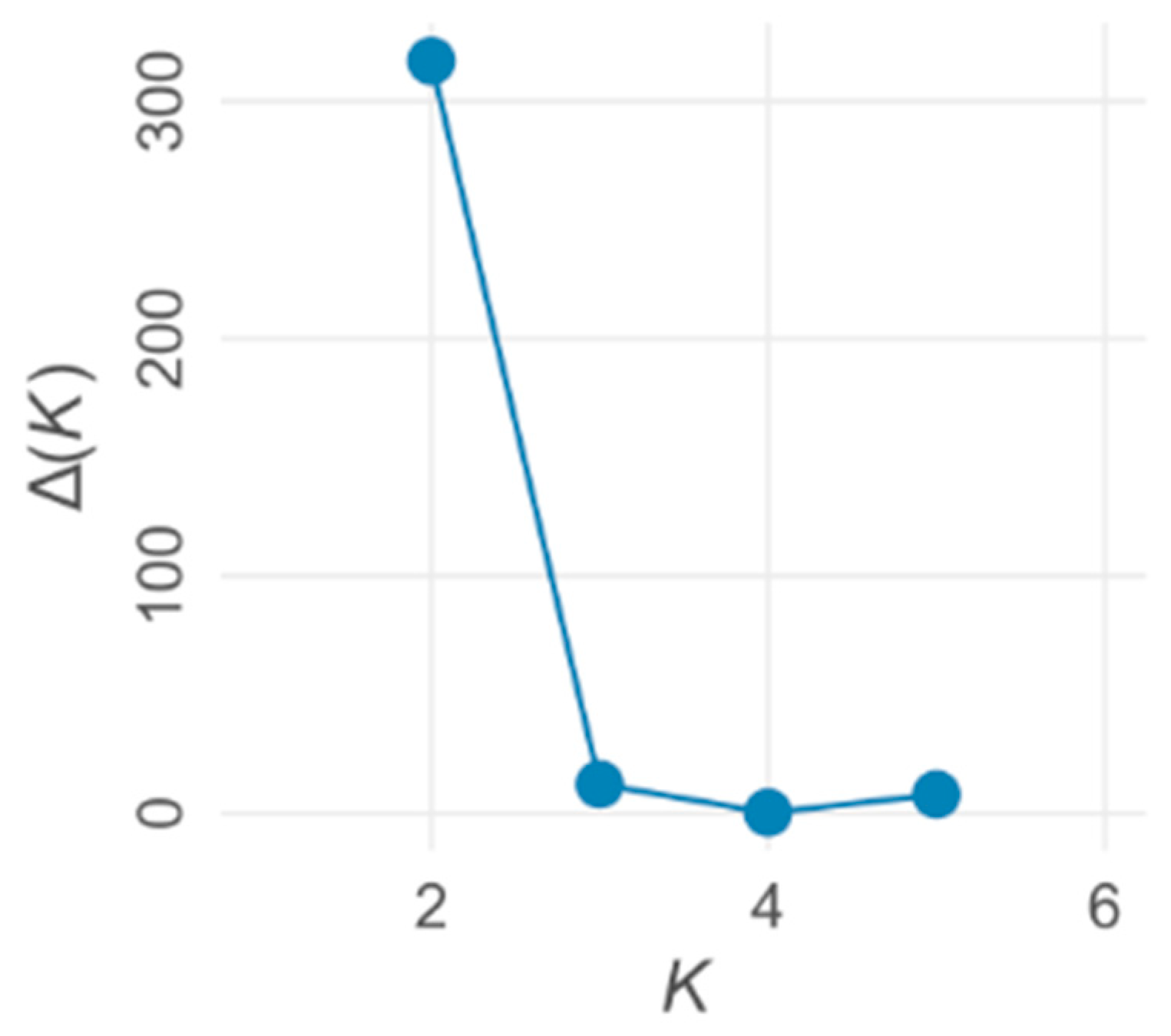
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bayley, P.B. Understanding large river-floodplain ecosystems. BioScience 1995, 45, 153–158. [Google Scholar] [CrossRef]
- Amoros, C.; Bornette, G. Connectivity and biocomplexity in waterbodies of riverine floodplains. Freshw. Biol. 2002, 47, 761–776. [Google Scholar] [CrossRef]
- Guan, Q.; Wu, H.; Xu, L.; Kang, Y.; Lu, K.; Liu, D.; Han, D.; Xue, Z.; Yuan, Y.; Wang, W.; et al. Hydrological connectivity shapes multiple diversity facets of snail (Mollusca: Gastropoda) assemblages in freshwater floodplain wetlands. Ecol. Indic. 2023, 153, 110467. [Google Scholar] [CrossRef]
- Brauer, C.J.; Beheregaray, L.B. Recent and rapid anthropogenic habitat fragmentation increases extinction risk for freshwater biodiversity. Evol. Appl. 2020, 13, 2857–2869. [Google Scholar] [CrossRef]
- Pavlova, A.; Beheregaray, L.B.; Coleman, R.; Gilligan, D.; Harrisson, K.A.; Ingram, B.A.; Kearns, J.; Lamb, A.M.; Lintermans, M.; Lyon, J.; et al. Severe consequences of habitat fragmentation on genetic diversity of an endangered Australian freshwater fish: A call for assisted gene flow. Evol. Appl. 2017, 10, 531–550. [Google Scholar] [CrossRef]
- Fan, X.R.; Njeri, H.K.; Li, W.; Chen, Y.Y. Abundant historical gene flow within and among river systems for populations of Ottelia acuminata var. jingxiensis, an endangered macrophyte from southwest China. Aquat. Bot. 2019, 157, 1–9. [Google Scholar] [CrossRef]
- Wu, Z.; Xu, X.; Zhang, J.; Wiegleb, G.; Hou, H. Influence of environmental factors on the genetic variation of the aquatic macrophyte Ranunculus subrigidus on the Qinghai-Tibetan Plateau. BMC Evol. Biol. 2019, 19, 228. [Google Scholar] [CrossRef]
- Reitsema, R.E.; Wolters, J.-W.; Preiner, S.; Meire, P.; Hein, T.; De Boeck, G.; Blust, R.; Schoelynck, J. Response of submerged macrophyte growth, morphology, chlorophyll content and nutrient stoichiometry to increased flow velocity and elevated CO2 and dissolved organic carbon concentrations. Front. Environ. Sci. 2020, 11, 527801. [Google Scholar] [CrossRef]
