

Metabolomics Analysis of Phenolic Composition and Content in Five Pear Cultivars Leaves


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Qualitative Analysis of Metabolites in Leaves
2.2. Quantitative Analysis and Trend Assessment of Metabolite Expression Levels
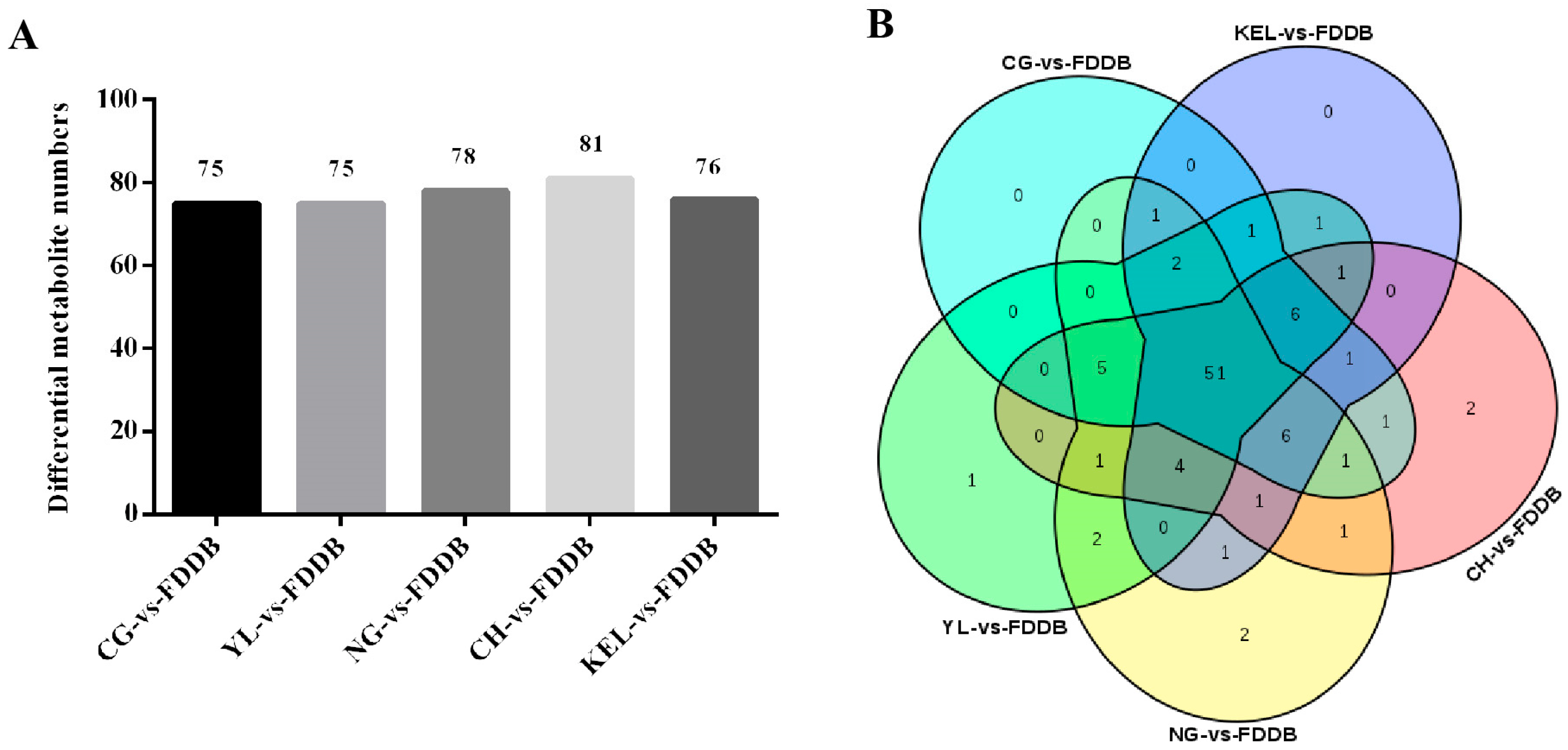
2.3. Screening and Analysis of Differential Metabolites
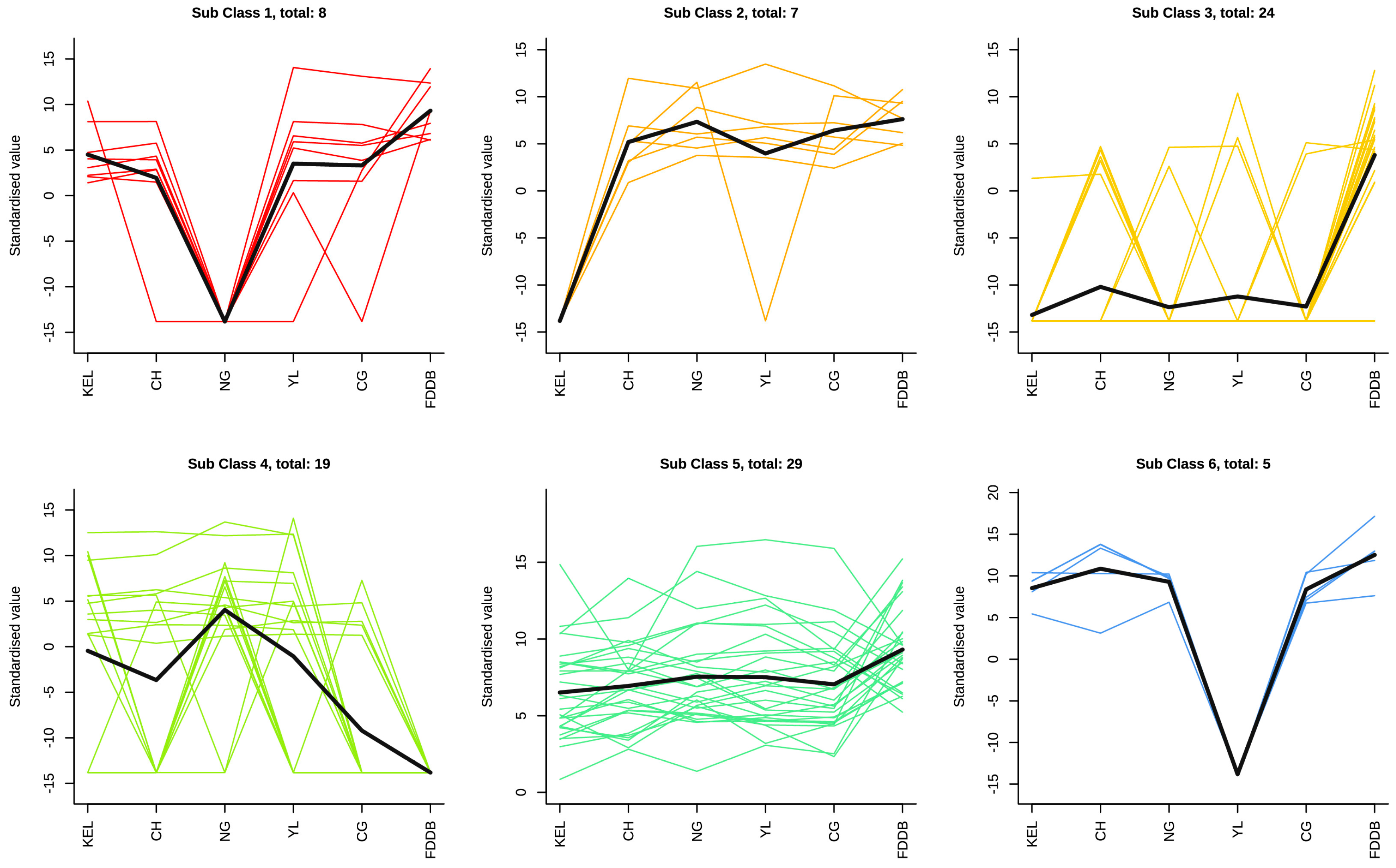
2.4. K-Means Analysis of Differential Metabolites
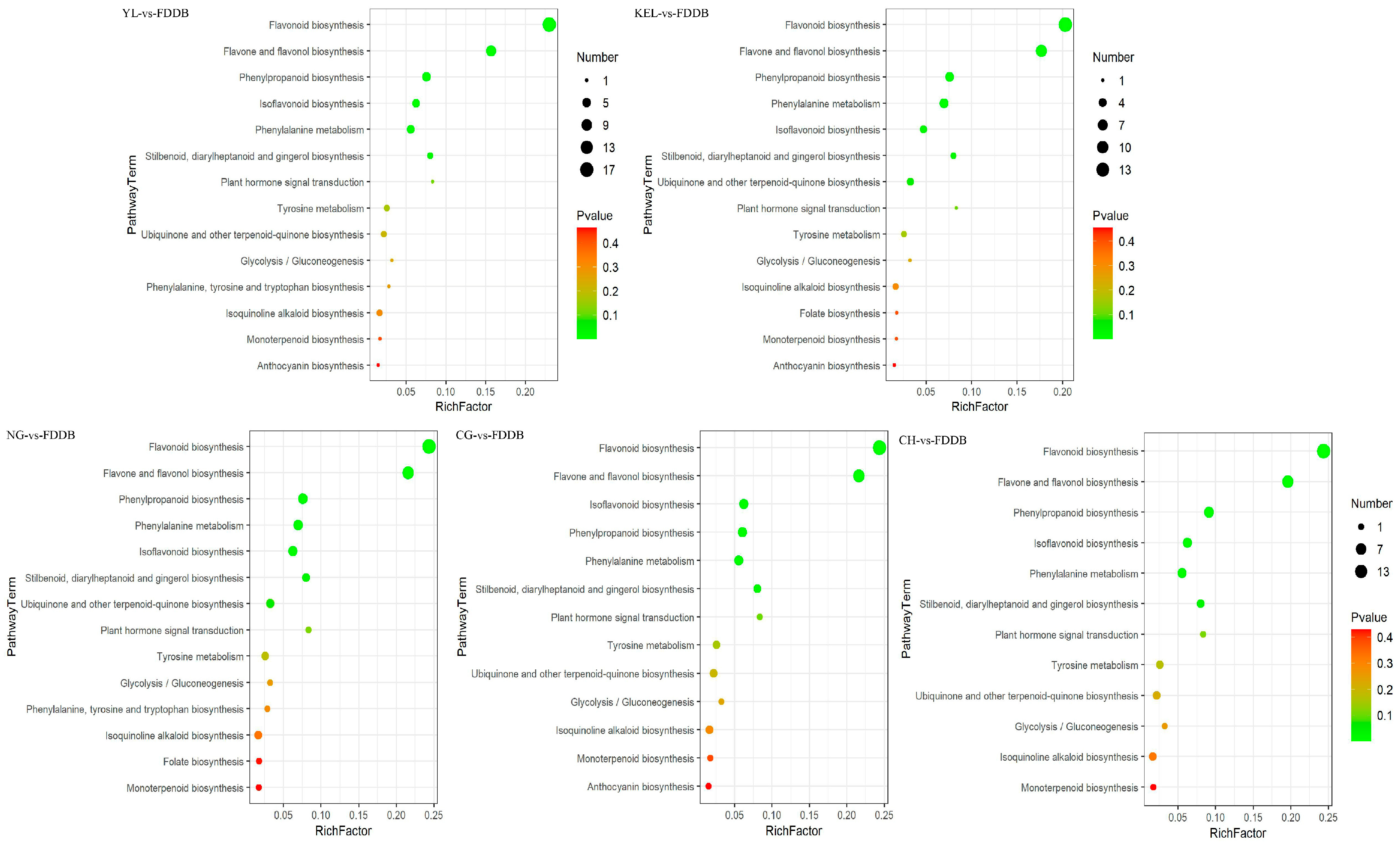
2.5. KEGG Analysis of Differential Metabolites
2.6. Differential Metabolite Correlation Analysis
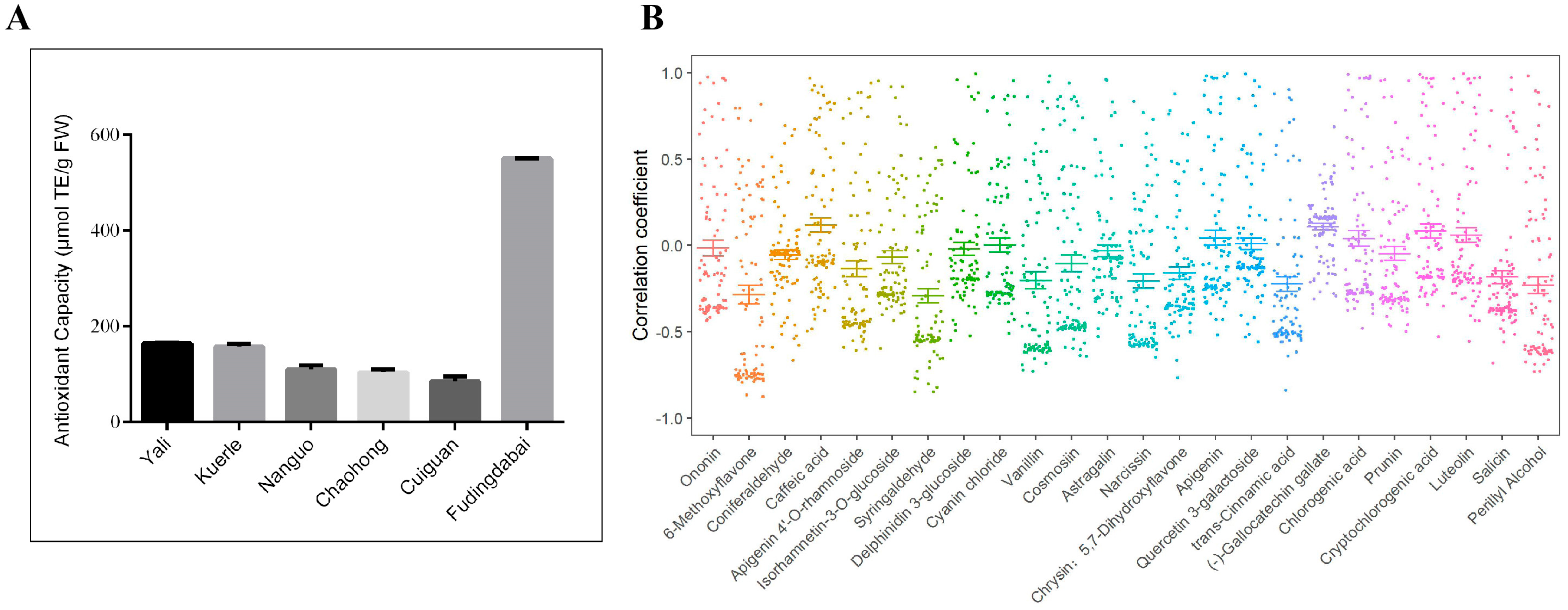
2.7. Evaluation of Antioxidant Capacity of Leaves
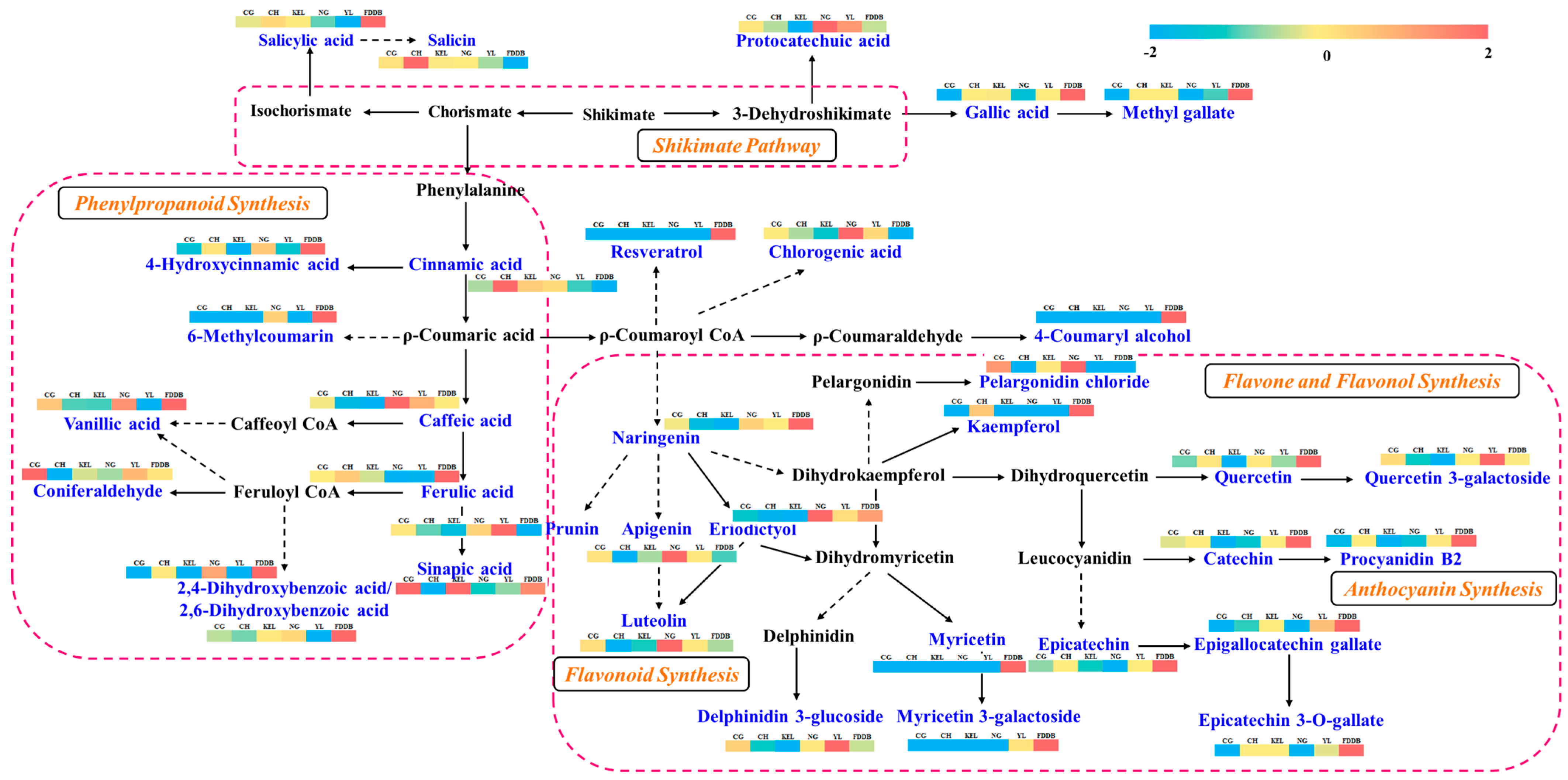
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Metabolite Extraction
4.3. Chromatography Mass Spectrometry
4.4. Qualitative and Quantitative Analysis of Metabolites
4.5. Differential Metabolite Screening
4.6. Heatmap and Venn Analysis
4.7. K-Means Analysis
4.8. KEGG Analysis
4.9. DPPH (1,1-Diphenyl-2-picrylhydrazyl Radical) Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, J.; Wang, Y.T.; Xu, J.B.; Korban, S.S.; Fei, Z.J.; Tao, S.T.; Ming, R.; Tai, S.S.; Khan, A.M.; Postman, J.D.; et al. Diversification and independent domestication of Asian and European pears. Genome Biol. 2018, 19, 77. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, T.T.; Zhou, B.; Gao, W.Y.; Cao, J.G.; Huang, L.Q. Chemical composition and antioxidant and anti-inflammatory potential of peels and flesh from 10 different pear varieties (Pyrus spp.). Food Chem. 2014, 152, 531–538. [Google Scholar] [CrossRef]
