

Aminoethoxyvinylglicine and 1-Methylcyclopropene: Effects on Preharvest Drop, Fruit Maturity, Quality, and Associated Gene Expression of ‘Honeycrisp’ Apples in the US Mid-Atlantic

Abstract
:1. Introduction
2. Results
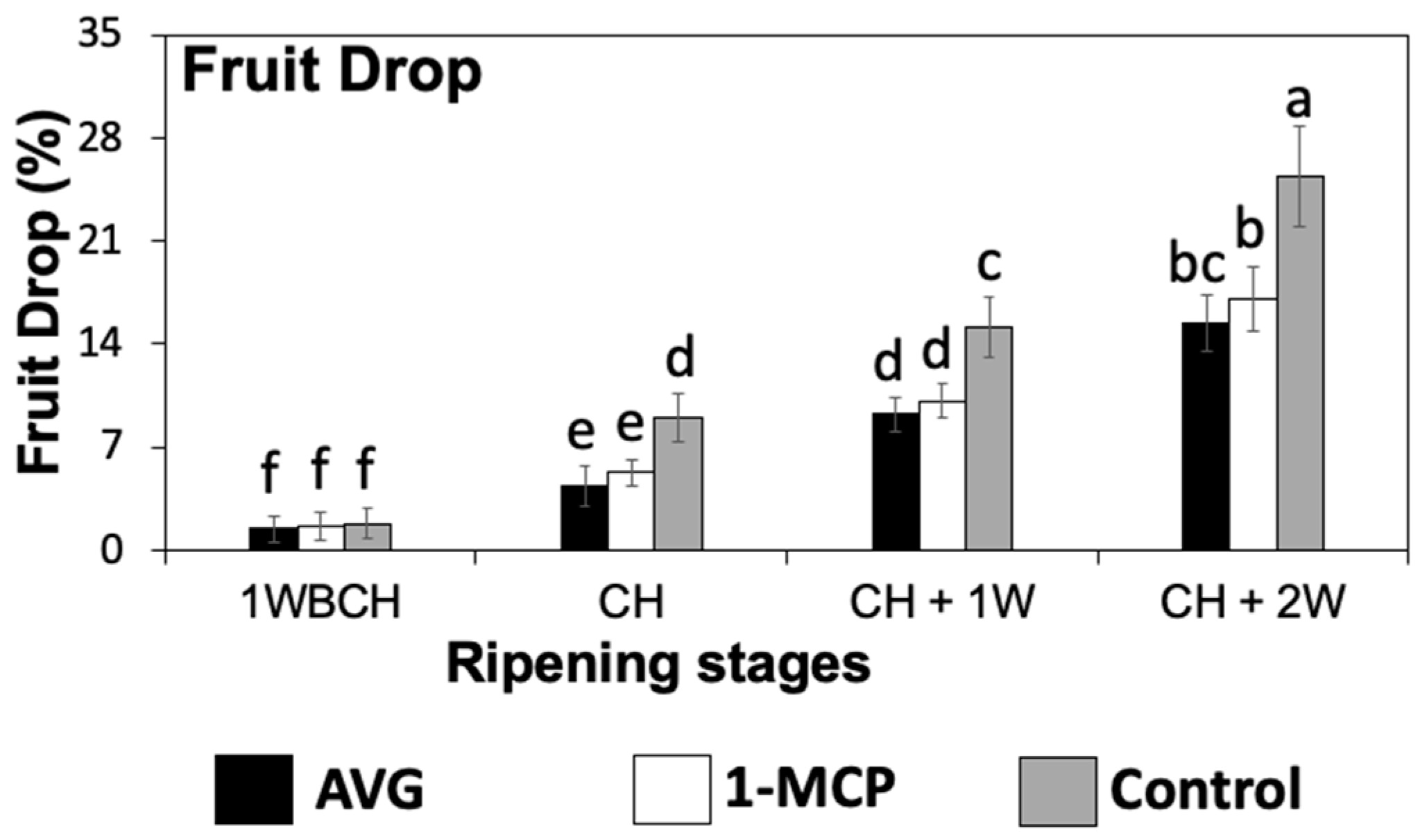
2.1. AVG and 1-MCP Effects on ‘Honeycrisp’ Fruit Drop during On-the-Tree Ripening
2.2. AVG and 1-MCP Effects on Ethylene Production and Physicochemical Parameters of ‘Honeycrisp’ during On-the-Tree Ripening and AVG and 1-MCP Effects on ‘Honeycrisp’ Fruit Drop during On-the-Tree Ripening
2.3. AVG and 1-MCP Effects on ‘Honeycrisp’ Skin Color during On-the-Tree Ripening
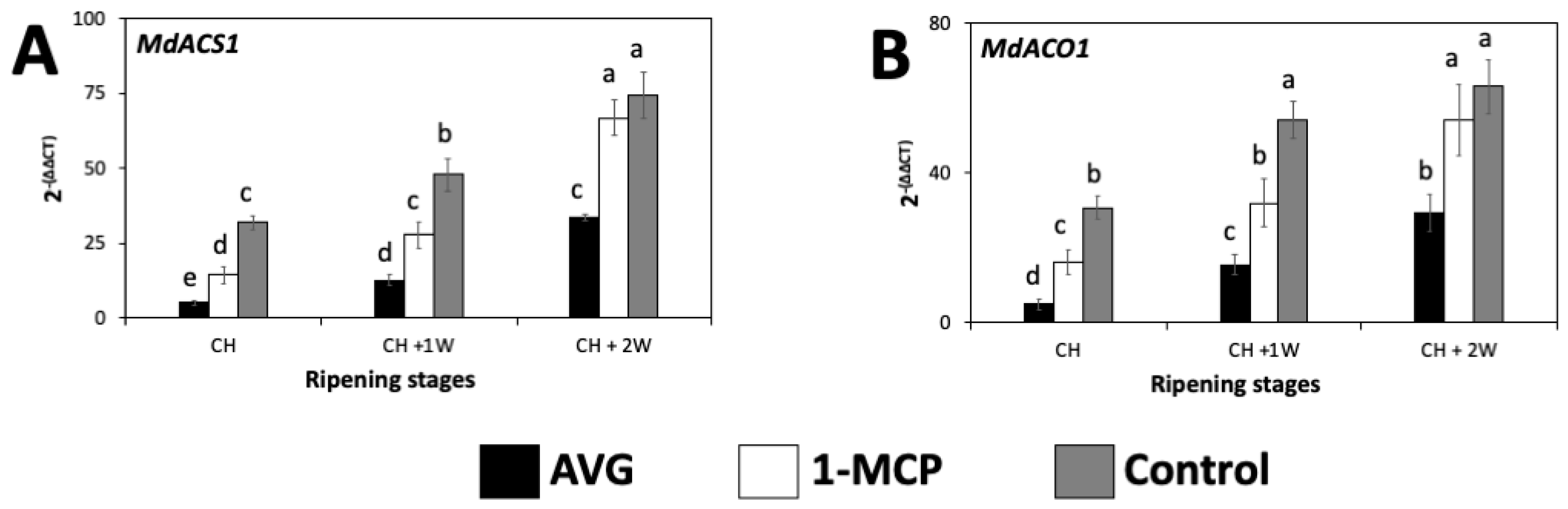
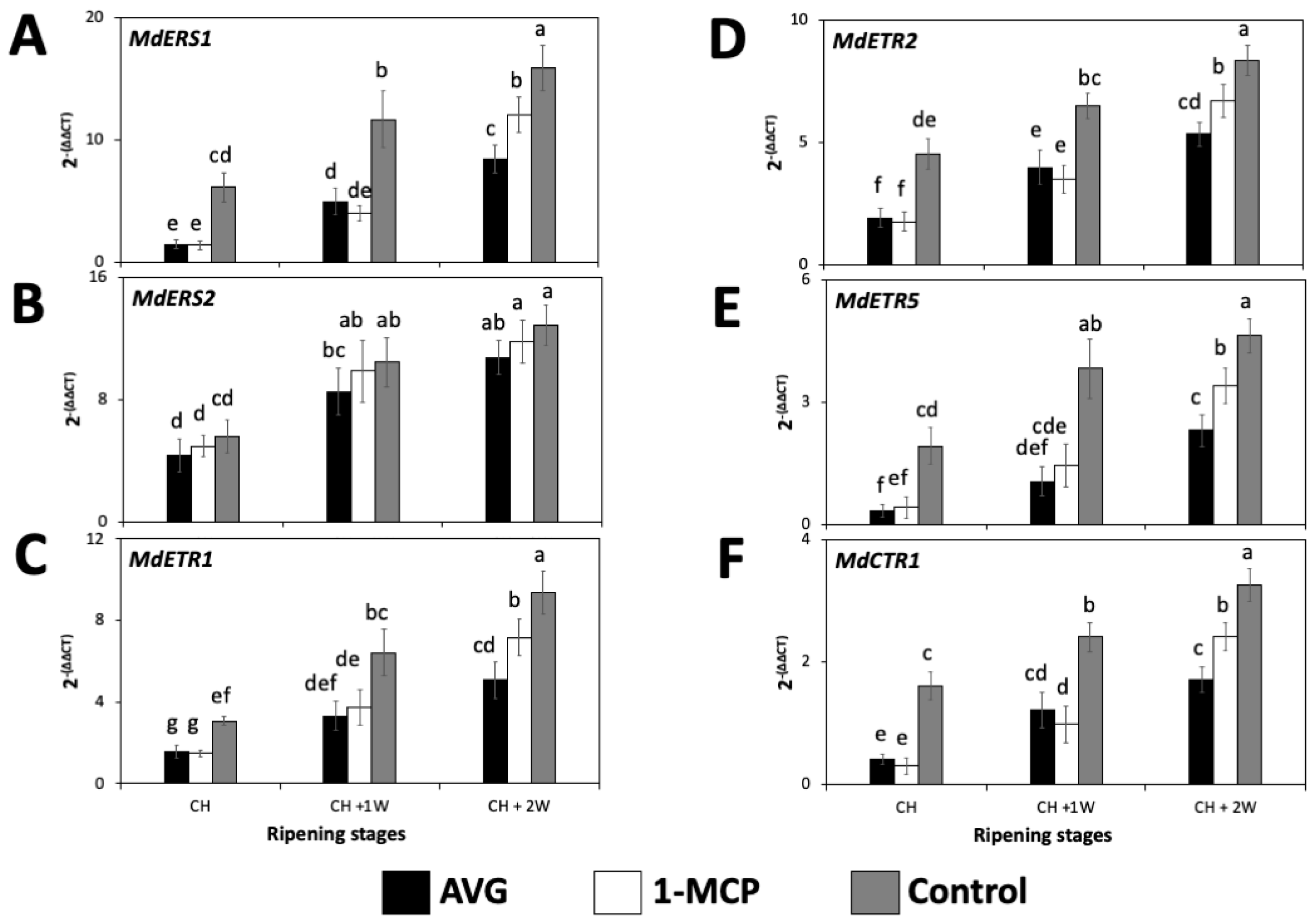
2.4. AVG and 1-MCP Effects on the Expression of Ethylene-Related Genes in ‘Honeycrisp’ Fruit Flesh during On-the-Tree Ripening
2.4.1. Ethylene-Biosynthesis-Related Genes
2.4.2. Ethylene-Perception-Related Genes
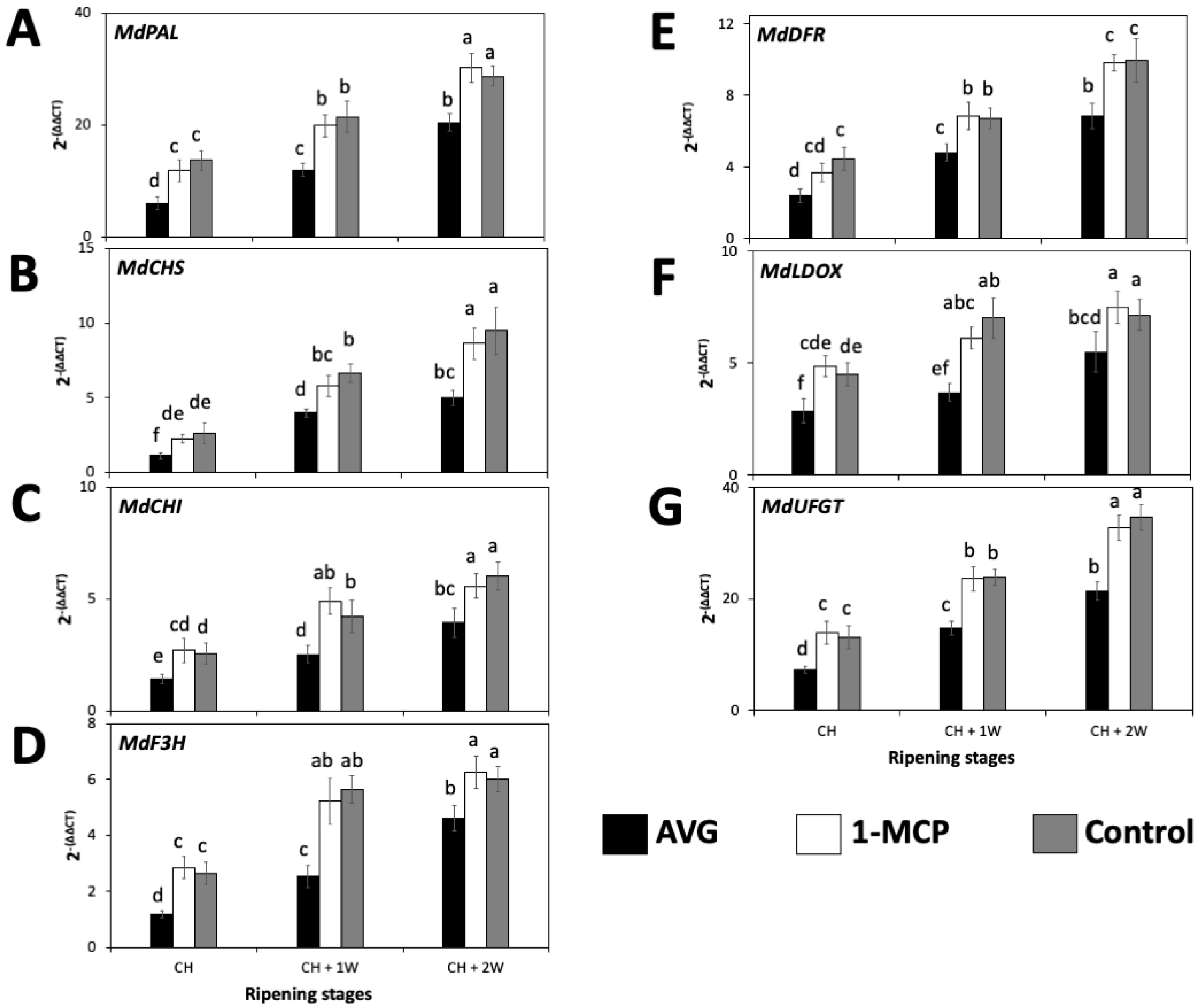
2.5. AVG and 1-MCP Effects on the Expression of Anthocyanin-Biosynthesis-Related Genes in ‘Honeycrisp’ Fruit Skin during On-the-Tree Ripening
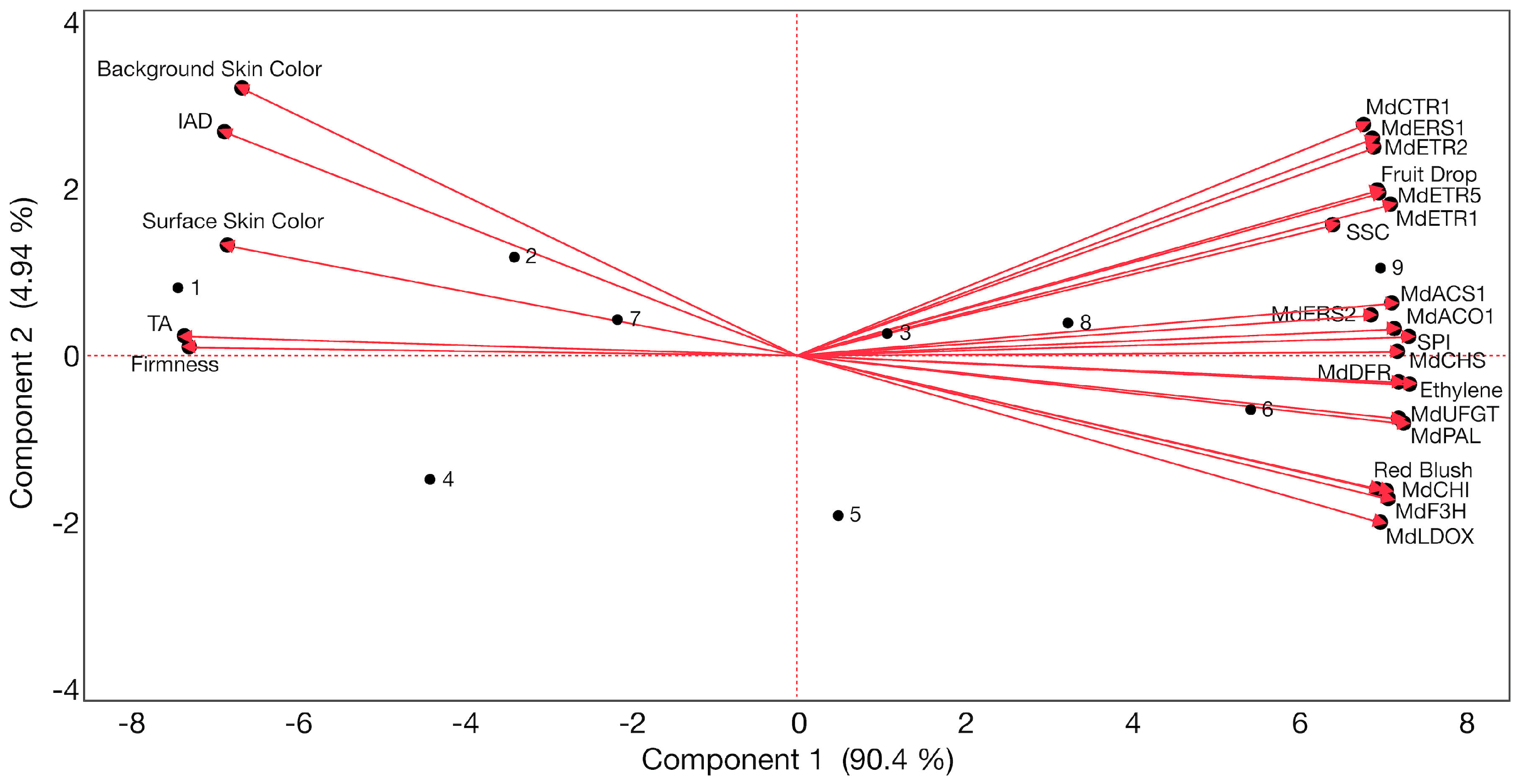
2.6. Associations between Fruit Drop, Ethylene Production, Physicochemical Properties, Skin Color, and Expression of Key Anthocyanin- and Ethylene-Related Genes in ‘Honeycrisp’ Fruit
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. Fruit Drop Assessments
4.3. Ethylene Production
4.4. Fruit Physicochemical and Skin Color Evaluations
4.5. Real-Time Quantitative RT-PCR Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schupp, J.R.; Greene, D.W. Effect of Aminoethoxyvinylglycine (AVG) on Preharvest Drop, Fruit Quality, and Maturation of ‘McIntosh’ Apples. I. Concentration and Timing of Dilute Applications of AVG. HortScience 2004, 39, 1030–1035. [Google Scholar] [CrossRef]
- Arseneault, M.H.; Cline, J.A. AVG, NAA, Boron, and Magnesium Influence Preharvest Fruit Drop and Fruit Quality of ‘Honeycrisp’ Apples. Can. J. Plant Sci. 2018, 98, 741–752. [Google Scholar] [CrossRef]
- Arseneault, M.H.; Cline, J.A. A Review of Apple Preharvest Fruit Drop and Practices for Horticultural Management. Sci. Hortic. 2016, 211, 40–52. [Google Scholar] [CrossRef]
- Irish-Brown, A.; Schwallier, P.; Shane, B.; Tritten, B. Why Does Apple Fruit Drop Prematurely? Michigan State University Extension: East Lansing, MI, USA, 2011. [Google Scholar]
- Baugher, T.A.; Schupp, J.R. Relationship between ’Honeycrisp’ Crop Load and Sensory Panel Evaluations of the Fruit. J. Am. Pomol. Soc. 2010, 64, 226–233. [Google Scholar]
- Byers, R.E.; Eno, D.R. Harvest Date Influences Fruit Size and Yield of ‘York’ and ‘Golden Delicious’ Apple Trees. J. Tree Fruit Prod. 2002, 3, 63–79. [Google Scholar] [CrossRef]
