
Fine-Scale Lithogeochemical Features Influence Plant Distribution Patterns in Alpine Grasslands in the Western Alps of Italy

Abstract
:1. Introduction
2. Results
2.1. Bedrock Typification Based on Geological Maps and Stereomicrosopic Observations
2.2. Bedrock Typification Based on Geochemistry
- -
- FS did not contain CaCO3 and had low contents of Ni and MgO as well as of other mafic indicators such as Co and Cr. However, FS was quite rich in Fe2O3, another mafic indicator. FS showed the highest contents of SiO2, Al2O3, Na2O and P2O5, Ba, Ce, Ga, Hf, La, Nd, Th, and Zr (Table 1). The rather high Na2O content was presumably linked to Na-rich plagioclase feldspar, a discriminating mineral in intermediate rocks. From a petrological point of view, the FS group was heterogeneous and included both meta-gabbros and non-calcareous mica-schists.
- -
- MO had an intermediate CaCO3 content but had discrepancies in the concentrations of mafic indicators, with intermediate contents of MgO, Fe2O3, Co, and Cr but high concentrations of Ni and Cu. Furthermore, MO presented high concentrations of MnO, K2O, Ba, Ga, Nb, Nd, Pb, Rb, Th, and Zn (Table 1). Petrologically, MO also was heterogeneous and included chlorite schists together with calcareous and non-calcareous mica-schists.