- Li, X.; Zhang, J.Y.; Gao, W.Y.; Wang, T.T.; Wang, H.Y.; Cao, J.G.; Huang, L.Q. Chemical composition and anti-inflammatory and antioxidant activities of eight pear cultivars. J. Agri. Food Chem. 2012, 60, 8738–8744. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Yang, C.Q.; Wei, Y.K.; Qi-Xia, M.A.; Yang, L.; Chen, X.Y. Genomics grand for diversified plant secondary metabolites. Plant Divers. 2011, 33, 53. [Google Scholar]
- Kolniak-Ostek, J.; Kopotowska, D.; Rutkowski, K.P.; Skorupińska, A.; Kruczyńska, D.E. Bioactive compounds and health-promoting properties of pear (Pyrus communis L.) fruits. Molecules 2020, 25, 4444. [Google Scholar] [CrossRef] [PubMed]
- Carbonaro, M.; Mattera, M.; Nicoli, S.; Bergamo, P.; Cappelloni, M. Modulation of antioxidant compounds in organic vs conventional fruit (peach, Prunus persica L., and pear, Pyrus communis L.). J. Agri. Food Chem. 2002, 50, 5458–5462. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, X.; Yu, J.; Pu, Y.F.; Lin, C.X.; Wu, C.Y. Dynamics of phenolic substances and their antioxidant activity during fruit development of ‘Lyubaoshi’ pears. J. Tarim Univ. 2021, 33, 25–31. [Google Scholar]
- Kolniak-Ostek, J. Content of bioactive compounds and antioxidant capacity in skin tissues of pear. J. Funct. Foods 2016, 23, 40–51. [Google Scholar] [CrossRef]