- Burg, S.P.; Burg, E.A. Ethylene Action and the Ripening of Fruits: Ethylene Influences the Growth and Development of Plants and Is the Hormone Which Initiates Fruit Ripening. Science 1965, 148, 1190–1196. [Google Scholar] [CrossRef]
- Klee, H.J. Ethylene Signal Transduction. Moving beyond Arabidopsis. Plant Physiol. 2004, 135, 660–667. [Google Scholar] [CrossRef]
- Liu, J.; Islam, M.T.; Sherif, S.M. Effects of Aminoethoxyvinylglycine (AVG) and 1-Methylcyclopropene (1-MCP) on the Pre-Harvest Drop Rate, Fruit Quality, and Stem-End Splitting in ‘Gala’ Apples. Horticulturae 2022, 8, 1100. [Google Scholar] [CrossRef]
- Bonghi, C.; Tonutti, P.; Ramina, A. Biochemical and Molecular Aspects of Fruitlet Abscission. Plant Growth Regul. 2000, 31, 35–42. [Google Scholar] [CrossRef]
- Roberts, J.A.; Elliott, K.A.; Gonzalez-Carranza, Z.H. Abscission, Dehiscence, and Other Cell Separation Processes. Annu. Rev. Plant Biol. 2002, 53, 131–158. [Google Scholar] [CrossRef]
- Farcuh, M.; Rivero, R.M.; Sadka, A.; Blumwald, E. Ethylene Regulation of Sugar Metabolism in Climacteric and Non-Climacteric Plums. Postharvest Biol. Technol. 2018, 139, 20–30. [Google Scholar] [CrossRef]
- Costa, F.; Stella, S.; Van de Weg, W.E.; Guerra, W.; Cecchinel, M.; Dallavia, J.; Koller, B.; Sansavini, S. Role of the Genes Md-ACO1 and Md-ACS1 in Ethylene Production and Shelf Life of Apple (Malus domestica Borkh). Euphytica 2005, 141, 181–190. [Google Scholar] [CrossRef]
- Chu, C.L. Internal Ethylene Concentration of ‘McIntosh’, ‘Northern Spy’, ‘Empire’, ‘Mutsu’, and ‘Idared’ Apples during the Harvest Season. J. Am. Soc. Hortic. Sci. 1988, 113, 226–229. [Google Scholar] [CrossRef]
- Gussman, C.D.; Goffreda, J.C.; Gianfagna, T.J. Ethylene Production and Fruit-Softening Rates in Several Apple Fruit Ripening Variants. HortScience 1993, 28, 135–137. [Google Scholar] [CrossRef]
- Li, J.; Zhu, H.; Yuan, R. Profiling the Expression of Genes Related to Ethylene Biosynthesis, Ethylene Perception, and Cell Wall Degradation during Fruit Abscission and Fruit Ripening in Apple. J. Am. Soc. Hortic. Sci. 2010, 135, 391–401. [Google Scholar] [CrossRef]
- Sun, L.; John Bukovac, M.; Forsline, P.L.; van Nocker, S. Natural Variation in Fruit Abscission-Related Traits in Apple (Malus). Euphytica 2009, 165, 55–67. [Google Scholar] [CrossRef]
- Yuan, R.; Li, J. Effect of Sprayable 1-MCP, AVG, and NAA on Ethylene Biosynthesis, Preharvest Fruit Drop, Fruit Maturity, and Quality of ‘Delicious’ Apples. HortScience 2008, 43, 1454–1460. [Google Scholar] [CrossRef]
- Sisler, E.C.; Serek, M. Inhibitors of Ethylene Responses in Plants at the Receptor Level: Recent Developments. Physiol. Plant. 1997, 100, 577–582. [Google Scholar] [CrossRef]