- Goldenberg-Vilar, A.; Delgado, C.; Penas, F.J.; Barquín, J. The effect of altered flow regimes on aquatic primary producer communities: Diatoms and macrophytes. Ecohydrology 2022, 15, e2353. [Google Scholar] [CrossRef]
- Santamaría, L. Why are most aquatic plants widely distributed? Dispersal, clonal growth and small-scale heterogeneity in a stressful environment. Acta Oecol. 2002, 23, 137–154. [Google Scholar] [CrossRef]
- Cao, Q.; Hu, F.; Liu, N. Local-scale patterns of genetic variation in coexisting floating-leaved Nymphoides peltata and submerged Myriophyllum spicatum in Donghu Lake. J. Oceanol. Limnol. 2020, 38, 1825–1834. [Google Scholar] [CrossRef]
- Cao, Q.; Liu, B.; Hu, F. Effects of hydrological connection and human disturbance on genetic variation of submerged Vallisneria natans populations in four lakes in China. J. Oceanol. Limnol. 2021, 39, 1403–1416. [Google Scholar] [CrossRef]
- Chen, Y.; Li, X.; Yin, L.; Cheng, Y.; Li, W. Genetic diversity and migration patterns of the aquatic macrophyte Potamogeton malaianus in a potamolacustrine system. Freshw. Biol. 2009, 54, 1178–1188. [Google Scholar] [CrossRef]
- Triest, L.; Tran Thi, V.; Le Thi, D.; Sierens, T.; Van Geert, A. Genetic differentiation of submerged plant populations and taxa between habitats. Hydrobiologia 2010, 656, 15–27. [Google Scholar] [CrossRef]
- Jones, P.E.; Consuegra, S.; Börger, L.; Jones, J.; Garcia de Leaniz, C. Impacts of artificial barriers on the connectivity and dispersal of vascular macrophytes in rivers: A critical review. Freshw. Biol. 2020, 65, 1165–1180. [Google Scholar] [CrossRef]
- Pollux, B.J.A.; Luteijn, A.; Van Groenendael, J.M.; Ouborg, N.J. Gene flow and genetic structure of the aquatic macrophyte Sparganium emersum in a linear unidirectional river. Freshw. Biol. 2009, 54, 64–76. [Google Scholar] [CrossRef]
- Arber, A. Water Plants; Cambridge University Press: London, UK, 1920. [Google Scholar]
