
Species-Specific Responses of Bloom-Forming Algae to the Ocean Warming and Acidification

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
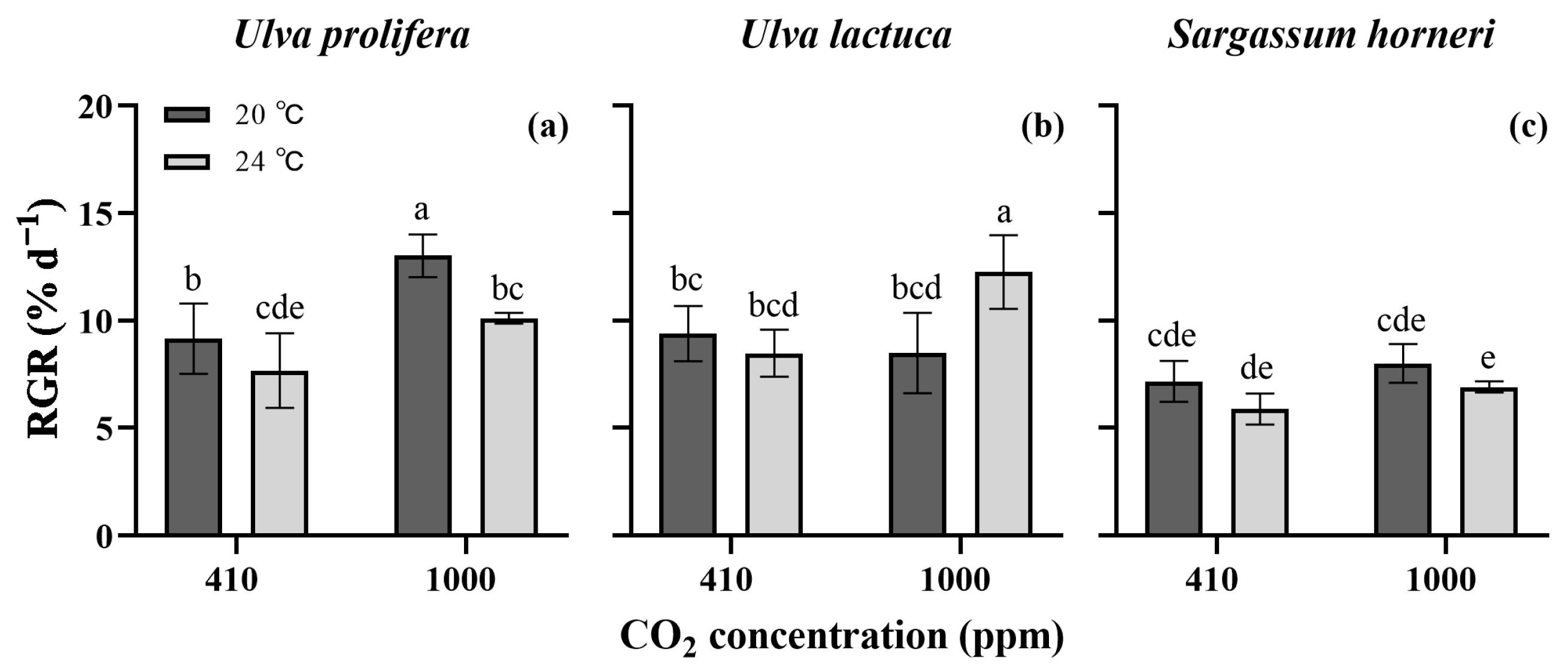
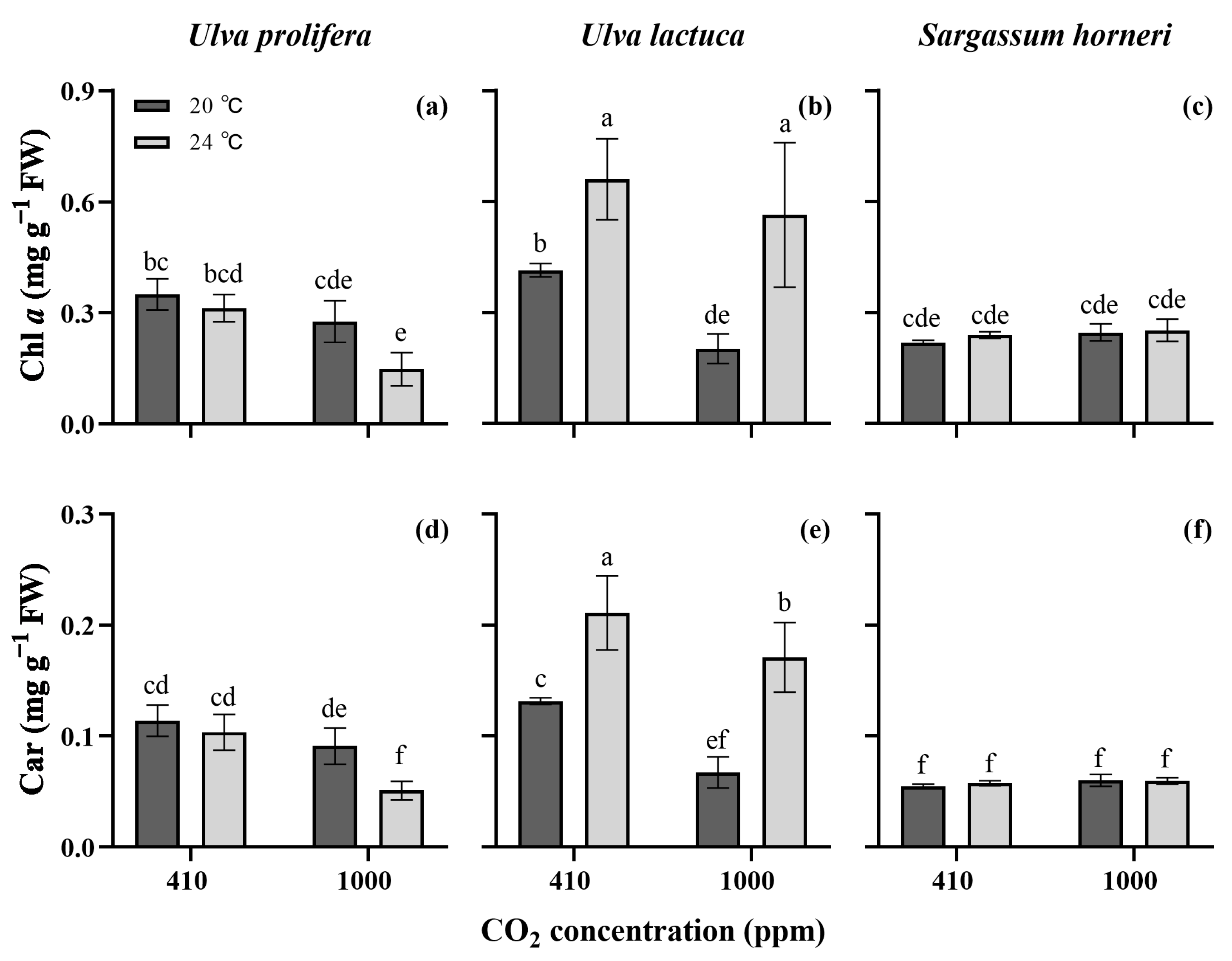
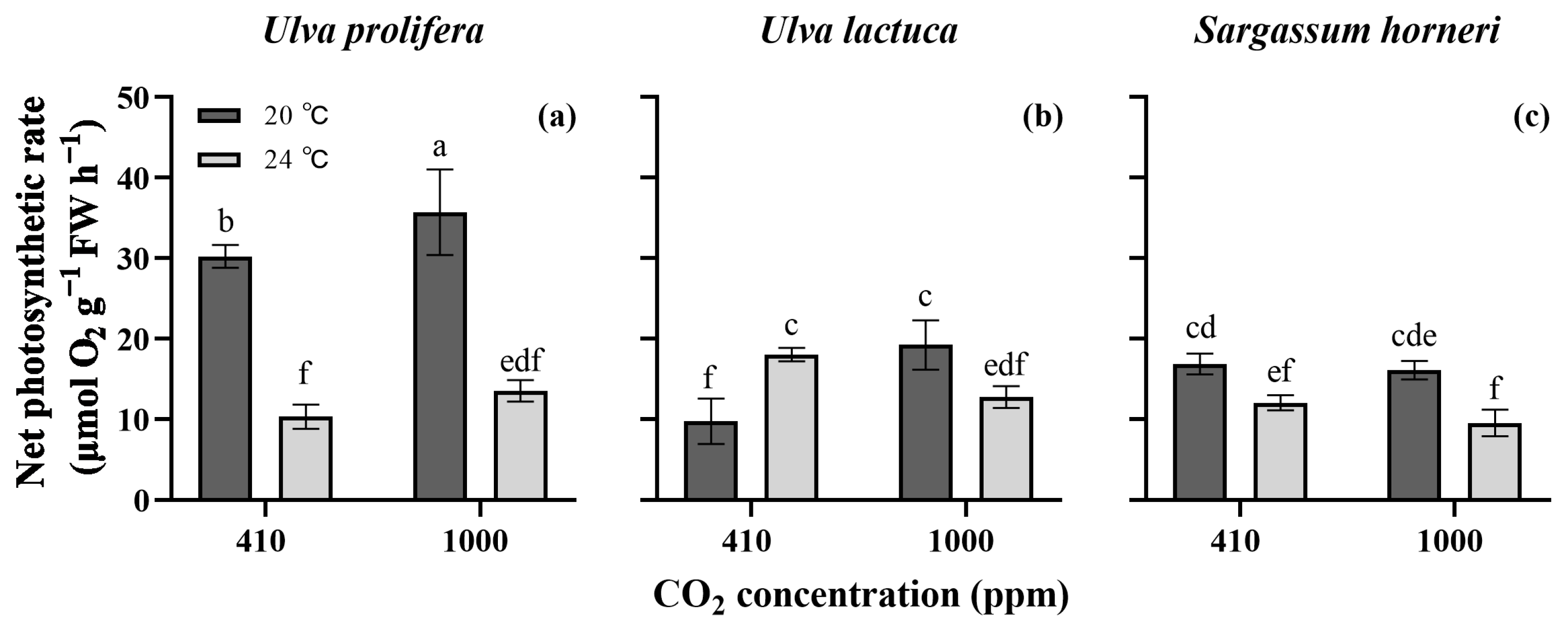
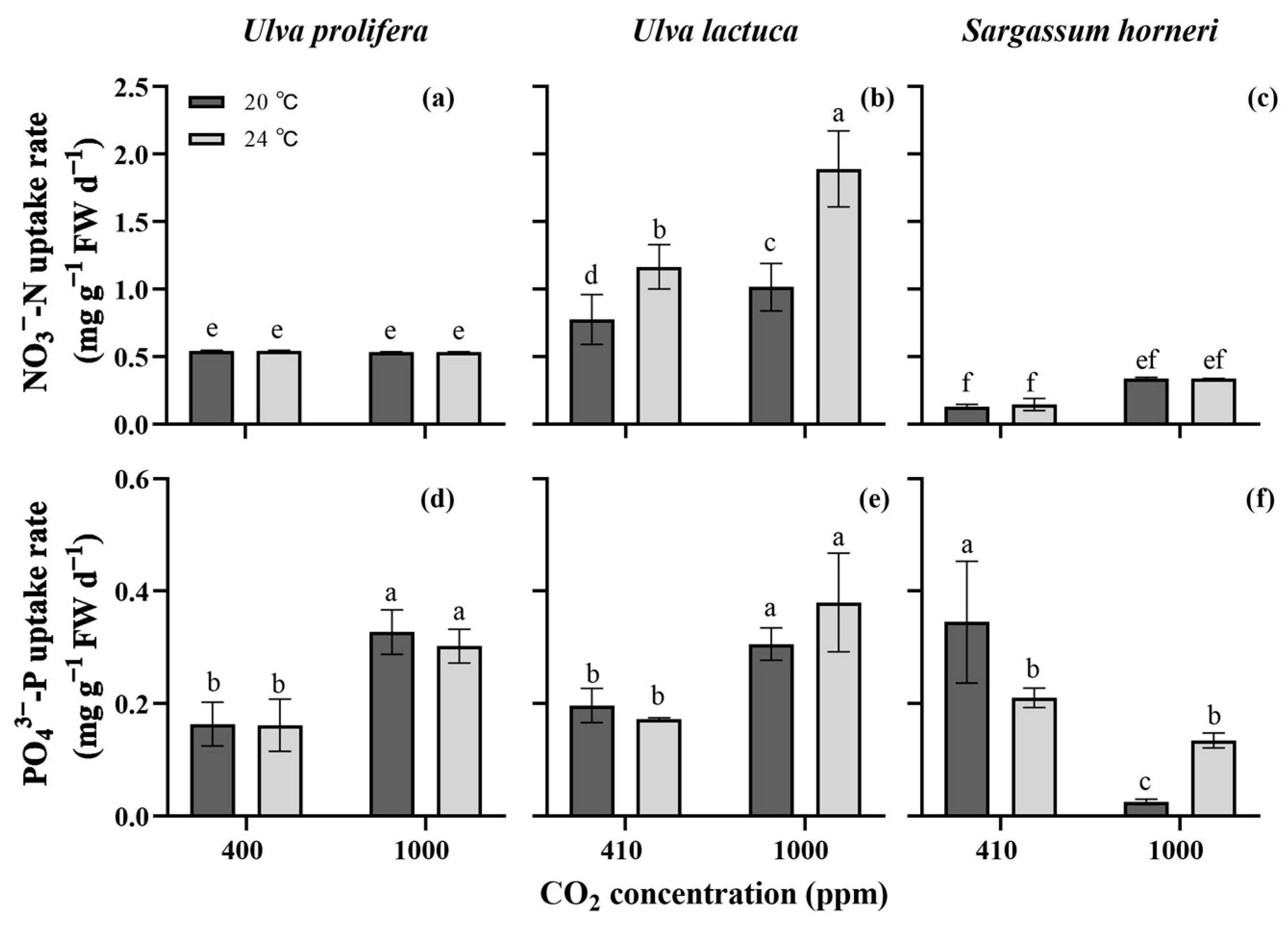
2. Results
2.1. Growth
2.2. Pigment Contents
2.3. Photosynthetic Rate
2.4. Nutrients Uptake Rates
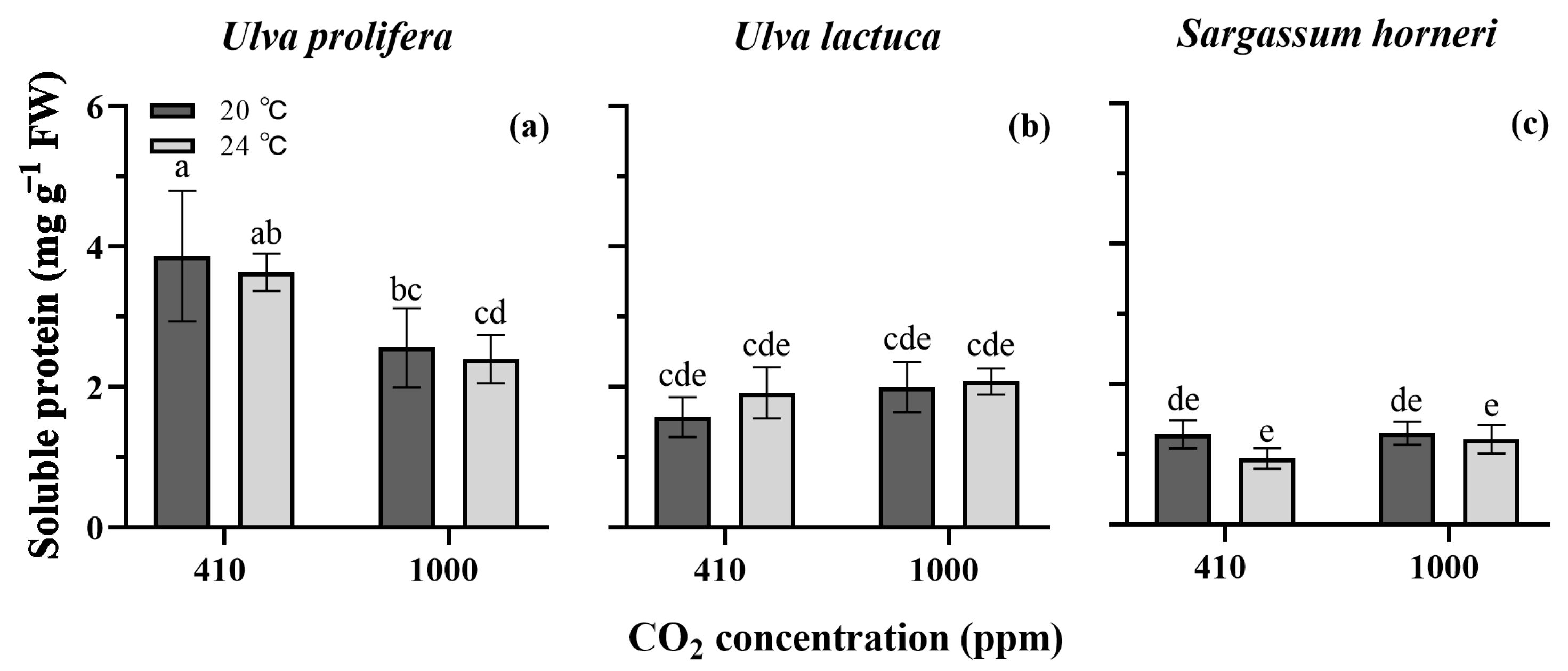
2.5. Soluble Protein Content
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Experimental Design
4.3. Growth Rate
4.4. Biochemical Components
4.5. Photosynthetic Oxygen Evolution
4.6. Nutrient Uptake Rates
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC The Intergovernmental Panel on Climate Change. Climate Change 2013: The Physical Science Basis. In Summary for Policymakers; Cambridge University Press: Cambridge, UK, 2013; pp. 24–25. [Google Scholar]
- Liu, L.; Zou, D.H.; Jiang, H.; Chen, B.B.; Zeng, X.P. Effects of Increased CO2 and Temperature on the Growth and Photosynthesis in the Marine Macroalga Gracilaria lemaneiformis from the Coastal Waters of South China. J. Appl. Phycol. 2018, 30, 1271–1280. [Google Scholar] [CrossRef]
- Scheffer, M.; Schulz, V.; Bellerby, R.G.J.; Botros, M.; Fritsche, P.; Meyerhöfer, M.; Neill, C.; Nondal, G.; Oschlies, A.; Wohlers, J.; et al. Positive Feedback between Global Warming and Atmospheric CO2 Concentration Inferred from Past Climate Change. Geophys. Res. Lett. 2006, 33, 229–237. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Burrows, M.T.; Brown, C.J.; García Molinos, J.; Halpern, B.S.; Hoegh-Guldberg, O.; Kappel, C.V.; Moore, P.J.; Richardson, A.J.; Schoeman, D.S.; et al. Responses of Marine Organisms to Climate Change across Oceans. Front. Mar. Sci. 2016, 3, 62. [Google Scholar] [CrossRef]
