
Contrasting Retromer with a Newly Described Retriever in Arabidopsis thaliana


{kind=link}
Abstract
1. Introduction
2. Fates of Endocytosed Cargo
3. Retromer in Yeast and Mammals
4. Plasma Membrane Protein Recycling by Retromer and Retriever
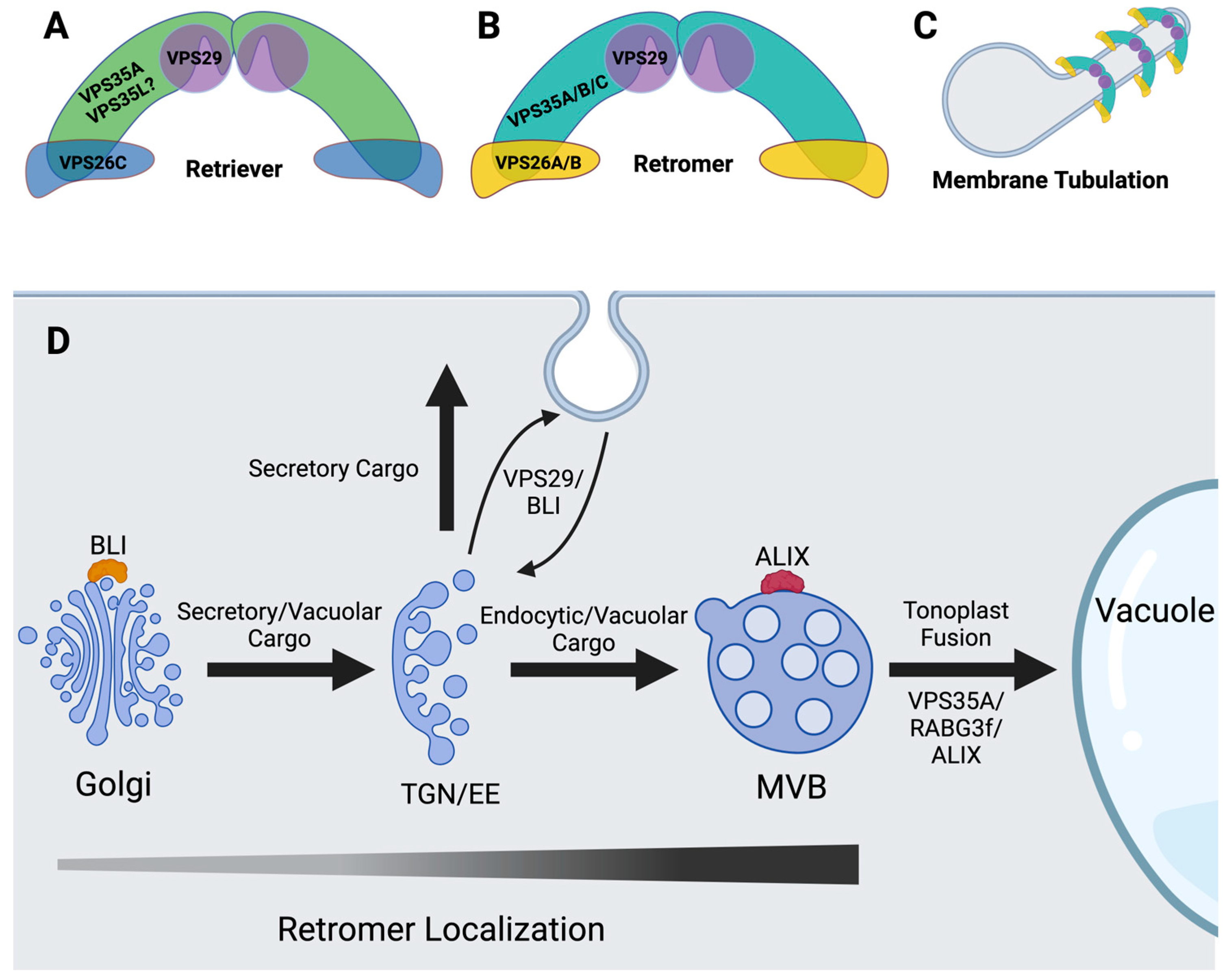
5. Plant Retromer and Retriever
6. Retromer-Binding Proteins
7. Is There a Retriever Complex in Plants?
8. Genetic Interactions between VTI11- and VTI13-Dependent Pathways to the Lytic Vacuole and Retromer/Retriever Function in Plants
9. Future Directions
- Is retriever involved in the recycling of plasma membrane proteins in plants and what are its cargoes?