- Wang, Z.N.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. A comparative investigation on phenolic composition, characterization and antioxidant potentials of five different Australian grown pear varieties. Antioxidants 2021, 10, 151. [Google Scholar] [CrossRef]
- Cui, T.; Nakamura, K.; Ma, L.; Li, J.Z.; Kayahara, H. Analyses of arbutin and chlorogenic acid, the major phenolic constituents in Oriental pear. J. Agric. Food Chem. 2005, 53, 3882–3887. [Google Scholar] [CrossRef]
- Lee, K.H.; Cho, J.Y.; Lee, H.J.; Park, K.Y.; Ma, Y.K.; Lee, S.H.; Cho, J.A.; Kim, W.S.; Park, K.H.; Moon, J.H. Isolation and identification of phenolic compounds from an Asian pear (Pyrus pyrifolia Nakai) fruit peel. Food Sci. Biotechnol. 2011, 20, 1539–1545. [Google Scholar] [CrossRef]
- Li, J.; Nie, J.Y.; Cao, Y.F.; Li, Z.X.; Yan, Z.; Wu, Y.L. UPLC–PDA–MS/MS–ESI Analysis of phenolic compounds in fruits of DangshanSuli and Qiubaili Pears (Pyrus bretschneideri). Acta Hortic. Sinica 2016, 43, 752–762. [Google Scholar]
- Andreotti, C.; Costa, G.; Treutter, D. Composition of phenolic compounds in pear leaves as affected by genetics, ontogenesis and the environment. Sci. Hortic-Amst. 2006, 109, 130–137. [Google Scholar] [CrossRef]
- Park, Y.O.; Choi, J.J.; Choi, J.H. Antioxidant activities of young and mature fruit in three Asian pear cultivars. Korean J. Hortic. Sci. Technol. 2012, 30, 208–213. [Google Scholar]