- Les, D.H. The phytogeography of Ceratophyllum demersum and C. echinatum (Ceratophyllaceae) in glaciated North America. Can. J. Bot. 1986, 64, 498–509. [Google Scholar] [CrossRef]
- Les, D.H. Genetic diversity in the monoecious hydrophile Ceratophyllum (Ceratophyllaceae). Am. J. Bot. 1991, 78, 1070–1082. [Google Scholar] [CrossRef]
- Wade, P.M. The influence of vegetation pre-dredging on the post-dredging community. J. Aquat. Plant Manag. 1993, 31, 141–144. [Google Scholar]
- Fukuhara, H.; Tanaka, T.; Izumi, M. Growth and turion formation of Ceratophyllum demersum in a shallow lake in Japan. Jpn. J. Limnol. 1997, 58, 335–347. [Google Scholar] [CrossRef]
- Capers, R.S. Macrophyte colonization in a freshwater tidal wetland (Lyme, CT, USA). Aquat. Bot. 2003, 77, 325–338. [Google Scholar] [CrossRef]
- Engloner, A.I.; Németh, K.; Kós, P.B.; Meglécz, E.; Bereczki, J. Genetic diversity of the submerged macrophyte Ceratophyllum demersum depends on habitat hydrology and habitat fragmentation. Front. Plant Sci. 2023, 14, 1277916. [Google Scholar] [CrossRef] [PubMed]
- O’Briain, R. Climate change and European rivers: An eco-hydromorphological perspective. Ecohydrology 2019, 12, e2099. [Google Scholar] [CrossRef]
- Rivaes, R.P.; Feio, M.J.; Almeida, S.F.P.; Calapez, A.R.; Sales, M.; Gebler, D.; Lozanovska, I.; Aguiar, F.C. River ecosystem endangerment from climate change-driven regulated flow regimes. Sci. Total Environ. 2022, 818, 151857. [Google Scholar] [CrossRef] [PubMed]
- Hungarian Hydrological Forecasting Service. Available online: https://www.hydroinfo.hu/en/index.html (accessed on 2 July 2024).
- Bousquet, J.; Simon, L.; Lalonde, M. DNA amplification from vegetative and sexual tissues of trees using polymerase chain reaction. Can. J. For. Res. 1990, 20, 254–257. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research–an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Goudet, J. Fstat (ver. 2.9.4), a Program to Estimate and Test Population Genetics Parameters. Available online: http://www2.unil.ch/popgen/softwares/fstat.htm (accessed on 11 August 2023).