- BLISTER and ALIX associate with retromer in Arabidopsis. What other proteins are associated with core retromer and retriever complexes in plants to modify/regulate their function?
- Do CCDC22 and CCDC93 associate with retriever alone or are they also involved in retromer function?
- Do CCDC22 and CCDC93 interact with other protein complexes in plants to compensate for the loss of COMMD proteins in seed plants?
- Are specific versions of the VPS35/29/26 complex required for cell-type-specific endosomal trafficking in plants?
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALIX | Apoptosis-linked gene-2-interacting protein X |
| BAR | Bin-Amphiphysin-Rvs |
| BLI | BLISTER |
| CCDC | Coiled-Coil Domain-Containing |
| CI-MPR | Cation-independent mannose 6-phosphate receptor |
| CLASP1 | Cytoplasmic linker-associated protein 1 |
| COMMD | Commander |
| CPY | Carboxypeptidase Y |
| DSCR3 | Down syndrome critical region 3 |
| EE | Early endosome |
| ER | Endoplasmic reticulum |
| ESCRT | Endosomal sorting complex required for transport |
| FREE1 | FYVE DOMAIN PROTEIN REQUIRED FOR ENDOSOMAL SORTING 1 |
| GEF | Guanine nucleotide exchange factor |
| GTPase | Guanosine triphosphate-activating protein |
| ILV | Intraluminal vesicles |
| LE | Late endosome |
| MBV | Multivesicular body |
| PI3P | Phosphatidylinositol 3-phosphate |
| PIN1 | Pin-formed |
| RAB | ras-associated binding |
| RFP | Red fluorescent protein |
| SCAR | Suppressor of cAMP receptor |
| SNX | Sorting nexin |
| TGN | trans-Golgi network |
| VPS | Vacuolar protein sorting |
| VSR | Vacuolar sorting receptor |
| VTI | VPS10 interacting |
| WASH | Wiskott–Aldrich syndrome |
References
- Fisher, T.J.; Flores-Sandoval, E.; Alvarez, J.P.; Bowman, J.L. PIN-FORMED is required for shoot phototropism/gravitropism and facilitates meristem formation in Marchantia polymorpha. New Phytol. 2023, 238, 1498–1515. [Google Scholar] [CrossRef] [PubMed]
- Jha, S.G.; Larson, E.R.; Humble, J.; Domozych, D.S.; Barrington, D.S.; Tierney, M.L. Vacuolar Protein Sorting 26C encodes an evolutionarily conserved large retromer subunit in eukaryotes that is important for root hair growth in Arabidopsis thaliana. Plant J. 2018, 94, 595–611. [Google Scholar] [CrossRef]
- Szumlanski, A.L.; Nielsen, E. The Rab GTPase RabA4d regulates pollen tube tip growth in Arabidopsis thaliana. Plant Cell 2009, 2, 526–544. [Google Scholar] [CrossRef] [PubMed]
- Dubeaux, G.; Neveu, J.; Zelazny, E.; Vert, G. Metal Sensing by the IRT1 Transporter-Receptor Orchestrates Its Own Degradation and Plant Metal Nutrition. Mol. Cell 2018, 69, 953–964.e5. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Koumoto, Y.; Li, L.; Yamazaki, M.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. AtVPS29, a Putative Component of a Retromer Complex, is Required for the Efficient Sorting of Seed Storage Proteins. Plant Cell Physiol. 2006, 47, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Munch, D.; Teh, O.-K.; Malinovsky, F.G.; Liu, Q.; Vetukuri, R.R.; Kasmi, F.E.; Brodersen, P.; Hara-Nishimura, I.; Dangl, J.L.; Petersen, M.; et al. Retromer contributes to immunity-associated cell death in Arabidopsis. Plant Cell 2015, 27, 463–479. [Google Scholar] [CrossRef]