- -
- OC was a hybrid lithological group with intermediate characteristics between ophiolites s.l. and calc-schists s.l. This was due to the medium-high concentrations of CaCO3, CaO, MgO, Ni, and Sr. OC also had a high Pb content and was La-free (Table 1).
- -
- CS had the highest contents of CaCO3, CaO, and Sr and lower concentrations of Fe2O3, MgO, Cr, Ni, Sc, and Zn than almost all other lithological groups (Table 1).
- -
- TS had extremely high contents of mafic indicators such as MgO, Co, Cr, and Ni. Furthermore, TS did not contain CaCO3, K2O, Ga, Hf, La, and Nd (Table 1).
- -
- SS had a particular geochemical composition. Similar to TS, this group was rich in MgO, Ni, Co, Cr, and Sc and had high concentrations of TiO2, Fe2O3, MnO, Na2O, P2O5, V, and Y as well (Table 1).
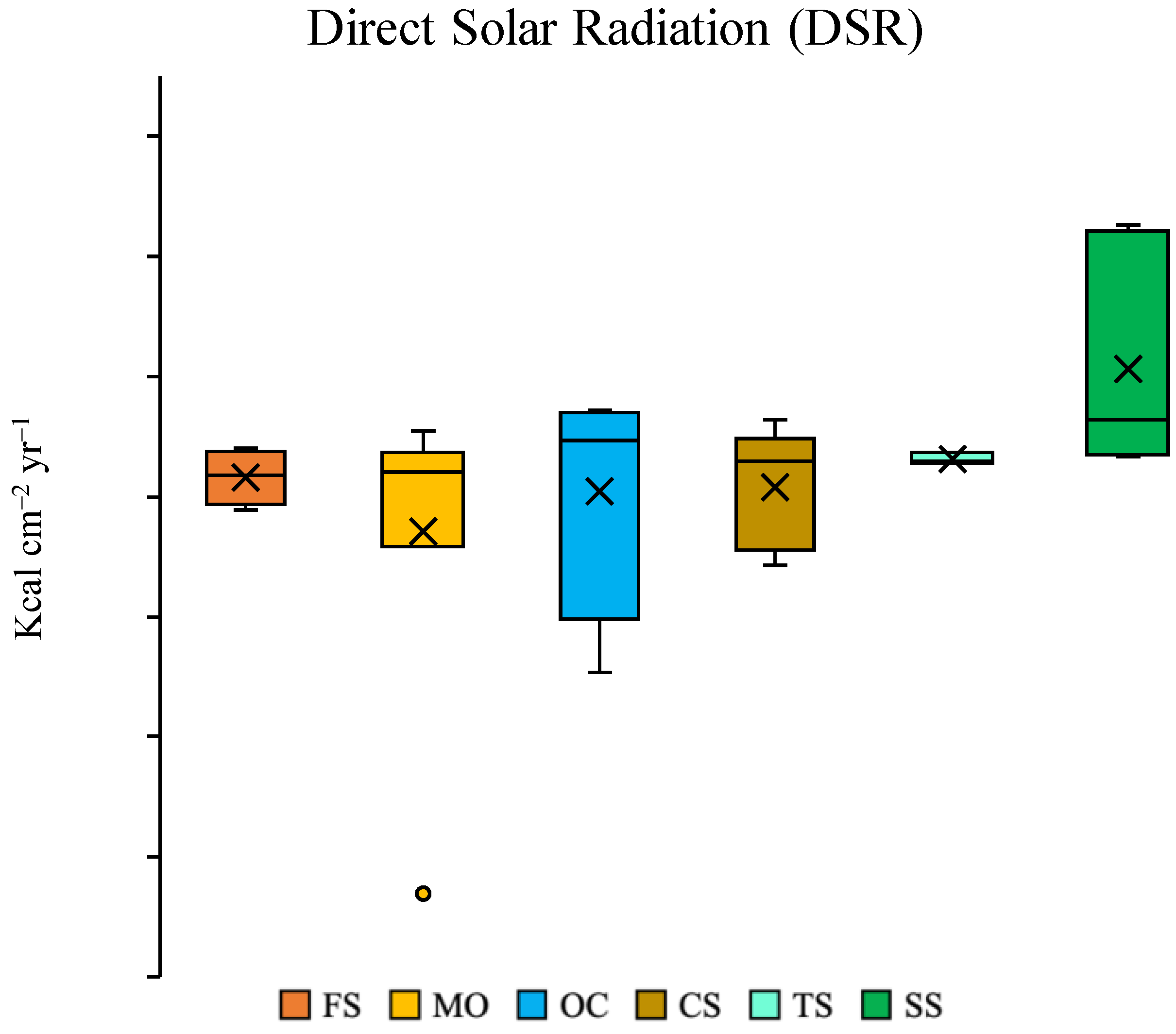
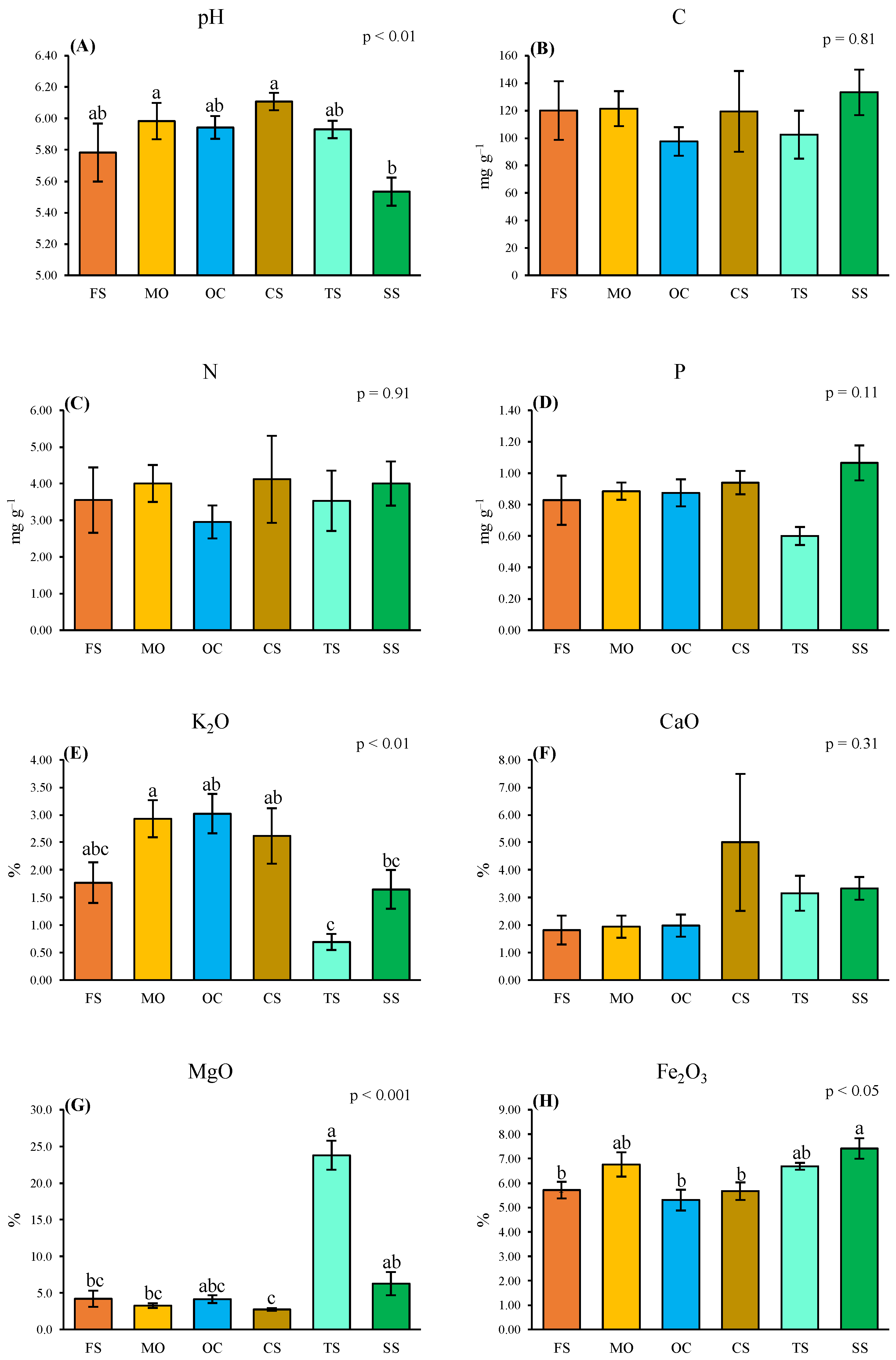
2.3. Direct Solar Radiation (DSR) and Soil Chemistry
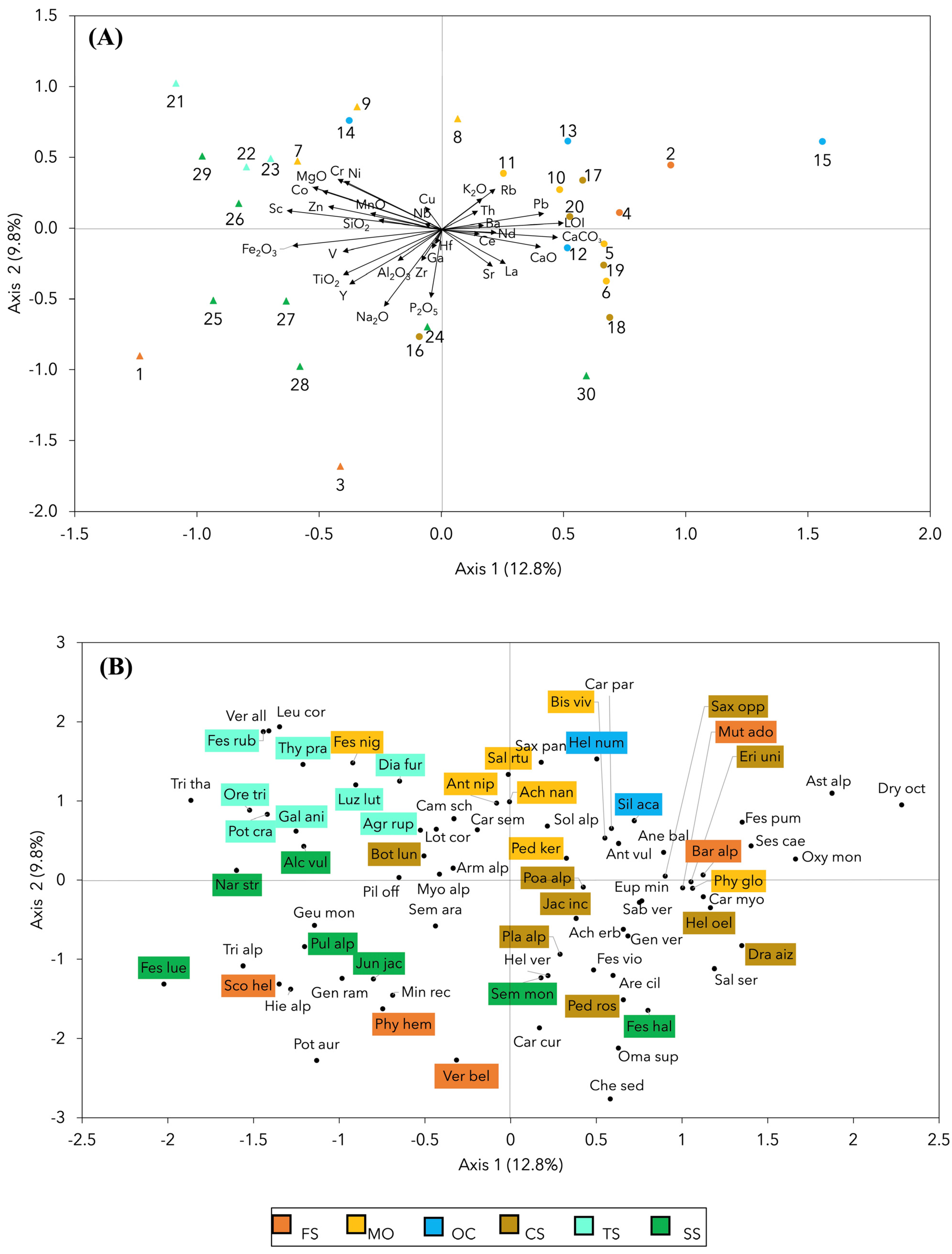
2.4. Vegetation and Its Relationships with the Lithological Groups and Bedrock Chemistry
- -
- FS indicator species: Bartsia alpina, Mutellina adonidifolia, Phyteuma hemisphaericum, Scorzoneroides helvetica, and Veronica bellidioides.