- Leberg, P.L. Estimating allelic richness: Effects of sample size and bottlenecks. Mol. Ecol. 2002, 11, 2445–2449. [Google Scholar] [CrossRef]
- Weir, B.S. Genetic Data Analysis. Methods for Discrete Population Genetic Data; Sinauer Associates Inc.: Sunderland, MA, USA, 1990; pp. 124–125. [Google Scholar]
- Podani, J. SYN-TAX Computer Programs for Data Analysis in Ecology and Systematics; User’s Manual; Scientia: Budapest, Hungary, 2001. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Francis, R.M. POPHELPER: An R package and web app to analyse and visualize population structure. Mol. Ecol. Resour. 2017, 17, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Wu, Z.; Wang, H.; Chen, J.; Xu, X. Population genetic structure of the whorl-leaf watermilfoil Myriophyllum verticillatum shaped by topography and geographic distance. Hydrobiologia 2019, 838, 55–64. [Google Scholar] [CrossRef]
- de Witte, L.C.; Stöcklin, J. Longevity of clonal plants: Why it matters and how to measure it. Ann. Bot. 2010, 106, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Eckert, G.C.; Dorken, M.E.; Barrett, S.C.H. Ecological and evolutionary consequences of sexual and clonal reproduction in aquatic plants. Aquat. Bot. 2016, 135, 46–61. [Google Scholar] [CrossRef]
- Zhu, J.; Xu, X.; Tao, Q.; Yi, P.; Yu, D.; Xu, X. High invasion potential of Hydrilla verticillata in the Americas predicted using ecological niche modeling combined with genetic data. Ecol. Evol. 2017, 7, 4982–4990. [Google Scholar] [CrossRef] [PubMed]
- Gornall, R.J.; Hollingsworth, P.M.; Preston, C.D. Evidence for spatial structure and directional gene flow in a population of an aquatic plant, Potamogeton coloratus. Heredity 1998, 80, 414–421. [Google Scholar] [CrossRef]
- Coppi, A.; Lastrucci, L.; Carta, A.; Foggi, B. Analysis of genetic structure of Ranunculus baudotii in a Mediterranean wetland. Implications for selection of seeds and seedlings for conservation. Aquat. Bot. 2015, 126, 25–31. [Google Scholar] [CrossRef]
- Stoffers, T.; Altermatt, F.; Baldan, D.; Bilous, O.; Borgwardt, F.; Buijse, A.D.; Bondar-Kunze, E.; Cid, N.; Erős, T.; Ferreira, M.T.; et al. Reviving Europe’s rivers: Seven challenges in the implementation of the Nature Restoration Law to restore free-flowing rivers. WIREs Water 2024, 11, e1717. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Connection to the Riverbed | Flow of Water | |
|---|---|---|
| Tisza-Reservoir (T) | permanent, close | permanent |
| Schisler (S) | permanent, close | temporary |
| Zátonyi (Z) | permanent, distant | absent |
| Decsi (D) | temporary, rare | temporary |
| Mocskos (M) | temporary, moderately rare | temporary |
| Riha (R) | absent | absent |
| N | Na | Ne | AR | I | MG | %P | |
|---|---|---|---|---|---|---|---|
| Tisza-Reservoir (T) | 19.778 | 4.444 | 2.605 | 3.726 | 1.023 | 10 | 100.00 |
| Schisler (S) | 15.778 | 2.556 | 2.311 | 2.529 | 0.781 | 13 | 77.78 |
| Zátonyi (Z) | 8.000 | 2.444 | 1.873 | 2.444 | 0.630 | 3 | 77.78 |
| Decsi (D) | 13.000 | 1.667 | 1.665 | 1.667 | 0.462 | 2 | 66.67 |
| Mocskos (M) | 10.000 | 2.889 | 2.125 | 2.817 | 0.804 | 8 | 100.00 |
| Riha (R) | 27.000 | 1.778 | 1.359 | 1.505 | 0.285 | 5 | 66.67 |
| Total | 15.593 | 2.630 | 1.990 | 3.611 | 0.664 | 40 | 81.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Engloner, A.I.; Németh, K.; Bereczki, J. The Genetic Diversity of the Macrophyte Ceratophyllum demersum in Backwaters Reflects Differences in the Hydrological Connectivity and Water Flow Rate of Habitats. Plants 2024, 13, 2220. https://doi.org/10.3390/plants13162220
Engloner AI, Németh K, Bereczki J. The Genetic Diversity of the Macrophyte Ceratophyllum demersum in Backwaters Reflects Differences in the Hydrological Connectivity and Water Flow Rate of Habitats. Plants. 2024; 13(16):2220. https://doi.org/10.3390/plants13162220
Chicago/Turabian StyleEngloner, Attila I., Kitti Németh, and Judit Bereczki. 2024. "The Genetic Diversity of the Macrophyte Ceratophyllum demersum in Backwaters Reflects Differences in the Hydrological Connectivity and Water Flow Rate of Habitats" Plants 13, no. 16: 2220. https://doi.org/10.3390/plants13162220
APA StyleEngloner, A. I., Németh, K., & Bereczki, J. (2024). The Genetic Diversity of the Macrophyte Ceratophyllum demersum in Backwaters Reflects Differences in the Hydrological Connectivity and Water Flow Rate of Habitats. Plants, 13(16), 2220. https://doi.org/10.3390/plants13162220