- Yamazaki, M.; Shimada, T.; Takahashi, H.; Tamura, K.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. Arabidopsis VPS35, a Retromer Component, is Required for Vacuolar Protein Sorting and Involved in Plant Growth and Leaf Senescence. Plant Cell Physiol. 2008, 49, 142–156. [Google Scholar] [CrossRef]
- Cui, Y.; Yang, Z.; Teasdale, R.D. The functional roles of retromer in Parkinson’s disease. FEBS Lett. 2018, 7, 1096–1112. [Google Scholar] [CrossRef]
- Oliviusson, P.; Heinzerling, O.; Hillmer, S.; Hinz, G.; Tse, Y.C.; Jiang, L.; Robinson, D.G. Plant retromer, localized to the prevacuolar compartment and microvesicles in Arabidopsis, may interact with vacuolar sorting receptors. Plant Cell. 2006, 5, 1239–1252. [Google Scholar] [CrossRef]
- McNally, K.E.; Faulkner, R.; Steinberg, F.; Gallon, M.; Ghai, R.; Pim, D.; Langton, P.; Pearson, N.; Danson, C.M.; Nägele, H.; et al. Retriever is a multiprotein complex for retromer-independent endosomal cargo recycling. Nat. Cell Biol. 2017, 19, 1214–1225. [Google Scholar] [CrossRef]
- Rodriguez-Furlan, C.; Minina, E.A.; Hicks, G.R. Remove, Recycle, Degrade: Regulating Plasma Membrane Protein Accumulation. Plant Cell 2019, 31, 2833–2854. [Google Scholar] [CrossRef]
- Cui, Y.; Shen, J.; Gao, C.; Zhuang, X.; Wang, J.; Jiang, L. Biogenesis of Plant Prevacuolar Multivesicular Bodies. Mol. Plant 2016, 9, 774–786. [Google Scholar] [CrossRef]
- Cai, Q.; Qiao, L.; Wang, M.; He, B.; Lin, F.M.; Palmquist, J.; Huang, S.D.; Jin, H. Plants send small RNAs in extracellular vesicles to fungal pathogen to silence virulence genes. Science 2018, 360, 1126–1129. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Li, B.; Huang, S.; Li, H.; Cao, W.; Chen, Y.; Liu, G.; Li, Z.; Yang, C.; Feng, L.; et al. The plant unique ESCRT component FREE1 regulates autophagosome closure. Nat. Commun. 2023, 14, 1768. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Zhuang, X.; Shen, J.; Jiang, L. Plant ESCRT Complexes: Moving Beyond Endosomal Sorting. Trends Plant Sci. 2017, 22, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Viotti, C.; Bubeck, J.; Stierhof, Y.D.; Krebs, M.; Langhans, M.; van den Berg, W.; van Dongen, W.; Richter, S.; Geldner, N.; Takano, J.; et al. Endocytic and secretory traffic in Arabidopsis merge in the trans-Golgi network/early endosome, an independent and highly dynamic organelle. Plant Cell 2010, 22, 1344–1357. [Google Scholar] [CrossRef]
- Grant, B.; Donaldson, J. Pathways and mechanisms of endocytic recycling. Nat. Rev. Mol. Cell Biol. 2009, 10, 597–608. [Google Scholar] [CrossRef]
- Kleine-Vehn, J.; Wabnik, K.; Martinière, A.; Langowski, L.; Willig, K.; Naramoto, S.; Leitner, J.; Tanaka, H.; Jakobs, S.; Robert, S.; et al. Recycling, clustering, and endocytosis jointly maintain PIN auxin carrier polarity at the plasma membrane. Mol. Syst. Biol. 2011, 7, 540. [Google Scholar] [CrossRef]
- Geldner, N.; Hyman, D.L.; Wang, X.; Schumacher, K.; Chory, J. Endosomal signaling of plant steroid receptor kinase BRI1. Genes. Dev. 2007, 21, 1598–1602. [Google Scholar] [CrossRef]
- Chinchilla, D.; Bauer, Z.; Regenass, M.; Boller, T.; Felix, G. The Arabidopsis receptor kinase FLS2 binds flg22 and determines the specificity of flagellin perception. Plant Cell 2006, 18, 465–476. [Google Scholar] [CrossRef]