- Yao, C.-L.; Somero, G.N. The Impact of Ocean Warming on Marine Organisms. Chin. Sci. Bull. 2014, 59, 468–479. [Google Scholar] [CrossRef]
- Evans, R.D.; Wilson, S.K.; Field, S.N.; Moore, J.A.Y. Importance of Macroalgal Fields as Coral Reef Fish Nursery Habitat in North-west Australia. Mar. Biol. 2014, 161, 599–607. [Google Scholar] [CrossRef]
- Zhang, X.S.; Xu, D.; Guan, Z.; Wang, S.; Zhang, Y.; Wang, W.; Zhang, X.W.; Fan, X.; Li, F.; Ye, N. Elevated CO2 Concentrations Promote Growth and Photosynthesis of the Brown Alga Saccharina japonica. J. Appl. Phycol. 2020, 32, 1949–1959. [Google Scholar] [CrossRef]
- Fulton, C.J.; Berkström, C.; Wilson, S.K.; Abesamis, R.A.; Bradley, M.; Åkerlund, C.; Barrett, L.T.; Bucol, A.A.; Chacin, D.H.; Chong-Seng, K.M.; et al. Macroalgal Meadow Habitats Support Fish and Fisheries in Diverse Tropical Seascapes. Fish Fish. 2020, 21, 700–717. [Google Scholar] [CrossRef]
- Revilla-Lovano, S.; Sandoval-Gil, J.M.; Zertuche-González, J.A.; Belando-Torrentes, M.D.; Bernardeau-Esteller, J.; Rangel-Mendoza, L.K.; Ferreira-Arrieta, A.; Guzmán-Calderón, J.M.; Camacho-Ibar, V.F.; Muñiz-Salazar, R.; et al. Physiological Responses and Productivity of the Seaweed Ulva ohnoi (Chlorophyta) under Changing Cultivation Conditions in Pilot Large Land-Based Ponds. Algal Res. 2021, 56, 102316. [Google Scholar] [CrossRef]