- -
- MO indicator species: Achillea nana, Pedicularis kerneri, Salix retusa, Anthoxanthum nipponicum, Festuca nigricans, and Bistorta vivipara.
- -
- OC indicator species: Helianthemum nummularium subsp. grandiflorum, and Silene acaulis.
- -
- CS indicator species: Helianthemum oelandicum subsp. alpestre, Botrychium lunaria, Draba aizoides subsp. aizoides, Erigeron uniflorus, Pedicularis rosea subsp. allionii, Phyteuma globulariifolium subsp. pedemontanum, Plantago alpina, Poa alpina, Saxifraga oppositifolia subsp. oppositifolia, and Jacobaea incana.
- -
- TS indicator species: Luzula lutea subsp. lutea, Potentilla crantzii subsp. crantzii, Thymus praecox subsp. polytrichus, Agrostis rupestris subsp. rupestris, Dianthus furcatus, Festuca rubra, Galium anisophyllon, and Oreojuncus trifidus.
- -
- SS indicator species: Juncus jacquinii, Alchemilla vulgaris, Festuca halleri, Festuca scabriculmis subsp. luedii, Nardus stricta, Pulsatilla alpina subsp. apiifolia, and Sempervivum montanum.
3. Discussion
4. Materials and Methods
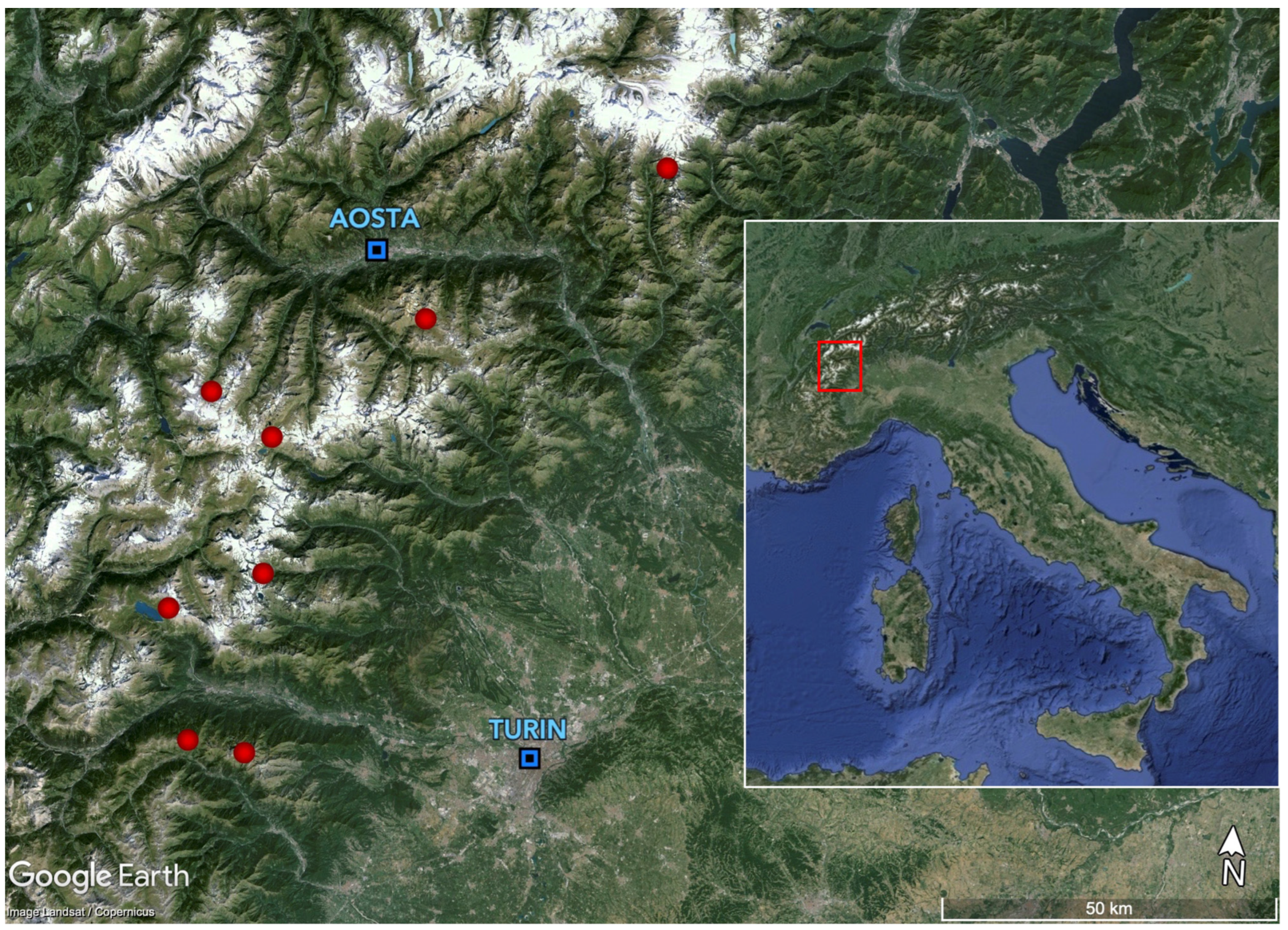
4.1. Study Area, Sampling Design, and Data Collection
4.2. Bedrock and Soil Analyses
4.3. Data Compilation and Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Poulenard, J.; Podwojewski, P. Alpine soils. In Encyclopedia of Soil Science; Chesworth, W., Ed.; Marcel Dekker: New York, NY, USA, 2003; pp. 1–4. [Google Scholar]
- Väre, H.; Lampinen, R.; Humphries, C.; Williams, P. Taxonomic diversity of vascular plants in the European alpine areas. In Alpine Biodiversity in Europe; Körner, C., Grabherr, G., Nagy, L., Thompson, D.B.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 133–148. [Google Scholar]
- Ellenberg, H. The vegetation above the alpine tree line. In Vegetation Ecology of Central Europe, 4th ed.; Cambridge University Press: Melbourne, Australia, 1988; pp. 388–455. [Google Scholar]
- Leuschner, C.; Ellenberg, H. Vegetation of the alpine and nival belts. In Ecology of Central European Nonforest Vegetation: Coastal to Alpine, Natural to Man-made Habitats: Vegetation Ecology of Central Europe; Springer Nature: Cham, Switzerland, 2017; Volume 2, pp. 271–431. [Google Scholar]
- Körner, C. Concepts in alpine plant ecology. Plants 2023, 12, 2666. [Google Scholar] [CrossRef]
- Braun-Blanquet, J.; Jenny, H. Vegetationsentwicklung und Bodenbildung in der alpinen Stufe der Zentralalpen (Klimaxgebiet des Caricion curvulae). Mèmoires Société Helvétique Sci. Nat. 1926, 63, 183–349. [Google Scholar]
- Gigon, A. Vergleich alpiner Rasen auf Silikat- und auf Karbonatboden: Konkurrenz- und Stickstofformen Versuche sowie standörtliche Untersuchungen im Nardetum und im Seslerietum bei Davos. Veröffentlichungen Geobot. Inst. Eidg. Techn. Hochsch. Stift. Rübel 1971, 48, 1–159. [Google Scholar]
- Reisigl, H.; Keller, R. Alpenpflanzen im Lebensraum (Alpine Rasen, Schuttund Felsvegetation); Gustav Fischer Verlag: Stuttgart, Germany, 1987; 149p. [Google Scholar]