- Mcclure, A.K.; Gong, Y.H.; Song, J.; Vinqvist-Tymchuk, M.; Palmer, L.C.; Fan, L.H.; Burgher-Maclellan, K.; Zhang, Z.Q.; Celton, J.M.; Forney, C.F.; et al. Genome-wide association studies in apple reveal loci of large effect controlling apple polyphenols. Hortic. Res. 2019, 1, 444–455. [Google Scholar] [CrossRef] [PubMed]
- Burton-Freeman, B.M.; Sandhu, A.K.; Edirisinghe, I. Red raspberries and their bioactive polyphenols: Cardiometabolic and neuronal health links. Adv. Nutr. 2016, 7, 44–65. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Tang, Z.H.; Wu, C.Y.; Wang, X.; Pu, Y.F.; Guo, L. Phenolic composition and antioxidant capacity of developing pear fruit from three cultivars. Food Sci. 2021, 42, 99–105. [Google Scholar]
- Savic, A.; Aradski, A.A.; Ivkovi, J.; Avikin, J.; Duleti-Lauev, S. Phenolic composition, and antioxidant and antineurodegenerative potential of methanolic extracts of fruit peel and flesh of pear varieties from Serbia. Pol. J. Food Nutr. Sci. 2021, 71, 225–236. [Google Scholar] [CrossRef]
- Twaij, B.M.; Hasan, M.N. Bioactive secondary metabolites from plant sources: Types, synthesis, and their therapeutic uses. Int. J. Plant Biol. 2022, 13, 4–14. [Google Scholar] [CrossRef]
- Pant, P.; Pandey, S.; Dall’Acqua, S. The influence of environmental conditions on secondary metabolites in medicinal plants: A literature review. Chem. Biodivers 2021, 18, e2100345. [Google Scholar] [CrossRef]