- Borburema, H.D.S.; Graiff, A.; Marinho-Soriano, E.; Karsten, U. Photosynthetic Performance, Growth, Pigment Content, and Photoprotective Compounds of the Mangrove Macroalgae Bostrychia calliptera and Bostrychia montagnei (Rhodophyta) under Light Stress. Front. Mar. Sci. 2022, 9, 989454. [Google Scholar] [CrossRef]
- Zhang, W.; Shi, Y.; He, L.; Chen, X.; Hu, F.; Chen, Y.; Pang, Y.; Li, S.; Chu, Y. Decreased Salinity Offsets the Stimulation of Elevated pCO2 on Photosynthesis and Synergistically Inhibits the Growth of Juvenile Sporophyte of Saccharina japonica (Laminariaceae, Phaeophyta). Plants 2022, 11, 2978. [Google Scholar] [CrossRef]
- Raven, J.A.; Beardall, J. Carbon Acquisition Mechanisms of Algae: Carbon Dioxide Diffusion and Carbon Dioxide Concentrating Mechanisms. In Advances in Photosynthesis and Respiration; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 225–244. [Google Scholar]
- Laterre, R.; Pottier, M.; Remacle, C.; Boutry, M. Photosynthetic Trichomes Contain a Specific Rubisco with a Modified pH-Dependent Activity. Plant Physiol. 2017, 173, 2110–2120. [Google Scholar] [CrossRef]
- Liu, C.; Zou, D. Do Increased Temperature and CO2 Levels Affect the Growth, Photosynthesis, and Respiration of the Marine Macroalga Pyropia haitanensis (Rhodophyta)? An Experimental Study. Hydrobiologia 2015, 745, 285–296. [Google Scholar] [CrossRef]
- Raven, J. Inorganic Carbon Acquisition by Marine Autotrophs. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 1997; Volume 27, pp. 85–209. ISBN 978-0-12-005927-0. [Google Scholar]