- Lurie, S. Regulation of Ethylene Biosynthesis in Fruits by Aminoethoxyvinyl Glycine and 1-Methylcyclopropene. In Proceedings of the International Conference on Ripening Regulation and Postharvest Fruit Quality 796, Weingarten, Germany, 12–13 November 2007; pp. 31–41. [Google Scholar]
- Yang, S.F.; Hoffman, N.E. Ethylene Biosynthesis and Its Regulation in Higher Plants. Annu. Rev. Plant Physiol. 1984, 35, 155–189. [Google Scholar] [CrossRef]
- Cherian, S.; Figueroa, C.R.; Nair, H. ‘Movers and Shakers’ in the Regulation of Fruit Ripening: A Cross-Dissection of Climacteric versus Non-Climacteric Fruit. J. Exp. Bot. 2014, 65, 4705–4722. [Google Scholar] [CrossRef]
- Grierson, D. Ethylene and the Control of Fruit Ripening. In The Molecular Biology and Biochemistry of Fruit Ripening; Seymour, G.B., Poole, M., Giovannoni, J.J., Tucker, G.A., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 43–73. ISBN 9780813820392. [Google Scholar]
- Dal Cin, V.; Danesin, M.; Boschetti, A.; Dorigoni, A.; Ramina, A. Ethylene Biosynthesis and Perception in Apple Fruitlet Abscission (Malus domestica L. Borck). J. Exp. Bot. 2005, 56, 2995–3005. [Google Scholar]
- Dal Cin, V.; Rizzini, F.M.; Botton, A.; Tonutti, P. The Ethylene Biosynthetic and Signal Transduction Pathways Are Differently Affected by 1-MCP in Apple and Peach Fruit. Postharvest Biol. Technol. 2006, 42, 125–133. [Google Scholar] [CrossRef]
- Lee, S.A.; Ross, G.S.; Gardener, R.C. An Apple Homolog of the Ethylene Receptor Gene ETR1 (Accession No. AF 032448). Plant Physiol. 1998, 117, 1126. [Google Scholar]
- Wiersma, P.A.; Zhang, H.; Lu, C.; Quail, A.; Toivonen, P.M.A. Survey of the Expression of Genes for Ethylene Synthesis and Perception during Maturation and Ripening of ‘Sunrise’and ‘Golden Delicious’ Apple Fruit. Postharvest Biol. Technol. 2007, 44, 204–211. [Google Scholar] [CrossRef]
- Kieber, J.J.; Rothenberg, M.; Roman, G.; Feldmann, K.A.; Ecker, J.R. CTR1, a Negative Regulator of the Ethylene Response Pathway in Arabidopsis, Encodes a Member of the Raf Family of Protein Kinases. Cell 1993, 72, 427–441. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.L.; Larsen, P.B.; Wang, X.; Chang, C. Association of the Arabidopsis CTR1 Raf-like Kinase with the ETR1 and ERS Ethylene Receptors. Proc. Natl. Acad. Sci. USA 1998, 95, 5401–5406. [Google Scholar] [CrossRef] [PubMed]
- Miah, M.S.; Farcuh, M. The Expression of Key Ethylene and Anthocyanin Biosynthetic Genes of ‘Honeycrisp’ Apples Subjected to the Combined Use of Reflective Groundcovers and Aminoethoxyvinylglycine in the Mid-Atlantic US. Plants 2024, 13, 1141. [Google Scholar] [CrossRef]