- Tyler, G. Some ecophysiological and historical approaches to species richness and calcicole/calcifuge behaviour—Contribution to a debate. Folia Geobot. 2003, 38, 419–428. [Google Scholar] [CrossRef]
- Lee, J.A. The calcicole-calcifuge problem revisited. Adv. Bot. Res. 1999, 29, 2–30. [Google Scholar]
- Zohlen, A.; Tyler, G. Immobilization of tissue iron on calcareous soil: Differences between calcicole and calcifuge plants. Oikos 2000, 89, 95–106. [Google Scholar] [CrossRef]
- Zohlen, A.; Tyler, G. Soluble inorganic tissue phosphorus and calcicole–calcifuge behaviour of plants. Ann. Bot.-London 2004, 94, 427–432. [Google Scholar] [CrossRef]
- Lambers, H.; Hayes, P.E.; Laliberté, E.; Oliveira, R.S.; Turner, B.L. Leaf manganese accumulation and phosphorus-acquisition efficiency. Trends Plant Sci. 2015, 20, 83–90. [Google Scholar] [CrossRef] [PubMed]
- de Souza, M.C.; Habermann, G.; do Amaral, C.L.; Rosa, A.L.; Ongaro Pinheiro, M.H.; Da Costa, F.B. Vochysia tucanorum Mart.: An aluminum-accumulating species evidencing calcifuge behavior. Plant Soil 2017, 419, 377–389. [Google Scholar] [CrossRef]
- Hayes, P.E.; Guilherme Pereira, C.; Clode, P.L.; Lambers, H. Calcium-enhanced phosphorus toxicity in calcifuge and soil-indifferent Proteaceae along the Jurien Bay chronosequence. New Phytol. 2019, 221, 764–777. [Google Scholar] [CrossRef]
- Wala, M.; Kolodziejek, J.; Mazur, J. The diversity of iron acquisition strategies of calcifuge plant species from dry acidic grasslands. J. Plant Physiol. 2023, 280, 153898. [Google Scholar] [CrossRef] [PubMed]
- Kovács, G.; Radovics, B.G.; Tóth, T.M. Petrologic comparison of the Gyód and Helesfa serpentinite bodies (Tisia Mega Unit, SW Hungary). J. Geosci.-Czech 2016, 61, 255–279. [Google Scholar] [CrossRef]
- Wakabayashi, J. Serpentinites and serpentinites: Variety of origins and emplacement mechanisms of serpentinite bodies in the California Cordillera. Island Arc. 2017, 26, e12205. [Google Scholar] [CrossRef]
- Burianek, D.; Pertoldova, J. Garnet-forming reactions in calc-silicate rocks from the Policka Unit, Svratka Unit and SE part of the Moldanubian Zone. J. Geosci.-Czech 2009, 54, 245–268. [Google Scholar] [CrossRef]
- Nabelek, P.I.; Morgan, S.S. Metamorphism and fluid flow in the contact aureole of the Eureka Valley–Joshua Flat–Beer Creek pluton, California. Geol. Soc. Am. Bull. 2012, 124, 228–239. [Google Scholar] [CrossRef]
- Pfiffner, A. Geology of the Alps; Wiley-Blackwell: Hoboken, NJ, USA, 2014; 376p. [Google Scholar]
- Lagabrielle, Y.; Cannat, M. Alpine Jurassic ophiolites resemble the modern central Atlantic basement. Geology 1990, 18, 319–322. [Google Scholar] [CrossRef]
- Fioraso, G.; Balestro, G.; Festa, A.; Lanteri, L. Role of structural inheritance in the gravitational deformation of the Monviso meta-ophiolite complex: The Pui-Orgiera serpentinite landslide (Varaita Valley, Western Alps). J. Maps 2019, 15, 372–381. [Google Scholar] [CrossRef]
- Geological Map of Italy, Scale 1:100,000, Sheets n. 28, 29, 41, 55; Geological Survey of Italy. Available online: https://sgi.isprambiente.it/geologia100k/nord.aspx (accessed on 30 April 2024).