- Wang, Y.; Xu, D.; Ma, J.; Zhang, X.; Fan, X.; Zhang, Y.; Wang, W.; Sun, K.; Ye, N. Elevated CO2 Accelerated the Bloom of Three Ulva Species after One Life Cycle Culture. J. Appl. Phycol. 2021, 33, 3963–3973. [Google Scholar] [CrossRef]
- Zhou, W.; Wu, H.; Huang, J.; Wang, J.; Zhen, W.; Wang, J.; Ni, J.; Xu, J. Elevated-CO2 and Nutrient Limitation Synergistically Reduce the Growth and Photosynthetic Performances of a Commercial Macroalga Gracilariopsis lemaneiformis. Aquaculture 2022, 550, 737878. [Google Scholar] [CrossRef]
- Liu, T.; Chen, J.A.; Wang, W.; Simon, M.; Wu, F.; Hu, W.; Chen, J.B.; Zheng, H. A Combined Proteomic and Transcriptomic Analysis on Sulfur Metabolism Pathways of Arabidopsis thaliana under Simulated Acid Rain. PLoS ONE 2014, 9, e90120. [Google Scholar] [CrossRef]
- Gordillo, F.J.L.; Carmona, R.; Viñegla, B.; Wiencke, C.; Jiménez, C. Effects of Simultaneous Increase in Temperature and Ocean Acidification on Biochemical Composition and Photosynthetic Performance of Common Macroalgae from Kongsfjorden (Svalbard). Polar Biol. 2016, 39, 1993–2007. [Google Scholar] [CrossRef]
- Wu, H.; Feng, J.; Li, X.; Zhao, C.; Liu, Y.; Yu, J.; Xu, J. Effects of Increased CO2 and Temperature on the Physiological Characteristics of the Golden Tide Blooming Macroalgae Sargassum horneri in the Yellow Sea, China. Mar. Pollut. Bull. 2019, 146, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, F.; Bonomi Barufi, J.; Malta, E.; Conde-Álvarez, R.; Nitschke, U.; Arenas, F.; Mata, M.; Connan, S.; Abreu, M.; Marquardt, R.; et al. Short-Term Effects of Increasing CO2, Nitrate and Temperature on Three Mediterranean Macroalgae: Biochemical Composition. Aquat. Biol. 2014, 22, 177–193. [Google Scholar] [CrossRef]