- Whale, S.K.; Singh, Z.; Behboudian, M.H.; Janes, J.; Dhaliwal, S.S. Fruit Quality in ‘Cripp’s Pink’ Apple, Especially Colour, as Affected by Preharvest Sprays of Aminoethoxyvinylglycine and Ethephon. Sci. Hortic. 2008, 115, 342–351. [Google Scholar] [CrossRef]
- Watkins, C.B. The Use of 1-Methylcyclopropene (1-MCP) on Fruits and Vegetables. Biotechnol. Adv. 2006, 24, 389–409. [Google Scholar] [CrossRef]
- Yang, X.; Song, J.; Campbell-Palmer, L.; Fillmore, S.; Zhang, Z. Effect of Ethylene and 1-MCP on Expression of Genes Involved in Ethylene Biosynthesis and Perception during Ripening of Apple Fruit. Postharvest Biol. Technol. 2013, 78, 55–66. [Google Scholar] [CrossRef]
- Layne, D.R.; Jiang, Z.; Rushing, J.W. The Influence of Reflective Film and ReTain on Red Skin Coloration and Maturity of ‘Gala’ Apples. Horttechnology 2002, 12, 640–645. [Google Scholar] [CrossRef]
- Miah, M.S.; Farcuh, M. Combining the Use of Reflective Groundcovers and Aminoethoxyvinylglycine to Assess Effects on Skin Color, Preharvest Drop, and Quality of ‘Honeycrisp’ Apples in the Mid-Atlantic US. Horticulturae 2024, 10, 179. [Google Scholar] [CrossRef]
- Malladi, A.; Tonapi, K.V.; Kon, T.M. Aminoethoxyvinylglycine Reduces Preharvest Fruit Drop and Fruit Ethylene Evolution in ‘Red Delicious’ Apple but Affects Fruit Size and Quality Inconsistently. HortScience 2023, 58, 1410–1417. [Google Scholar] [CrossRef]
- Greene, D.W.; Schupp, J.R. Effect of Aminoethoxyvinylglycine (AVG) on Preharvest Drop, Fruit Quality, and Maturation of ‘McIntosh’ Apples. II. Effect of Timing and Concentration Relationships and Spray Volume. HortScience 2004, 39, 1036–1041. [Google Scholar] [CrossRef]
- Osorio, S.; Fernie, A.R. Biochemistry of Fruit Ripening. In The Molecular Biology and Biochemistry of Fruit Ripening; Seymour, G.B., Poole, M., Giovannoni, J.J., Tucker, G.A., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 1–19. [Google Scholar]
- Miah, M.S.; Hinson, C.; Farcuh, M. Assessing Fruit Maturity and Quality of ‘Buckeye Gala’Grown on a Diverse Panel of Apple (Malus domestica Borkh.) Rootstocks in Western Maryland. Agronomy 2023, 13, 2528. [Google Scholar] [CrossRef]
- Farcuh, M.; Copes, B.; Le-Navenec, G.; Marroquin, J.; Cantu, D.; Bradford, K.J.; Guinard, J.-X.; Van Deynze, A. Sensory, Physicochemical and Volatile Compound Analysis of Short and Long Shelf-Life Melon (Cucumis melo L.) Genotypes at Harvest and after Postharvest Storage. Food Chem. X 2020, 8, 100107. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.; Carbaugh, D.H. Effects of NAA, AVG, and 1-MCP on Ethylene Biosynthesis, Preharvest Fruit Drop, Fruit Maturity, and Quality of ‘Golden Supreme’ and ‘Golden Delicious’ Apples. HortScience 2007, 42, 101–105. [Google Scholar] [CrossRef]