- Takano, J.; Tanaka, M.; Toyoda, A.; Miwa, K.; Kasai, K.; Fuji, K.; Onouchi, H.; Naito, S.; Fujiwara, T. Polar localization and degradation of Arabidopsis boron transporters through distinct trafficking pathways. Proc. Natl. Acad. Sci. USA 2010, 107, 5220–5225. [Google Scholar] [CrossRef] [PubMed]
- Barberon, M.; Dubeaux, G.; Kolb, C.; Isono, E.; Zelazny, E.; Vert, G. Polarization of IRON-REGULATED TRANSPORTER 1 (IRT1) to the plant-soil interface plays crucial role in metal homeostasis. Proc. Natl. Acad. Sci USA 2014, 111, 8293–8298. [Google Scholar] [CrossRef] [PubMed]
- Jaillais, Y.; Santambrogio, M.; Rozier, F.; Fobis-Loisy, I.; Miège, C.; Gaude, T. The retromer protein VPS29 links cell polarity and organ initiation in plants. Cell 2007, 130, 1057–1070. [Google Scholar] [CrossRef]
- Seaman, M.N.; Marcusson, E.G.; Cereghino, J.L.; Emr, S.D. Endosome to Golgi retrieval of the vacuolar protein sorting receptor, Vps10p, requires the function of the VPS29, VPS30, and VPS35 gene products. J. Cell Biol. 1997, 137, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Seaman, M.N.; McCaffery, J.M.; Emr, S.D. A membrane coat complex essential for endosome-to-Golgi retrograde transport in yeast. J. Cell Biol. 1998, 142, 665–681. [Google Scholar] [CrossRef]
- Seaman, M.N.; Williams, H.P. Identification of the functional domains of yeast sorting nexins Vps5p and Vps17p. Mol. Biol. Cell. 2002, 13, 2826–2840. [Google Scholar] [CrossRef]
- van Weering, J.R.; Verkade, P.; Cullen, P.J. SNX-BAR proteins in phosphoinositide-mediated, tubular-based endosomal sorting. Semin. Cell Dev. Biol. 2010, 21, 371–380. [Google Scholar] [CrossRef]
- Chen, K.E.; Healy, M.D.; Collins, B.M. Towards a molecular understanding of endosomal trafficking by Retromer and Retriever. Traffic 2019, 20, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Koumandou, V.L.; Klute, M.J.; Herman, E.K.; Nunez-Miguel, R.; Dacks, J.B.; Field, M.C. Evolutionary reconstruction of the retromer complex and its function in Trypanosoma brucei. J. Cell Sci. 2011, 124, 1496–1509. [Google Scholar] [CrossRef]
- Simonetti, B.; Danson, C.M.; Heesom, K.J.; Cullen, P.J. Sequence-dependent cargo recognition by SNX-BARs mediates retromer-independent transport of CI-MPR. J. Cell Biol. 2017, 216, 3695–3712. [Google Scholar] [CrossRef]
- Meyer, C.; Zizioli, D.; Lausmann, S.; Eskelinen, E.L.; Hamann, J.; Saftig, P.; von Figura, K.; Schu, P. mu1A-adaptin-deficient mice: Lethality, loss of AP-1 binding and rerouting of mannose 6-phosphate receptors. EMBO J. 2000, 19, 2193–2203. [Google Scholar] [CrossRef] [PubMed]
- Kvainickas, A.; Jimenez-Orgaz, A.; Nagele, H.; Hu, Z.; Dengjel, J.; Steinberg, F. Cargo-selective SNX-BAR proteins mediate retromer trimer independent retrograde transport. J. Cell Biol. 2017, 216, 3677–3693. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Carosi, J.M.; Yang, Z.; Ariotti, N.; Kerr, M.C.; Parton, R.G.; Sargeant, T.J.; Teasdale, R.D. Retromer has a selective function in cargo sorting via endosome transport carriers. J. Cell Biol. 2019, 218, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Temkin, P.; Lauffer, B.; Jäger, S.; Cimermancic, P.; Krogan, N.J.; von Zastrow, M. SNX27 mediates retromer tubule entry and endosome-to-plasma membrane trafficking of signalling receptors. Nat. Cell Biol. 2011, 13, 715–721. [Google Scholar] [CrossRef]