- War, A.R.; Buhroo, A.A.; Hussain, B.; Ahmad, T.; Nair, R.M.; Sharma, H.C. Plant defense and insect adaptation with reference to secondary metabolites. In Co-Evolution of Secondary Metabolites, 1st ed.; Mérillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2020; Volume 5, pp. 795–822. [Google Scholar]
- Franzoni, G.; Trivellini, A.; Bulgari, R.; Cocetta, G.; Ferrante, A. Bioactive molecules as regulatory signals in plant responses to abiotic stresses. In Plant Signaling Molecules, 1st ed.; Khan, M.I.R., Reddy, P.S., Ferrante, A., Khan, N.A., Eds.; Woodhead Publishing: London, UK, 2019; Volume 10, pp. 169–182. [Google Scholar]
- Sharma, A. Gene expression analysis in medicinal plants under abiotic stress conditions. In Plant Metabolites and Regulation under Environmental Stress, 1st ed.; Ahmad, P., Ahanger, M.A., Singh, V.P., Tripathi, D.K., Alam, P., Alyemeni, M.N., Eds.; Academic Press: Pittsburgh, PA, USA, 2018; Volume 23, pp. 407–414. [Google Scholar]
- Tang, C.; Guo, T.Y.; Zhang, Z.X.; Yang, P.; Song, H.L. Rapid visualized characterization of phenolic taste compounds in tea extract by high-performance thin-layer chromatography coupled to desorption electrospray ionization mass spectrometry. Food Chem. 2021, 355, 129555. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Q.; Fu, Y.Q.; Chen, J.X.; Wang, F.; Feng, Z.H.; Yin, J.F.; Zeng, L.; Xu, Y.Q. Effects of baking treatment on the sensory quality and physicochemical properties of green tea with different processing methods. Food Chem. 2022, 380, 132217. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Yan, S.J.; Feng, Z.S.; Xiao, L.X.; Hu, X.S. Changes in the volatile compounds and chemical and physical properties of Yali pear (Pyrus bertschneideri Reld) during storage. Food Chem. 2006, 97, 248–255. [Google Scholar] [CrossRef]
- Imran, M.; Salehi, B.; Sharifi-Rad, J.; Gondal, T.A.; Saeed, F.; Imran, A.; Shahbaz, M.; Fokou, P.V.T.; Arshad, M.U.; Khan, H.; et al. Kaempferol: A key emphasis to its anticancer potential. Molecules 2019, 24, 2277. [Google Scholar] [CrossRef] [PubMed]
- Na, W.; Kang, M.K.; Park, S.H.; Kim, D.Y.; Oh, S.Y.; Oh, M.S.; Park, S.; Kang, I.J.; Kang, Y.H. Aesculetin accelerates osteoblast differentiation and matrix-vesicle-mediated mineralization. Int. J. Mol. Sci. 2021, 22, 12391. [Google Scholar] [CrossRef]
- Chen, Y.Z.; Wang, S.R.; Li, T.; Zhang, G.C.; Yang, J. Antifungal activity of 6-Methylcoumarin against Valsa mali and its possible mechanism of action. J. Fungi 2022, 9, 5. [Google Scholar] [CrossRef]
- Pandey, P.; Khan, F. A mechanistic review of the anticancer potential of hesperidin, a natural flavonoid from citrus fruits. Nutr. Res. 2021, 92, 21–31. [Google Scholar] [CrossRef]
- Tejada, S.; Tejada, S.; Pinya, S.; Martorell, M.; Capó, X.; Tur, J.A.; Pons, A.; Sureda, A. Potential anti-inflammatory effects of hesperidin from the genus citrus. Curr. Med. Chem. 2018, 25, 4929–4945. [Google Scholar] [CrossRef]