- Pospíšil, P. Production of Reactive Oxygen Species by Photosystem II as a Response to Light and Temperature Stress. Front. Plant Sci. 2016, 7, 1950. [Google Scholar] [CrossRef]
- Xiao, J.; Wang, Z.L.; Liu, D.Y.; Fu, M.Z.; Yuan, C.; Yan, T. Harmful Macroalgal Blooms (HMBs) in China’s Coastal Water: Green and Golden Tides. Harmful Algae 2021, 107, 102061. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Gao, L.; Huang, R.; Lin, X.; Gao, G. Differential Responses of Bloom-Forming Ulva intestinalis and Economically Important Gracilariopsis lemaneiformis to Marine Heatwaves under Changing Nitrate Conditions. Sci. Total Environ. 2022, 840, 156591. [Google Scholar] [CrossRef] [PubMed]
- Peter, N.R.; Raja, N.R.; Rengarajan, J.; Radhakrishnan Pillai, A.; Kondusamy, A.; Saravanan, A.K.; Changaramkumarath Paran, B.; Kumar Lal, K. A Comprehensive Study on Ecological Insights of Ulva lactuca Seaweed Bloom in a Lagoon along the Southeast Coast of India. Ocean Coast. Manag. 2024, 248, 106964. [Google Scholar] [CrossRef]
- Zhang, L.; Pei, H.; Chen, S.; Jiang, L.; Hou, Q.; Yang, Z.; Yu, Z. Salinity-Induced Cellular Cross-Talk in Carbon Partitioning Reveals Starch-to-Lipid Biosynthesis Switching in Low-Starch Freshwater Algae. Bioresour. Technol. 2018, 250, 449–456. [Google Scholar] [CrossRef]
- Halsey, K.H.; Jones, B.M. Phytoplankton Strategies for Photosynthetic Energy Allocation. Annu. Rev. Mar. Sci. 2015, 7, 265–297. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bera, S. Revisiting Nitrogen Utilization in Algae: A Review on the Process of Regulation and Assimilation. Bioresour. Technol. Rep. 2020, 12, 100584. [Google Scholar] [CrossRef]