- Scolaro, A.; Tomazini, M.; Argenta, L.C.; Amarante, C.; Petri, J.L.; Haweroth, F.J. Preharvest Control of ‘Royal Gala’ Apple Fruit Maturation by the Inhibition of Ethylene Action or Synthesis. Rev. Bras. Frutic. 2015, 37, 38–47. [Google Scholar] [CrossRef]
- Doerflinger, F.C.; Nock, J.F.; Miller, W.B.; Watkins, C.B. Preharvest Aminoethoxyvinylglycine (AVG) and 1-Methylcyclopropene (1-MCP) Effects on Ethylene and Starch Concentrations of ‘Empire’ and ‘McIntosh’ Apples. Sci. Hortic. 2019, 244, 134–140. [Google Scholar] [CrossRef]
- do Amarante, C.V.T.; Argenta, L.C.; de Freitas, S.T.; Steffens, C.A. Efficiency of Pre-Harvest Application of 1-MCP (HarvistaTM 1.3 SC) to Delay Maturation of ‘Cripps Pink’ Apple Fruit. Sci. Hortic. 2022, 293, 110715. [Google Scholar] [CrossRef]
- Argenta, L.C.; Scolaro, A.M.T.; Do Amarante, C.V.T.; Vieira, M.J.; Werner, S.S. Preharvest Treatment of’Gala’apples with 1-MCP and AVG–I: Effects on Fruit Maturation on the Tree. In Proceedings of the VIII International Postharvest Symposium: Enhancing Supply Chain and Consumer Benefits-Ethical and Technological Issues 1194, Cartagena, Spain, 21–24 June 2016; pp. 113–120. [Google Scholar]
- McArtney, S.J.; Obermiller, J.D.; Schupp, J.R.; Parker, M.L.; Edgington, T.B. Preharvest 1-Methylcyclopropene Delays Fruit Maturity and Reduces Softening and Superficial Scald of Apples during Long-Term Storage. HortScience 2008, 43, 366–371. [Google Scholar] [CrossRef]
- Doerflinger, F.C.; Al Shoffe, Y.; Sutanto, G.; Nock, J.F.; Watkins, C.B. Preharvest 1-Methylcyclopropene (1-MCP) Treatment Effects on Quality of Spot and Strip Picked ‘Gala’ Apples at Harvest and after Storage as Affected by Postharvest 1-MCP and Temperature Conditioning Treatments. Sci. Hortic. 2024, 325, 112682. [Google Scholar] [CrossRef]
- Tomala, K.; Grzęda, M.; Guzek, D.; Głąbska, D.; Gutkowska, K. Analysis of Possibility to Apply Preharvest 1-Methylcyclopropene (1-MCP) Treatment to Delay Harvesting of Red Jonaprince Apples. Sustainability 2020, 12, 4575. [Google Scholar] [CrossRef]
- Tomala, K.; Grzęda, M.; Guzek, D.; Głąbska, D.; Gutkowska, K. The Effects of Preharvest 1-Methylcyclopropene (1-MCP) Treatment on the Fruit Quality Parameters of Cold-Stored ‘Szampion’ Cultivar Apples. Agriculture 2020, 10, 80. [Google Scholar] [CrossRef]
- Kon, T.M.; Clavet, C.D. Enhancing Red Fruit Coloration of Apples in the Southeastern US with Reflective Fabrics. Horticulturae 2023, 9, 1125. [Google Scholar] [CrossRef]
- USDA Agricultural Marketing Service Apples Grades and Standards. Available online: https://www.ams.usda.gov/grades-standards/apple-grades-standards (accessed on 13 July 2024).