- Ague, J.J. Element mobility during regional metamorphism in crustal and subduction zone environments with a focus on the rare earth elements (REE). Am. Mineral. 2017, 102, 1796–1821. [Google Scholar] [CrossRef]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; Gavilán García, R.; et al. Vegetation of Europe: Hierarchical floristic classification systemof vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Vercesi, G.V. Valutazioni ecologiche sulla flora e sulla vegetazione delle ofioliti della Valmalenco (SO). Rev. Valdôtaine Hist. Nat. 2004, 58, 121–128. [Google Scholar]
- Mainetti, A.; Ravetto Enri, S.; Lonati, M. Vegetation trajectories in proglacial primary successions within Gran Paradiso National Park: A comparison between siliceous and basic substrates. Ibex-J. Mt. Ecol. 2022, 14, 1–18. [Google Scholar]
- Verger, J.P. Premières considerations sur la végétation alpine et les sols developpés sur serpentinites, prasinites et gabbros dans les Alpes Graies (Italie). Webbia 1993, 47, 313–328. [Google Scholar] [CrossRef]
- Pignatti, E.; Pignatti, S. Plant Life of the Dolomites. Vegetation Structure and Ecology; Springer: Berlin/Heidelberg, Germany, 2014; p. 769. [Google Scholar]
- Dal Piaz, G.V.; Bistacchi, A.; Massironi, M. Geological outline of the Alps. Episodes J. Int. Geosci. 2003, 26, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Principi, G.; Bortolotti, V.; Chiari, M.; Cortesogno, L.; Gaggero, L.; Marcucci, M.; Saccani, E.; Treves, B. The pre-orogenic volcano-sedimentary covers of the western Tethys oceanic basin: A revue. Ofioliti 2004, 29, 177–211. [Google Scholar]
- Tartarotti, P.; Martin, S.; Festa, A.; Balestro, G. Metasediments covering ophiolites in the HP internal belt of the Western Alps: Review of tectono-stratigraphic successions and constraints for the Alpine evolution. Minerals 2021, 11, 411. [Google Scholar] [CrossRef]
- Prinetti, F. Brevi note per i botanici sulle rocce ofiolitiche. Rev. Valdôtaine Hist. Nat. 2004, 58, 5–6. [Google Scholar]
- Proctor, J.; Woodell, S.R. The plant ecology of serpentine: I. Serpentine vegetation of England and Scotland. J. Ecol. 1971, 59, 375–395. [Google Scholar] [CrossRef]
- Baumbach, H. Metallophytes and metallicolous vegetation: Evolutionary aspects, taxonomic changes and conservational status in Central Europe. In Perspectives on Nature Conservation—Patterns, Pressures and Prospects; Tiefenbacher, J., Smiljanic, T., Eds.; InTech: Rijeka, Croatia, 2012; pp. 93–118. [Google Scholar]
- D’Amico, M.E.; Previtali, F. Small-scale variability of soil properties and soil–vegetation relationships in patterned ground on different lithologies (NW Italian Alps). Catena 2012, 135, 73–95. [Google Scholar] [CrossRef]
- Robinson, B.H.; Brooks, R.R.; Kirkman, J.H.; Gregg, E.H.; Gremigni, P. Plant-available elements in soils and their influence on the vegetation over ultramafic (‘serpentine’) rocks in New Zealand. J. Roy. Soc. New Zeal. 1996, 26, 457–468. [Google Scholar] [CrossRef]
- Cheng, C.H.; Jien, S.H.; Iizuka, Y.; Tsai, H.; Chang, Z.Y.; Hseu, Z.Y. Pedogenic chromium and nickel partitioning in serpentine soils along a toposequence. Soil Sci. Soc. Am. J. 2011, 75, 659–668. [Google Scholar] [CrossRef]
- Alexander, E.B. Arid to humid serpentine soils, mineralogy, and vegetation across the Klamath Mountains, USA. Catena 2014, 116, 114–122. [Google Scholar] [CrossRef]
- Kierczak, J.; Pędziwiatr, A.; Waroszewski, J.; Modelska, M. Mobility of Ni, Cr and Co in serpentine soils derived on various ultrabasic bedrocks under temperate climate. Geoderma 2016, 268, 78–91. [Google Scholar] [CrossRef]
- Yang, C.Y.; Nguyen, D.Q.; Ngo, H.T.T.; Navarrete, I.A.; Nakao, A.; Huang, S.T.; Hseu, Z.Y. Increases in Ca/Mg ratios caused the increases in the mobile fractions of Cr and Ni in serpentinite-derived soils in humid Asia. Catena 2022, 216, 106418. [Google Scholar] [CrossRef]
- Baker, A.J.; Brooks, R. Terrestrial higher plants which hyperaccumulate metallic elements. A review of their distribution, ecology and phytochemistry. Biorecovery 1989, 1, 81–126. [Google Scholar]
- Deng, T.H.B.; van der Ent, A.; Tang, Y.T.; Sterckeman, T.; Echevarria, G.; Morel, J.L.; Qiu, R.L. Nickel hyperaccumulation mechanisms: A review on the current state of knowledge. Plant Soil 2018, 423, 1–11. [Google Scholar] [CrossRef]
- Mahey, S.; Kumar, R.; Sharma, M.; Kumar, V.; Bhardwaj, R. A critical review on toxicity of cobalt and its bioremediation strategies. SN Appl. Sci. 2020, 2, 1279. [Google Scholar] [CrossRef]
- Roychoudhury, A. Vanadium uptake and toxicity in plants. SF J. Agri. Crop Manag. 2020, 1, 1010. [Google Scholar]
- Srivastava, D.; Tiwari, M.; Dutta, P.; Singh, P.; Chawda, K.; Kumari, M.; Chakrabarty, D. Chromium stress in plants: Toxicity, tolerance and phytoremediation. Sustainability 2021, 13, 4629. [Google Scholar] [CrossRef]
- Brady, K.U.; Kruckeberg, A.R.; Bradshaw, H.D. Evolutionary ecology of plant adaptation to ultramafic soils. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 243–266. [Google Scholar] [CrossRef]
- Wallossek, C. The acidophilous taxa of the Festuca varia group in the Alps: New studies on taxonomy and phytosociology. Folia Geobot. 1999, 34, 47–75. [Google Scholar] [CrossRef]
- Lüth, C.; Tasser, E.; Niedrist, G.; Dalla Via, J.; Tappeiner, U. Plant communities of mountain grasslands in a broad cross-section of the Eastern Alps. Flora 2011, 206, 433–443. [Google Scholar] [CrossRef]
- Gensac, P. Plant and soil groups in the alpine grasslands of the Vanoise massif, French Alps. Arctic Alpine Res. 1990, 22, 195–201. [Google Scholar] [CrossRef]
- Béguin, C.; Progin Sonney, M.; Vonlanthen, M. La végétation des sols polygonaux aux étages alpin supérieur et subnival en Valais (Alpes centro-occidentales, Suisse). Bot. Helv. 2006, 116, 41–54. [Google Scholar] [CrossRef]
- 33.4.1 All. Drabion Hoppeanae Zollitsch ex Merxm. & Zollitsch 1967. Available online: https://www.prodromo-vegetazione-italia.org/scheda/drabion-hoppeanae-zollitsch-ex-merxm--zollitsch-1967/469 (accessed on 30 April 2024).
- Aubert, S.; Boucher, F.; Lavergne, S.; Renaud, J.; Choler, P. 1914–2014: A revised worldwide catalogue of cushion plants 100 years after Hauri and Schröter. Alpine Bot. 2014, 124, 59–70. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer: Cham, Switzerland, 2021; pp. 23–51. Available online: https://link.springer.com/book/10.1007/978-3-030-59538-8 (accessed on 30 April 2024).