- Vendramin, V.; Viel, A.; Vincenzi, S. Caftaric acid isolation from unripe grape: A ‘green’ alternative for hydroxycinnamic acids recovery. Molecules 2021, 26, 1148. [Google Scholar] [CrossRef]
- Memariani, Z.; Abbas, S.Q.; Hassan, S.S.; Ahmadi, A.; Chabra, A. Naringin and naringeninin as anticancer agents and adjuvants in cancer combination therapy: Effcacy and molecular mechanisms of action, a comprehensive narrative review. Pharmacol. Res. 2021, 171, 105264. [Google Scholar] [CrossRef]
- Escarpa, A.; Gonzalez, M.C. Evaluation of high-performance liquid chromatography for determination of phenolic compounds in pear horticultural cultivars. Chromatographia 2000, 51, 37–43. [Google Scholar] [CrossRef]
- Salta, J.; Martins, A.; Santos, R.G.; Neng, N.R.; Nogueira, J.M.F.; Justino, J.; Rauter, A.P. Phenolics composition and antioxidant activity of Rocha pear and other pear cultivars—A comparative study. J. Funct. Foods 2010, 2, 153–157. [Google Scholar] [CrossRef]
- Yuan, J.; Zhang, S.L.; Cao, Y.F.; Wu, J.; Tian, L.M.; Tao, S.T.; Dong, X.G. Polyphenolic compound and substances determination of enzymatic browning in pear. Acta Hortic. Sin. 2011, 38, 7–14. [Google Scholar]
- Zeng, S.M.; Yang, J.; Wang, L.; Wang, S.K.; Li, X.G. Study on phenolics content and its antioxidant activity in the fruits of pear species. J. Fruit Sci. 2014, 31, 39–44. [Google Scholar]
- Colaric, M.; Stampar, F.; Hudina, M. Changes in sugars and phenolics concentrations of Williams pear leaves during the growing season. Can. J. Plant Sci. 2006, 86, 1203–1208. [Google Scholar] [CrossRef]