- Castañeda-Ovando, A.; de Lourdes Pacheco-Hernández, M.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical Studies of Anthocyanins: A Review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Feng, F.; Li, M.; Ma, F.; Cheng, L. Phenylpropanoid Metabolites and Expression of Key Genes Involved in Anthocyanin Biosynthesis in the Shaded Peel of Apple Fruit in Response to Sun Exposure. Plant Physiol. Biochem. 2013, 69, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Zheng, L.; He, S.; Zheng, Y.; Yi, S.; Deng, L. Anthocyanin Biosynthesis in Fruit Tree Crops: Genes and Their Regulation. Afr. J. Biotechnol. 2011, 10, 19890–19897. [Google Scholar]
- Ubi, B.E.; Honda, C.; Bessho, H.; Kondo, S.; Wada, M.; Kobayashi, S.; Moriguchi, T. Expression Analysis of Anthocyanin Biosynthetic Genes in Apple Skin: Effect of UV-B and Temperature. Plant Sci. 2006, 170, 571–578. [Google Scholar] [CrossRef]
- Honda, C.; Moriya, S. Anthocyanin Biosynthesis in Apple Fruit. Hortic. J. 2018, 87, 305–314. [Google Scholar] [CrossRef]
- Lancaster, J.E. Regulation of Skin Color in Apples. CRC Crit. Rev. Plant Sci. 1992, 10, 487–502. [Google Scholar] [CrossRef]
- Farcuh, M.; Tajima, H.; Lerno, L.A.; Blumwald, E. Changes in Ethylene and Sugar Metabolism Regulate Flavonoid Composition in Climacteric and Non-Climacteric Plums during Postharvest Storage. Food Chem. Mol. Sci. 2022, 4, 100075. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Dilley, D.R. Aminoethoxyvinylglycine, Combined with Ethephon, Can Enhance Red Color Development without over-Ripening Apples. HortScience 2001, 36, 328–331. [Google Scholar] [CrossRef]
- Blankenship, S.M.; Unrath, C.R. PAL and Ethylene Content during Maturation of Red and Golden Delicious Apples. Phytochemistry 1988, 27, 1001–1003. [Google Scholar] [CrossRef]
- Whale, S.K.; Singh, Z. Endogenous Ethylene and Color Development in the Skin of ‘Pink Lady’ Apple. J. Am. Soc. Hortic. Sci. 2007, 132, 20–28. [Google Scholar] [CrossRef]
- Byers, R.E.; Carbaugh, D.H.; Combs, L.D. Ethylene Inhibitors Delay Fruit Drop, Maturity, and Increase Fruit Size of “Arlet” Apples. HortScience 2005, 40, 2061–2065. [Google Scholar] [CrossRef]
- Sato, T.; Kudo, T.; Akada, T.; Wakasa, Y.; Niizeki, M.; Harada, T. Allelotype of a Ripening-Specific 1-Aminocyclopropane-1-Carboxylate Synthase Gene Defines the Rate of Fruit Drop in Apple. J. Am. Soc. Hortic. Sci. 2004, 129, 32–36. [Google Scholar] [CrossRef]
- Li, J.; Yuan, R. NAA and Ethylene Regulate Expression of Genes Related to Ethylene Biosynthesis, Perception, and Cell Wall Degradation during Fruit Abscission and Ripening in ‘Delicious’ Apples. J. Plant Growth Regul. 2008, 27, 283–295. [Google Scholar] [CrossRef]
- McMurchie, E.J.; McGlasson, W.B.; Eaks, I.L. Treatment of Fruit with Propylene Gives Information about the Biogenesis of Ethylene. Nature 1972, 237, 235–236. [Google Scholar] [CrossRef]
- Lelièvre, J.; Latchè, A.; Jones, B.; Bouzayen, M.; Pech, J. Ethylene and Fruit Ripening. Physiol. Plant. 1997, 101, 727–739. [Google Scholar] [CrossRef]
- Harada, T.; Sunako, T.; Wakasa, Y.; Soejima, J.; Satoh, T.; Niizeki, M. An Allele of the 1-Aminocyclopropane-1-Carboxylate Synthase Gene (Md-ACS1) Accounts for the Low Level of Ethylene Production in Climacteric Fruits of Some Apple Cultivars. Theor. Appl. Genet. 2000, 101, 742–746. [Google Scholar] [CrossRef]
- Varanasi, V.; Shin, S.; Johnson, F.; Mattheis, J.P.; Zhu, Y. Differential Suppression of Ethylene Biosynthesis and Receptor Genes in ‘Golden Delicious’ Apple by Preharvest and Postharvest 1-MCP Treatments. J. Plant Growth Regul. 2013, 32, 585–595. [Google Scholar] [CrossRef]
- Costa, F.; Alba, R.; Schouten, H.; Soglio, V.; Gianfranceschi, L.; Serra, S.; Musacchi, S.; Sansavini, S.; Costa, G.; Fei, Z. Use of Homologous and Heterologous Gene Expression Profiling Tools to Characterize Transcription Dynamics during Apple Fruit Maturation and Ripening. BMC Plant Biol. 2010, 10, 229. [Google Scholar] [CrossRef] [PubMed]
- Dias, C.; Ribeiro, T.; Rodrigues, A.C.; Ferrante, A.; Vasconcelos, M.W.; Pintado, M. Improving the Ripening Process after 1-MCP Application: Implications and Strategies. Trends Food Sci. Technol. 2021, 113, 382–396. [Google Scholar] [CrossRef]
- Tieman, D.M.; Taylor, M.G.; Ciardi, J.A.; Klee, H.J. The Tomato Ethylene Receptors NR and LeETR4 Are Negative Regulators of Ethylene Response and Exhibit Functional Compensation within a Multigene Family. Proc. Natl. Acad. Sci. USA 2000, 97, 5663–5668. [Google Scholar] [CrossRef]
- Kevany, B.M.; Tieman, D.M.; Taylor, M.G.; Cin, V.D.; Klee, H.J. Ethylene Receptor Degradation Controls the Timing of Ripening in Tomato Fruit. Plant J. 2007, 51, 458–467. [Google Scholar] [CrossRef]