- Shahar, B.; Shpigel, M.; Barkan, R.; Masasa, M.; Neori, A.; Chernov, H.; Salomon, E.; Kiflawi, M.; Guttman, L. Changes in Metabolism, Growth and Nutrient Uptake of Ulva fasciata (Chlorophyta) in Response to Nitrogen Source. Algal Res. 2020, 46, 101781. [Google Scholar] [CrossRef]
- Solovchenko, A.E.; Ismagulova, T.T.; Lukyanov, A.A.; Vasilieva, S.G.; Konyukhov, I.V.; Pogosyan, S.I.; Lobakova, E.S.; Gorelova, O.A. Luxury Phosphorus Uptake in Microalgae. J. Appl. Phycol. 2019, 31, 2755–2770. [Google Scholar] [CrossRef]
- Gao, G.; Beardall, J.; Wang, C.; Ren, W.; Xu, J. Ocean Acidification and Nutrient Limitation Synergistically Reduce Growth and Photosynthetic Performances of a Green Tide Alga Ulva linza. Biogeosciences 2018, 15, 3409–3420. [Google Scholar] [CrossRef]
- Cui, J.; Zhang, J.; Huo, Y.; Zhou, L.; Wu, Q.; Chen, L.; Yu, K.; He, P. Adaptability of Free-Floating Green Tide Algae in the Yellow Sea to Variable Temperature and Light Intensity. Mar. Pollut. Bull. 2015, 101, 660–666. [Google Scholar] [CrossRef]
- Muñoz, P.T.; Sáez, C.A.; Martínez-Callejas, M.B.; Flores-Molina, M.R.; Bastos, E.; Fonseca, A.; Gurgel, C.F.D.; Barufi, J.B.; Rörig, L.; Hall-Spencer, J.M.; et al. Eduardo Short-Term Interactive Effects of Increased Temperatures and Acidification on the Calcifying Macroalgae Lithothamnion crispatum and Sonderophycus capensis. Aquat. Bot. 2018, 148, 46–52. [Google Scholar] [CrossRef]