- Gallon, M.; Cullen, P.J. Retromer and sorting nexins in endosomal sorting. Biochem. Soc. Trans. 2015, 43, 33–47. [Google Scholar] [CrossRef]
- Harrison, M.S.; Hung, C.S.; Liu, T.T.; Christiano, R.; Walther, T.C.; Burd, C.G. A mechanism for retromer endosomal coat complex assembly with cargo. Proc. Natl. Acad. Sci. USA 2014, 111, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Gallon, M.; Clairfeuille, T.; Steinberg, F.; Mas, C.; Ghai, R.; Sessions, R.B.; Teasdale, R.D.; Collins, B.M.; Cullen, P.J. A unique PDZ domain and arrestin-like fold interaction reveals mechanistic details of endocytic recycling by SNX27-retromer. Proc. Natl. Acad. Sci. USA 2014, 111, E3604–E3613. [Google Scholar] [CrossRef]
- Yong, X.; Zhao, L.; Hu, W.; Sun, Q.; Ham, H.; Liu, Z.; Ren, J.; Zhang, Z.; Zhou, Y.; Yang, Q.; et al. SNX27-FERM-SNX1 complex structure rationalizes divergent trafficking pathways by SNX17 and SNX27. Proc. Natl. Acad. Sci. USA 2021, 118, e2105510118. [Google Scholar] [CrossRef]
- Hu, S.; Li, B.; Wu, F.; Zhu, D.; Zouhar, J.; Gao, C.; Shimada, T.; Rojo, E.; Hara-Nishimura, I.; Jiang, L.; et al. Plant ESCRT protein ALIX coordinates with retromer complex in regulating receptor-mediated sorting of soluble vacuolar proteins. Proc. Natl. Acad. Sci. USA 2022, 119, e2200492119. [Google Scholar] [CrossRef]
- Nodzynski, T.; Feraru, M.I.; Hirsch, S.; De Rycke, R.; Niculaes, C.; Boerjan, W.; Van Leene, J.; De Jaeger, G.; Vanneste, S.; Friml, J. Retromer subunits VPS35A and VPS29 mediate prevacuolar compartment (PVC) function in Arabidopsis. Mol. Plant 2013, 6, 1849–1862. [Google Scholar] [CrossRef]
- Niemes, S.; Labs, M.; Scheuring, D.; Krueger, F.; Longhand, M.; Jesenofsky, B.; Robinson, D.G.; Pimpl, P.N. Sorting of plant vacuolar proteins is initiated in the ER. Plant J. 2010, 62, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Niemes, S.; Longhans, M.; Viotti, C.; Scheuring, D.; Yan, M.S.W.; Jiang, L.; Hillmer, S.; Robinson, D.G.; Pimpl, P. Retromer recycles vacuolar sorting receptors from the trans-Golgi network. Plant J. 2010, 61, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Xu, H.; Pimpl, P. Nanobody-based VSR7 tracing shows clattering-dependent TGN to Golgi recycling. Nat. Commun. 2023, 14, 6926. [Google Scholar] [CrossRef]
- Salanenka, Y.; Verstraeten, I.; Löfke, C.; Tabata, K.; Naramoto, S.; Glanc, M.; Friml, J. Gibberellin DELLA signaling targets the retromer complex to redirect protein trafficking to the plasma membrane. Proc. Natl. Acad. Sci. USA 2018, 115, 3716–3721. [Google Scholar] [CrossRef]
- Chen, L.; Li, Y.X.; Li, C.; Shi, Y.; Song, Y.; Zhang, D.; Wang, H.; Li, Y.; Wang, T. The retromer protein ZmVPS29 regulates maize kernel morphology likely through an auxin-dependent process(es). Plant Biotechnol. J. 2020, 18, 1004–1014. [Google Scholar] [CrossRef]
- Zelazny, E.; Santambrogio, M.; Pourcher, M.; Chambrier, P.; Berne-Dedieu, A.; Fobis-Loisy, I.; Miège, C.; Jaillais, Y.; Gaude, T. Mechanisms governing the endosomal membrane recruitment of the core retromer in Arabidopsis. J. Biol. Chem. 2013, 288, 8815–8825. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Huang, R.; Liao, Y.; Yang, S.; Feng, B.; Qin, H.; Zhou, J.; Zeng, Y.; Shen, J.; Zhuang, X.; et al. A plant-unique protein BLISTER coordinates with core retromer to modulate endosomal sorting of plasma membrane and vacuolar proteins. Proc. Natl. Acad. Sci. USA 2023, 120, e2211258120. [Google Scholar] [CrossRef]