- Kieber, J.J. The Ethylene Signal Transduction Pathway in Arabidopsis. J. Exp. Bot. 1997, 48, 211–218. [Google Scholar] [CrossRef]
- Adams-Phillips, L.; Barry, C.; Kannan, P.; Leclercq, J.; Bouzayen, M.; Giovannoni, J. Evidence That CTR1-Mediated Ethylene Signal Transduction in Tomato Is Encoded by a Multigene Family Whose Members Display Distinct Regulatory Features. Plant Mol. Biol. 2004, 54, 387–404. [Google Scholar] [CrossRef]
- Farcuh, M.; Toubiana, D.; Sade, N.; Rivero, R.M.; Doron-Faigenboim, A.; Nambara, E.; Sadka, A.; Blumwald, E. Hormone Balance in a Climacteric Plum Fruit and Its Non-Climacteric Bud Mutant during Ripening. Plant Sci. 2019, 280, 51–65. [Google Scholar] [CrossRef]
- El-Sharkawy, I.; Kim, W.S.; El-Kereamy, A.; Jayasankar, S.; Svircev, A.M.; Brown, D.C.W. Isolation and Characterization of Four Ethylene Signal Transduction Elements in Plums (Prunus salicina L.). J. Exp. Bot. 2007, 58, 3631–3643. [Google Scholar] [CrossRef]
- El-Sharkawy, I.; Jones, B.; Li, Z.G.; Lelièvre, J.-M.; Pech, J.-C.; Latché, A. Isolation and Characterization of Four Ethylene Perception Elements and Their Expression during Ripening in Pears (Pyrus communis L.) with/without Cold Requirement. J. Exp. Bot. 2003, 54, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Klee, H.J. Control of Ethylene-mediated Processes in Tomato at the Level of Receptors. J. Exp. Bot. 2002, 53, 2057–2063. [Google Scholar] [CrossRef]
- Greene, D.W. Preharvest Drop Control and Maturity of ‘Delicious’ Apples as Affected by Aminoethoxyvinylglycine (AVG). J. Tree Fruit Prod. 2002, 3, 1–10. [Google Scholar] [CrossRef]
- Chen, Z.; Yu, L.; Liu, W.; Zhang, J.; Wang, N.; Chen, X. Research Progress of Fruit Color Development in Apple (Malus domestica Borkh.). Plant Physiol. Biochem. 2021, 162, 267–279. [Google Scholar] [CrossRef]
- Shafiq, M.; Singh, Z.; Khan, A.S. Pre-Harvest Ethephon Application and Training Systems Affect Colour Development, Accumulation of Flavonoids and Fruit Quality of ’Cripps Pink’ apple. Aust. J. Crop Sci. 2014, 8, 1579–1589. [Google Scholar]
- Faragher, J.D.; Brohier, R.L. Anthocyanin Accumulation in Apple Skin during Ripening: Regulation by Ethylene and Phenylalanine Ammonia-Lyase. Sci. Hortic. 1984, 22, 89–96. [Google Scholar] [CrossRef]
- Yu, J.; Qiu, K.; Sun, W.; Yang, T.; Wu, T.; Song, T.; Zhang, J.; Yao, Y.; Tian, J. A Long Noncoding RNA Functions in High-Light-Induced Anthocyanin Accumulation in Apple by Activating Ethylene Synthesis. Plant Physiol. 2022, 189, 66–83. [Google Scholar] [CrossRef] [PubMed]
- An, J.-P.; Wang, X.-F.; Li, Y.-Y.; Song, L.-Q.; Zhao, L.-L.; You, C.-X.; Hao, Y.-J. EIN3-LIKE1, MYB1, and ETHYLENE RESPONSE FACTOR3 Act in a Regulatory Loop That Synergistically Modulates Ethylene Biosynthesis and Anthocyanin Accumulation. Plant Physiol. 2018, 178, 808–823. [Google Scholar] [CrossRef]
- Awad, M.A.; De Jager, A. Formation of Flavonoids, Especially Anthocyanin and Chlorogenic Acid in ‘Jonagold’ Apple Skin: Influences of Growth Regulators and Fruit Maturity. Sci. Hortic. 2002, 93, 257–266. [Google Scholar] [CrossRef]
- Farcuh, M.; Hopfer, H. Aroma Volatiles as Predictors of Chilling Injury Development during Peach (Prunus persica (L.) Batsch) Cold Storage and Subsequent Shelf-Life. Postharvest Biol. Technol. 2023, 195, 112137. [Google Scholar] [CrossRef]
- Kim, H.; Farcuh, M.; Cohen, Y.; Crisosto, C.; Sadka, A.; Blumwald, E. Non-Climacteric Ripening and Sorbitol Homeostasis in Plum Fruits. Plant Sci. 2015, 231, 30–39. [Google Scholar] [CrossRef]
- Tong, C.B.S.; Bedford, D.S.; Luby, J.J.; Propsom, F.M.; Beaudry, R.M.; Mattheis, J.P.; Watkins, C.B.; Weis, S.A. Location and Temperature Effects on Soft Scald in ‘Honeycrisp’ Apples. HortScience 2003, 38, 1153–1155. [Google Scholar] [CrossRef]
- Infante, R.; Farcuh, M.; Meneses, C. Monitoring the Sensorial Quality and Aroma through an Electronic Nose in Peaches during Cold Storage. J. Sci. Food Agric. 2008, 88, 2073–2078. [Google Scholar] [CrossRef]
- Ziosi, V.; Noferini, M.; Fiori, G.; Tadiello, A.; Trainotti, L.; Casadoro, G.; Costa, G. A New Index Based on Vis Spectroscopy to Characterize the Progression of Ripening in Peach Fruit. Postharvest Biol. Technol. 2008, 49, 319–329. [Google Scholar] [CrossRef]
- Blanpied, G.D.; Silsby, J. Cornell Cooperative Extension; Cornell Cooperative Extension: Ithaca, NY, USA, 1992; pp. 7–9. [Google Scholar]