- Arnesen, G.; Beck, P.S.A.; Engelskjøn, T. Soil acidity, content of carbonates, and available phosphorus are the soil factors best correlated with alpine vegetation: Evidence from Troms, North Norway. Arct-Antarct. Alp. Res. 2007, 39, 189–199. [Google Scholar] [CrossRef]
- D’Amico, M.E.; Gorra, R.; Freppaz, M. Relationships between serpentine soils and vegetation in a xeric inner-Alpine environment. Plant Soil 2015, 376, 111–128. [Google Scholar] [CrossRef]
- Poldini, L.; Martini, F. La vegetazione delle vallette nivali su calcare, dei conoidi e delle alluvioni nel Friuli (NE Italia). Stud. Geobot. 1993, 13, 141–214. [Google Scholar]
- Winkler, D.E.; Butz, R.; Germino, M.J.; Reinhardt, K.; Kueppers, L.M. Snowmelt timing regulates community composition, phenology, and physiological performance of alpine plants. Front. Plant Sci. 2018, 9, 1140. [Google Scholar] [CrossRef]
- do Carmo, F.F.; de Campos, I.C.; Jacobi, C.M. Effects of fine-scale surface heterogeneity on rockoutcrop plant community structure. J. Veg. Sci. 2016, 27, 50–59. [Google Scholar] [CrossRef]
- Portal to the Flora of Italy. Available online: https://dryades.units.it/floritaly/index.php (accessed on 30 April 2024).
- Lachance, G.R.; Traill, R.J. Practical solution to the matrix problem in X-ray analysis. Can. Spectrosc. 1966, 11, 43–48. [Google Scholar]
- Buffo, J.M.; Fritschen, L.J.; Murphy, J.L. Direct Solar Radiation on Various Slopes from 0 to 60 Degrees North Latitude; Pacific Northwest Forest and Range Experiment Station, Forest Service, US Department of Agriculture: Portland, OR, USA, 1972; Volume 142.
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Paleontol. Electron. 2001, 4, 1–9. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geochemical Variables | Lithological Group | ANOVAS’ Summary | ||||||
|---|---|---|---|---|---|---|---|---|
| FS (n = 4) | MO (n = 7) | OC (n = 4) | CS (n = 5) | TS (n = 3) | SS (n = 7) | F | p | |
| SiO2 (%) | 59.87 ± 1.48 a | 54.96 ± 4.68 a | 35.08 ± 2.46 bc | 13.37 ± 1.37 c | 43.16 ± 2.06 abc | 47.58 ± 1.63 ab | 27.82 | <0.001 |
| TiO2 (%) | 0.79 ± 0.08 a | 0.61 ± 0.03 ab | 0.27 ± 0.06 bc | 0.13 ± 0.02 c | 0.11 ± 0.05 c | 1.18 ± 0.16 a | 18.73 | <0.001 |
| Al2O3 (%) | 18.71 ± 1.23 a | 15.49 ± 1.30 ab | 6.67 ± 1.04 bc | 2.93 ± 0.53 c | 2.07 ± 0.11 c | 14.51 ± 0.48 ab | 45.15 | <0.001 |
| Fe2O3 (%) | 6.25 ± 0.45 ab | 5.10 ± 0.36 bc | 2.63 ± 0.41 cd | 1.97 ± 0.30 d | 5.78 ± 1.17 abc | 9.98 ± 1.13 a | 15.61 | <0.001 |
| MnO (%) | 0.08 ± 0.01 b | 0.20 ± 0.04 a | 0.08 ± 0.02 b | 0.07 ± 0.01 b | 0.09 ± 0.01 b | 0.17 ± 0.02 a | 5.53 | <0.001 |
| MgO (%) | 3.38 ± 0.37 bcd | 2.23 ± 0.23 cd | 4.50 ± 0.76 abc | 1.14 ± 0.13 d | 33.32 ± 2.94 a | 8.60 ± 0.55 ab | 158.30 | <0.001 |
| CaO (%) | 1.84 ± 0.80 c | 8.22 ± 2.80 c | 22.85 ± 1.58 ab | 36.54 ± 1.56 a | 6.82 ± 3.68 bc | 9.93 ± 1.38 bc | 32.83 | <0.001 |
| Na2O (%) | 3.43 ± 0.23 a | 1.10 ± 0.14 ab | 0.67 ± 0.14 bc | 0.39 ± 0.03 bc | 0.14 ± 0.04 c | 3.32 ± 0.29 a | 47.85 | <0.001 |
| K2O (%) | 2.44 ± 0.41 a | 2.81 ± 0.31 a | 1.44 ± 0.14 ab | 0.36 ± 0.15 bc | 0.00 ± 0.00 c | 0.29 ± 0.13 bc | 24.07 | <0.001 |
| P2O5% (%) | 0.16 ± 0.01 a | 0.08 ± 0.02 bc | 0.08 ± 0.01 abcd | 0.07 ± 0.01 cd | 0.01 ± 0.00 d | 0.15 ± 0.04 a | 4.67 | <0.001 |
| L.O.I. (%) | 3.06 ± 0.32 d | 9.19 ± 2.15 bc | 25.73 ± 0.75 ab | 43.03 ± 3.16 a | 8.52 ± 1.50 bcd | 4.29 ± 1.12 cd | 62.75 | <0.001 |
| Ba (mg kg−1) | 834.25 ± 155.27 a | 421.01 ± 51.50 a | 279.15 ± 27.08 ab | 82.94 ± 13.27 bc | 18.43 ± 6.28 c | 43.61 ± 13.59 c | 24.59 | <0.001 |
| Ce (mg kg−1) | 55.25 ± 11.85 a | 29.64 ± 3.10 ab | 9.83 ± 4.52 bc | 4.74 ± 1.99 c | 1.60 ± 0.83 c | 3.31 ± 1.51 c | 19.00 | <0.001 |
| Co (mg kg−1) | 5.80 ± 2.04 d | 17.87 ± 7.27 bc | 13.93 ± 1.91 cd | 9.02 ± 1.81 cd | 96.43 ± 7.29 a | 37.73 ± 5.91 ab | 44.35 | <0.001 |
| Cr (mg kg−1) | 59.53 ± 21.09 cd | 136.76 ± 19.51 bc | 167.25 ± 36.38 abc | 22.48 ± 2.42 d | 2718.10 ± 646.77 a | 251.74 ± 35.34 ab | 33.52 | <0.001 |
| Cu (mg kg−1) | 17.95 ± 6.79 b | 48.30 ± 9.87 a | 20.40 ± 2.38 ab | 17.44 ± 4.16 b | 17.70 ± 8.16 ab | 30.14 ± 7.33 ab | 2.75 | <0.001 |
| Ga (mg kg−1) | 19.25 ± 1.78 a | 16.99 ± 2.02 a | 6.73 ± 1.87 bc | 2.88 ± 0.22 c | 0.00 ± 0.00 c | 11.44 ± 1.17 ab | 19.86 | <0.001 |
| Hf (mg kg−1) | 5.23 ± 0.23 a | 3.47 ± 0.54 ab | 1.20 ± 0.17 bc | 0.24 ± 0.07 c | 0.00 ± 0.00 c | 1.97 ± 0.38 b | 21.22 | <0.001 |
| La (mg kg−1) | 11.63 ± 5.22 a | 7.93 ± 3.69 ab | 0.00 ± 0.00 b | 3.54 ± 3.54 ab | 0.00 ± 0.00 b | 3.99 ± 1.17 ab | 1.70 | 0.17 |
| Nb (mg kg−1) | 14.40 ± 1.05 ab | 15.34 ± 1.56 a | 3.38 ± 0.82 c | 0.86 ± 0.50 c | 0.30 ± 0.12 c | 4.64 ± 0.56 bc | 37.98 | <0.001 |
| Nd (mg kg−1) | 33.13 ± 9.06 a | 21.27 ± 2.21 a | 6.68 ± 1.34 b | 6.46 ± 1.10 b | 0.00 ± 0.00 b | 7.51 ± 1.07 b | 11.94 | <0.001 |
| Ni (mg kg−1) | 23.75 ± 7.68 bc | 91.53 ± 17.56 a | 81.23 ± 9.64 ab | 20.96 ± 1.64 c | 1653.33 ± 150.76 a | 84.83 ± 5.86 a | 214.90 | <0.001 |