- Suárez-Álvarez, S.; Gómez-Pinchetti, J.L.; García-Reina, G. Effects of Increased CO2 Levels on Growth, Photosynthesis, Ammonium Uptake and Cell Composition in the Macroalga Hypnea spinella (Gigartinales, Rhodophyta). J. Appl. Phycol. 2012, 24, 815–823. [Google Scholar] [CrossRef]
- Stitt, M.; Krapp, A. The Interaction between Elevated Carbon Dioxide and Nitrogen Nutrition: The Physiological and Molecular Background. Plant Cell Environ. 1999, 22, 583–621. [Google Scholar] [CrossRef]
- Ahmadi, H.; Jamialahmadi, M.; Soulgani, B.S.; Dinarvand, N.; Sharafi, M.S. Experimental Study and Modelling on Diffusion Coefficient of CO2 in Water. Fluid Phase Equilibria 2020, 523, 112584. [Google Scholar] [CrossRef]
- Liu, C.; Zou, D.; Yang, Y. Comparative Physiological Behaviors of Ulva lactuca and Gracilariopsis lemaneiformis in Responses to Elevated Atmospheric CO2 and Temperature. Environ. Sci. Pollut. Res. 2018, 25, 27493–27502. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I. Photosynthetic Apparatus: Major Site of Oxidative Damage. In Reactive Oxygen Species in Plants: The Right Balance; Springer Nature: Singapore, 2023; pp. 75–92. [Google Scholar]
- Schlodder, E.; Meyer, B. pH Dependence of Oxygen Evolution and Reduction Kinetics of Photooxidized Chlorophyll aII (P-680) in Photosystem II Particles from Synechococcus sp. Biochim. Biophys. Acta BBA-Bioenerg. 1987, 890, 23–31. [Google Scholar] [CrossRef]
- Yu, J.; Li, J.; Wang, Q.; Liu, Y.; Gong, Q. Growth and Resource Accumulation of Drifting Sargassum horneri (Fucales, Phaeophyta) in Response to Temperature and Nitrogen Supply. J. Ocean Univ. China 2019, 18, 1216–1226. [Google Scholar] [CrossRef]