- Chang, S.; Puryear, J.; Cairney, J. A Simple and Efficient Method for Isolating RNA from Pine Trees. Plant Mol. Biol. Report. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Kim, H.; Saha, P.; Farcuh, M.; Li, B.; Sadka, A.; Blumwald, E. RNA-Seq Analysis of Spatiotemporal Gene Expression Patterns during Fruit Development Revealed Reference Genes for Transcript Normalization in Plums. Plant Mol. Biol. Report. 2015, 33, 1634–1649. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Ethylene (μL C2H4 kg−1 h−1) | Firmness (N) | SPI (1 to 8) | SSC (%) | TA (% Malic Acid) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CH | CH + 1W | CH + 2W | CH | CH + 1W | CH + 2W | CH | CH + 1W | CH + 2W | CH | CH + 1W | CH + 2W | CH | CH + 1W | CH + 2W | |
| AVG | 1.9 ± 0.5 f | 11.5 ± 0.8 e | 28.0 ± 1.9 c | 79.9 ± 1.4 a | 73.0 ± 1.1 b | 63.8 ± 1.5 c | 3.2 ± 0.1 f | 4.8 ± 0.2 d | 6.5 ± 0.4 b | 12.7 ± 0.3 d | 13.7 ± 0.2 ab | 13.8 ± 0.3 a | 0.53 ± 0.03 a | 0.48 ± 0.01 b | 0.39 ± 0.02 e |
| 1-MCP | 9.9 ± 1.8 e | 24.3 ± 2.4 cd | 36.6 ± 3.5 ab | 74.4 ± 1.7 b | 66.1 ± 1.1 c | 58.0 ± 0.9 d | 3.8 ± 0.2 e | 5.8 ± 0.3 c | 7.6 ± 0.4 a | 12.9 ± 0.2 cd | 13.6 ± 0.2 ab | 13.8 ± 0.2 a | 0.49 ± 0.01 b | 0.42 ± 0.02 d | 0.33 ± 0.02 f |
| Control | 16.7 ± 2.0 d | 32.1 ± 1.6 b | 39.1 ± 3.2 a | 67.1 ± 1.3 c | 59.7 ± 1.7 d | 57.2 ± 1.5 d | 4.6 ± 0.2 d | 6.8 ± 0.4 b | 7.8 ± 0.5 a | 13.3 ± 0.4 bc | 13.9 ± 0.2 a | 14.1 ± 0.3 a | 0.45 ± 0.02 cd | 0.38 ± 0.01 e | 0.33 ± 0.01 f |
| Treatment | Surface Skin Color (Hue°) | Skin Blush (%) | Background Skin Color (Hue°) | Index of Absorbance Difference (IAD) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CH | CH + 1W | CH + 2W | CH | CH + 1W | CH + 2W | CH | CH + 1W | CH + 2W | CH | CH + 1W | CH + 2W | |
| AVG | 59.3 ± 2.6 a | 48.9 ± 2.5 bc | 47.3 ± 1.9 bc | 35 ± 1.8 d | 40 ± 2.9 d | 49 ± 2.3 b | 112.3 ± 3.7 a | 110.2 ± 3.3 a | 103.1 ± 2.8 bc | 0.9 ± 0.04 a | 0.8 ± 0.05 ab | 0.6 ± 0.01 c |
| 1-MCP | 50.3 ± 2.6 b | 41.7 ± 1.4 de | 39.5 ± 1.0 de | 43 ± 2.0 c | 50 ± 1.9 b | 58 ± 3.0 a | 104.4 ± 2.7 bc | 98.7 ± 2.9 de | 96.4 ± 2.5 e | 0.6 ± 0.03 c | 0.5 ± 0.02 de | 0.4 ± 0.01 ef |
| Control | 46.8 ± 1.8 bc | 38.5 ± 1.2 e | 39.1 ± 0.8 e | 44 ± 2.2 c | 51 ± 2.4 b | 60 ± 3.1 a | 105.6 ± 3.1 b | 100.9 ± 2.2 cd | 95.1 ± 2.3 e | 0.7 ± 0.03 bc | 0.5 ± 0.01 de | 0.4 ± 0.02 ef |
| Gene Name | Description | Primer Orientation | Primer Sequence (5′ to 3′) |
|---|---|---|---|
| MdPAL | Phenylalanine ammonia-lyase | Forward Reverse | GTGCTGTGGAGTCCCCGCTT GGTGAGGCTCTCTCCGCCAAGT |
| MdCHS | Chalcone synthase | Forward | GGAGACAACTGGAGAAGGACTGGAA |
| Reverse | CGACATTGATACTGGTGTCTTCA | ||
| MdCHI | Chalcone isomerase | Forward | GGGATAACCTCGCGGCCAAA |
| Reverse | GCATCCATGCCGGAAGCTACAA | ||
| MdF3H | Flavanone 3-hydroxylase | Forward | TGGAAGCTTGTGAGGACTGGGGT |
| Reverse | CTCCTCCGATGGCAAATCAAAGA | ||
| MdDFR | Dihydroflavonol 4-reductase | Forward | GATAGGGTTTGAGTTCAAGTA |
| Reverse | TCTCCTCAGCAGCCTCAGTTTTCT | ||
| MdLDOX | Leucoanthocyanidin dioxygenase | Forward | CCAAGTGAAGCGGGTTGTGCT |
| Reverse | CAAAGCAGGCGGACAGGAGTAGC | ||
| MdUFGT | UDP glucose-flavonoid 3- o -glucosyl transferase | Forward | CCACCGCCCTTCCAAACACTCT |
| Reverse | CCACCGCCCTTCCAAACACTCT | ||
| MdACS1 | 1-aminocyclopropane-carboxylase (ACC) synthase | Forward | CTCCTCCTTTCCTTCGTTGA |
| Reverse | ACCATGTCGTCGTTGGAGTAG | ||
| MdACO1 | ACC oxidase | Forward | ATCAATGATGCTTGTGAGAACTG |
| Reverse | GGTCTTCTTGTAGTGATCCTTGG | ||
| MdERS1 | Ethylene-response sensor | Forward | TCCAGAACTGGTATGAACCTACA |
| Reverse | AGAACTGTTGAAGACTTCGTTGA | ||
| MdERS2 | Ethylene-response sensor | Forward | TGCGAAACCAGAATCTTCAAGA |
| Reverse | CCTCAGTTGACGCTGGATAAAA | ||
| MdETR1 | Ethylene receptor-type | Forward | GCACCTAGGATGTGATGTAACAG |
| Reverse | TCATGTATACGGACAGCAAGTTC | ||
| MdETR2 | Ethylene receptor-type | Forward | AGGCAAACAAAGGGATGACA |
| Reverse | AGGCAAACAAAGGGATGACA | ||
| MdETR5 | Ethylene receptor-type | Forward | GTTCTTCCGGTTGCAGATTC |
| Reverse | ATGCATTGGCCTTCTCATTC | ||
| MdCTR1 | Constitutive triple response | Forward | ACAAGATTTTCATGCCGAAC |
| Reverse | TATGGACAAGTTTGGAGGCT | ||
| MdACT | Actin | Forward Reverse | TGACCGAATGAGCAAGGAAATTACT TACTCAGCTTTGGCAATCCACATC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, E.; Farcuh, M. Aminoethoxyvinylglicine and 1-Methylcyclopropene: Effects on Preharvest Drop, Fruit Maturity, Quality, and Associated Gene Expression of ‘Honeycrisp’ Apples in the US Mid-Atlantic. Plants 2024, 13, 2524. https://doi.org/10.3390/plants13172524
Johnson E, Farcuh M. Aminoethoxyvinylglicine and 1-Methylcyclopropene: Effects on Preharvest Drop, Fruit Maturity, Quality, and Associated Gene Expression of ‘Honeycrisp’ Apples in the US Mid-Atlantic. Plants. 2024; 13(17):2524. https://doi.org/10.3390/plants13172524
Chicago/Turabian StyleJohnson, Emily, and Macarena Farcuh. 2024. "Aminoethoxyvinylglicine and 1-Methylcyclopropene: Effects on Preharvest Drop, Fruit Maturity, Quality, and Associated Gene Expression of ‘Honeycrisp’ Apples in the US Mid-Atlantic" Plants 13, no. 17: 2524. https://doi.org/10.3390/plants13172524