| Pb (mg kg−1) | 7.43 ± 1.61 abc | 14.61 ± 1.96 a | 11.43 ± 2.13 a | 10.64 ± 2.84 ab | 3.83 ± 0.86 bc | 3.54 ± 0.55 c | 6.27 | <0.001 |
| Rb (mg kg−1) | 61.20 ± 11.56 ab | 104.01 ± 12.06 a | 47.25 ± 6.13 ab | 19.38 ± 4.28 bc | 1.47 ± 0.09 c | 5.36 ± 1.90 c | 25.05 | <0.001 |
| Sc (mg kg−1) | 10.70 ± 1.39 ab | 10.37 ± 2.66 ab | 1.43 ± 0.56 bc | 0.10 ± 0.03 c | 16.73 ± 5.59 ab | 18.39 ± 1.62 a | 10.92 | <0.001 |
| Sr (mg kg−1) | 129.75 ± 48.68 bc | 207.73 ± 39.00 b | 223.93 ± 19.10 ab | 629.04 ± 97.89 a | 9.23 ± 5.56 c | 201.47 ± 19.82 b | 15.45 | <0.001 |
| Th (mg kg−1) | 7.85 ± 1.04 a | 7.00 ± 0.70 a | 2.38 ± 0.27 ab | 1.14 ± 0.33 bc | 0.17 ± 0.09 c | 0.40 ± 0.11 bc | 37.50 | <0.001 |
| V (mg kg−1) | 135.93 ± 35.66 ab | 115.76 ± 12.84 ab | 60.23 ± 11.88 bc | 29.52 ± 2.46 c | 66.57 ± 4.89 bc | 188.43 ± 11.56 a | 15.14 | <0.001 |
| Y (mg kg−1) | 29.03 ± 5.22 a | 23.56 ± 2.19 ab | 12.08 ± 2.01 bc | 10.58 ± 2.72 bc | 1.83 ± 0.49 c | 32.37 ± 3.46 a | 12.37 | <0.001 |
| Zn (mg kg−1) | 73.18 ± 7.25 ab | 91.83 ± 7.59 a | 41.10 ± 5.58 bc | 19.02 ± 2.03 c | 70.03 ± 11.11 ab | 72.57 ± 7.61 ab | 13.36 | <0.001 |
| Zr (mg kg−1) | 230.58 ± 5.79 a | 173.84 ± 17.34 ab | 68.00 ± 5.03 bc | 18.66 ± 2.31 c | 1.43 ± 1.24 c | 128.27 ± 24.08 ab | 21.94 | <0.001 |
| CaCO3 (%) | 0.00 ± 0.00 c | 9.14 ± 4.11 bc | 34.00 ± 2.83 ab | 71.20 ± 1.80 a | 0.00 ± 0.00 c | 1.86 ± 1.86 c | 93.63 | <0.001 |
| FS | MO | OC | CS | TS | SS | ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plot | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | |
| Indicator species | |||||||||||||||||||||||||||||||
| FS | |||||||||||||||||||||||||||||||
| SC—Bartsia alpina (*) | Bar alp | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||
| Ot.—Mutellina adonidifolia (*) | Mut ado | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||
| CC—Phyteuma hemisphaericum (*) | Phy hem | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||
| CC – Scorzoneroides helvetica (*) | Sco hel | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||
| CC—Veronica bellidioides (*) | Ver bel | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||
| MO | |||||||||||||||||||||||||||||||
| DH—Achillea nana (**) | Ach nan | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| CC—Pedicularis kerneri (**) | Ped ker | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||
| AC—Salix retusa (**) | Sal rtu | 1 | 2 | 4 | 1 | 1 | |||||||||||||||||||||||||
| Ot.—Anthoxanthum nipponicum (*) | Ant nip | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| SC—Festuca nigricans (*) | Fes nig | 2 | 2 | 3 | 2 | 2 | 1 | 1 | 2 | 2 | |||||||||||||||||||||
| Ot.—Bistorta vivipara (*) | Bis viv | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||
| OC | |||||||||||||||||||||||||||||||
| SC—Helianthemum nummularium subsp. grandiflorum (**) | Hel num | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| CC—Silene acaulis (*) | Sil aca | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||
| CS | |||||||||||||||||||||||||||||||
| SC—Helianthemum oelandicum subsp. alpestre (**) | Hel oel | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | ||||||||||||||||||||
| CC—Botrychium lunaria (*) | Bot lun | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||
| SC—Draba aizoides subsp. aizoides (*) | Dra aiz | 1 | 1 | 1 | |||||||||||||||||||||||||||
| OE—Erigeron uniflorus (*) | Eri uni | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||
| SC—Pedicularis rosea subsp. allionii (*) | Ped ros | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| CC—Phyteuma globulariifolium subsp. pedemontanum (*) | Phy glo | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| Ot.—Plantago alpina (*) | Pla alp | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| Ot.—Poa alpina (*) | Poa alp | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||
| DH—Saxifraga oppositifolia subsp. oppositifolia (*) | Sax opp | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| CC—Jacobaea incana (*) | Jac inc | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||
| TS | |||||||||||||||||||||||||||||||
| CC—Luzula lutea subsp. lutea (**) | Luz lut | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||
| SC—Potentilla crantzii subsp. crantzii (**) | Pot cra | 1 | 2 | 2 | 1 | 1 | |||||||||||||||||||||||||
| Ot.—Thymus praecox subsp. polytrichus (**) | Thy pra | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||
| CC—Agrostis rupestris subsp. rupestris (*) | Agr rup | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 1 | 1 | 3 | 1 | 2 | |||||||||||
| CC—Dianthus furcatus (*) | Dia fur | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| Ot.—Festuca rubra (*) | Fes rub | 1 | 1 | 1 | |||||||||||||||||||||||||||
| SC—Galium anisophyllon (*) | Gal ani | 1 | 1 | 1 | |||||||||||||||||||||||||||
| CC—Oreojuncus trifidus (*) | Ore tri | 2 | 2 | 1 | 1 | 1 | 4 | 1 | 1 | 1 | 1 | ||||||||||||||||||||
| SS | |||||||||||||||||||||||||||||||
| CC—Juncus jacquinii (**) | Jun jac | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| Ot.—Alchemilla vulgaris (*) | Alc vul | 1 | 1 | 1 | |||||||||||||||||||||||||||