- IPCC. Climate Change, 2014: Synthesis Report; Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Li, S.F.; Liu, Y.; Gong, Q.L.; Gao, X.; Li, J.Y. Physiological and Ultrastructural Responses of the Brown Seaweed Undaria pinnatifida to Triphenyltin Chloride (TPTCL) Stress. Mar. Pollut. Bull. 2020, 153, 110978. [Google Scholar] [CrossRef]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as Well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Kochert, G. Protein Determination by Dye Binding; Cambridge University Press: London, UK, 1978. [Google Scholar]
- Liu, H.; Wang, F.; Dong, S.; Tian, X. A Comparative Study of the Nutrient Uptake and Growth Capacities of Seaweeds Caulerpa lentillifera and Gracilaria lichenoides. J. Appl. Phycol. 2016, 28, 3083–3089. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Cheng, F.; Chen, J.; Li, H.; Xu, J.; He, P.; Li, S. Species-Specific Responses of Bloom-Forming Algae to the Ocean Warming and Acidification. Plants 2024, 13, 2433. https://doi.org/10.3390/plants13172433
Wu H, Cheng F, Chen J, Li H, Xu J, He P, Li S. Species-Specific Responses of Bloom-Forming Algae to the Ocean Warming and Acidification. Plants. 2024; 13(17):2433. https://doi.org/10.3390/plants13172433
Chicago/Turabian StyleWu, Hailong, Fangsheng Cheng, Jiang Chen, He Li, Juntian Xu, Peimin He, and Sufang Li. 2024. "Species-Specific Responses of Bloom-Forming Algae to the Ocean Warming and Acidification" Plants 13, no. 17: 2433. https://doi.org/10.3390/plants13172433