- Hashiguchi, Y.; Niihama, M.; Takahashi, T.; Saito, C.; Nakano, A.; Tasaka, M.; Morita, M.T. Loss-of-function mutations of retromer large subunit genes suppress the phenotype of an Arabidopsis zig mutant that lacks Qb-SNARE VTI11. Plant Cell 2010, 22, 159–172. [Google Scholar] [CrossRef]
- Cui, Y.; Zhao, Q.; Gao, C.; Ding, Y.; Zeng, Y.; Ueda, T.; Nakano, A.; Jiang, L. Activation of the Rab7 GTPase by the MON1-CCZ1 Complex Is Essential for PVC-to-Vacuole Trafficking and Plant Growth in Arabidopsis. Plant Cell 2014, 26, 2080–2097. [Google Scholar] [CrossRef]
- Takemoto, K.; Ebine, K.; Askani, J.C.; Krüger, F.; Gonzalez, Z.A.; Ito, E.; Goh, T.; Schumacher, K.; Nakano, A.; Ueda, T. Distinct sets of tethering complexes, SNARE complexes, and Rab GTPases mediate membrane fusion at the vacuole in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E2457–E2466. [Google Scholar] [CrossRef]
- Rodriguez-Furlan, C.; Domozych, D.; Qian, W.; Enquist, P.A.; Li, X.; Zhang, C.; Schenk, R.; Winbigler, H.S.; Jackson, W.; Raikhel, N.V.; et al. Interaction between VPS35 and RABG3f is necessary as a checkpoint to control fusion of late compartments with the vacuole. Proc. Natl. Acad. Sci. USA 2019, 116, 21291–21301. [Google Scholar] [CrossRef] [PubMed]
- Laulumaa, S.; Kampala, E.-P.; Huskonen, J.T.; Varjosalo, M. Structure and interactions of the endogenous human Commander complex. Nat. Struct. Mol. Biol. 2024, 31, 925–938. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.D.; Preston, J.C.; Tierney, M.L. CCDC22 and CCDC93, two potential retriever-interacting proteins, are required for root and root hair growth in Arabidopsis. Front. Plant Sci. 2022, 13, 1051503. [Google Scholar] [CrossRef] [PubMed]
- Larson, E.R.; Domozych, D.S.; Tierney, M.L. SNARE VTI13 plays a unique role in endosomal trafficking pathways associated with the vacuole and is essential for cell wall organization and root hair growth in Arabidopsis. Ann. Bot. 2014, 114, 1147–1159. [Google Scholar] [CrossRef] [PubMed]
- Yano, D.; Sato, M.; Saito, C.; Sato, M.H.; Morita, M.T.; Tasaka, M. A SNARE complex containing SGR3/AtVAM3 and ZIG/VTI11 in gravity-sensing cells is important for Arabidopsis shoot gravitropism. Proc. Natl. Acad. Sci. USA 2003, 100, 8589–8594. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lewis, C.D.; Tierney, M.L. Contrasting Retromer with a Newly Described Retriever in Arabidopsis thaliana. Plants 2024, 13, 2470. https://doi.org/10.3390/plants13172470
Lewis CD, Tierney ML. Contrasting Retromer with a Newly Described Retriever in Arabidopsis thaliana. Plants. 2024; 13(17):2470. https://doi.org/10.3390/plants13172470
Chicago/Turabian StyleLewis, Connor D., and Mary L. Tierney. 2024. "Contrasting Retromer with a Newly Described Retriever in Arabidopsis thaliana" Plants 13, no. 17: 2470. https://doi.org/10.3390/plants13172470
APA StyleLewis, C. D., & Tierney, M. L. (2024). Contrasting Retromer with a Newly Described Retriever in Arabidopsis thaliana. Plants, 13(17), 2470. https://doi.org/10.3390/plants13172470