| CC—Festuca halleri (*) | Fes hal | 1 | 1 | 1 | |||||||||||||||||||||||||||
| CC—Festuca scabriculmis subsp. luedii (*) | Fes lue | 4 | 2 | 2 | 1 | ||||||||||||||||||||||||||
| CC—Nardus stricta (*) | Nar str | 3 | 2 | 2 | 2 | ||||||||||||||||||||||||||
| Ot.—Pulsatilla alpina subsp. apiifolia (*) | Pul alp | 1 | 1 | 1 | |||||||||||||||||||||||||||
| CC—Sempervivum montanum (*) | Sem mon | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||
| Other species | |||||||||||||||||||||||||||||||
| OE—Carex myosuroides | Car myo | 2 | 3 | 1 | 3 | 2 | 1 | 1 | 1 | 2 | 1 | 2 | 2 | 1 | 1 | 1 | 2 | ||||||||||||||
| Ot.—Campanula scheuchzeri | Cam sch | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||
| Ot.—Carex sempervirens subsp. sempervirens | Car sem | 1 | 3 | 2 | 1 | 2 | 1 | 1 | 3 | 1 | 1 | 2 | 2 | 2 | 1 | 1 | |||||||||||||||
| Ot.—Lotus corniculatus subsp. alpinus | Lot cor | 1 | 2 | 2 | 1 | 2 | 1 | 2 | 1 | 1 | 1 | 1 | |||||||||||||||||||
| CC—Carex curvula subsp. curvula | Car cur | 3 | 1 | 1 | 1 | 2 | 2 | 1 | 1 | 1 | 1 | ||||||||||||||||||||
| CC—Euphrasia minima | Eup min | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||
| CC—Helictochloa versicolor subsp. versicolor | Hel ver | 2 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 3 | 1 | ||||||||||||||||||||
| CC—Gentianella ramosa | Gen ram | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||
| SC—Sesleria caerulea | Ses cae | 1 | 2 | 1 | 2 | 2 | 1 | 2 | 1 | 2 | |||||||||||||||||||||
| CC—Soldanella alpina subsp. alpina | Sol alp | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||
| Ot.—Pilosella officinarum | Pil off | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | ||||||||||||||||||||||
| CC—Trifolium alpinum | Tri alp | 2 | 2 | 2 | 2 | 2 | 1 | 3 | 1 | ||||||||||||||||||||||
| SC—Anthyllis vulneraria subsp. alpicola | Ant vul | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||
| SC—Myosotis alpestris | Myo alp | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||
| SC—Oxytropis montana | Oxy mon | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||
| AC—Carex parviflora | Car par | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||
| OE—Dryas octopetala subsp. octopetala | Dry oct | 4 | 1 | 7 | 1 | 1 | 1 | ||||||||||||||||||||||||
| SC—Festuca violacea | Fes vio | 1 | 1 | 1 | 2 | 3 | 3 | ||||||||||||||||||||||||
| CC—Minuartia recurva | Min rec | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||
| OE –Salix serpillifolia | Sal ser | 1 | 1 | 2 | 1 | 1 | 3 | ||||||||||||||||||||||||
| SC—Festuca pumila | Fes pum | 1 | 1 | 1 | 1 | 3 | |||||||||||||||||||||||||
| SC—Gentiana verna | Gen ver | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||
| CC—Geum montanum | Geu mon | 1 | 1 | 1 | 1 | 3 | |||||||||||||||||||||||||
| Ot.—Sempervivum arachnoideum | Sem ara | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||
| SC—Anemonoides baldensis | Ane bal | 2 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| OE—Arenaria ciliata | Are cil | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| SC—Aster alpinus subsp. alpinus | Ast alp | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| CC—Hieracium alpinum | Hie alp | 2 | 1 | 1 | 1 | ||||||||||||||||||||||||||
| Ot.—Achillea erba-rotta subsp. erba-rotta | Ach erb | 1 | 1 | 1 | |||||||||||||||||||||||||||
| CC—Armeria alpina | Arm alp | 1 | 1 | 1 | |||||||||||||||||||||||||||
| SH—Omalotheca supina | Oma sup | 1 | 1 | 1 | |||||||||||||||||||||||||||
| Ot.—Leucanthemum coronopifolium | Leu cor | 1 | 1 | 1 | |||||||||||||||||||||||||||
| CC—Cherleria sedoides | Che sed | 1 | 1 | 1 | |||||||||||||||||||||||||||
| SC—Sabulina verna | Sab ver | 1 | 1 | 1 | |||||||||||||||||||||||||||
| CC—Potentilla aurea subsp. aurea | Pot aur | 1 | 2 | 1 | |||||||||||||||||||||||||||
| SC—Saxifraga paniculata | Sax pan | 2 | 1 | 1 | |||||||||||||||||||||||||||
| SC—Trifolium thalii | Tri tha | 1 | 1 | 3 | |||||||||||||||||||||||||||
| CC—Veronica allionii | Ver all | 2 | 1 | 1 | |||||||||||||||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cazzavillan, A.; Gerdol, R.; Marrocchino, E.; Vaccaro, C.; Brancaleoni, L. Fine-Scale Lithogeochemical Features Influence Plant Distribution Patterns in Alpine Grasslands in the Western Alps of Italy. Plants 2024, 13, 2280. https://doi.org/10.3390/plants13162280
Cazzavillan A, Gerdol R, Marrocchino E, Vaccaro C, Brancaleoni L. Fine-Scale Lithogeochemical Features Influence Plant Distribution Patterns in Alpine Grasslands in the Western Alps of Italy. Plants. 2024; 13(16):2280. https://doi.org/10.3390/plants13162280
Chicago/Turabian StyleCazzavillan, Anna, Renato Gerdol, Elena Marrocchino, Carmela Vaccaro, and Lisa Brancaleoni. 2024. "Fine-Scale Lithogeochemical Features Influence Plant Distribution Patterns in Alpine Grasslands in the Western Alps of Italy" Plants 13, no. 16: 2280. https://doi.org/10.3390/plants13162280