- Wu, F.; Pu, Y.f.; Lei, X.Y.; He, Z.H.; Shen, L.R.; Bai, H.J. Phenolic compounds and antioxidant activity of Kuerlexiangli pear fruit during the growth period. J. Fruit Sci. 2022, 39, 574–583. [Google Scholar]
- Azevedo, J.; Fernandes, I.; Faria, A.; Oliveira, J.; Fernandes, A.; Freitas, V.D.; Mateus, N. Antioxidant properties of anthocyanidins, anthocyanidin-3-glucosides and respective portisins. Food Chem. 2010, 119, 518–523. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, H.; Guan, Q.; Dong, R.; Ran, K.; Wang, H.; Dong, X.; Wei, S. Metabolomics Analysis of Phenolic Composition and Content in Five Pear Cultivars Leaves. Plants 2024, 13, 2513. https://doi.org/10.3390/plants13172513
Jiao H, Guan Q, Dong R, Ran K, Wang H, Dong X, Wei S. Metabolomics Analysis of Phenolic Composition and Content in Five Pear Cultivars Leaves. Plants. 2024; 13(17):2513. https://doi.org/10.3390/plants13172513
Chicago/Turabian StyleJiao, Huijun, Qiuzhu Guan, Ran Dong, Kun Ran, Hongwei Wang, Xiaochang Dong, and Shuwei Wei. 2024. "Metabolomics Analysis of Phenolic Composition and Content in Five Pear Cultivars Leaves" Plants 13, no. 17: 2513. https://doi.org/10.3390/plants13172513
APA StyleJiao, H., Guan, Q., Dong, R., Ran, K., Wang, H., Dong, X., & Wei, S. (2024). Metabolomics Analysis of Phenolic Composition and Content in Five Pear Cultivars Leaves. Plants, 13(17), 2513. https://doi.org/10.3390/plants13172513