
Genome-Wide Characterization and Expression Analysis of CsPALs in Cucumber (Cucumis sativus L.) Reveal Their Potential Roles in Abiotic Stress and Aphid Stress Tolerance

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
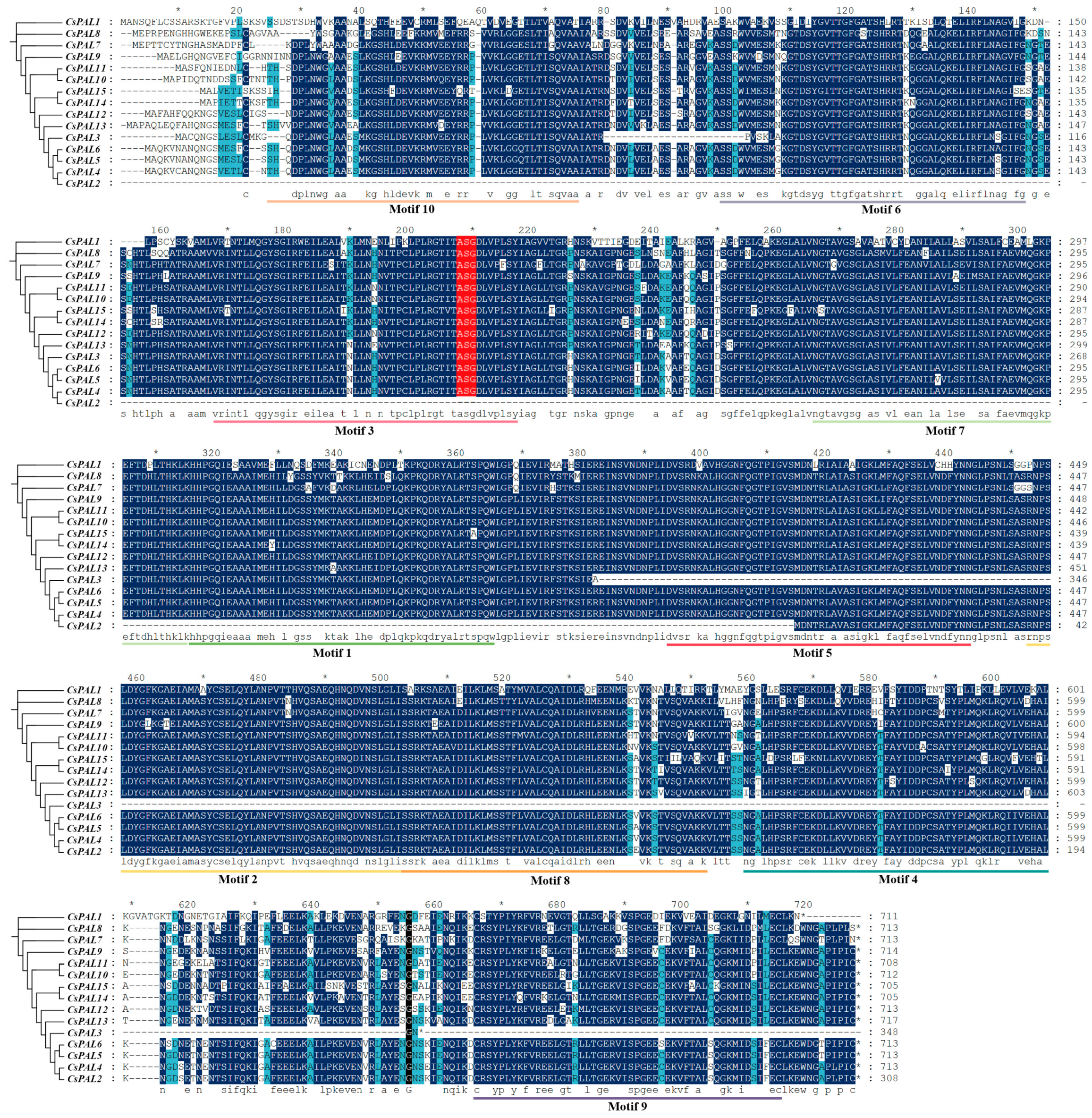
2.1. Identification of PAL Genes in Cucumber
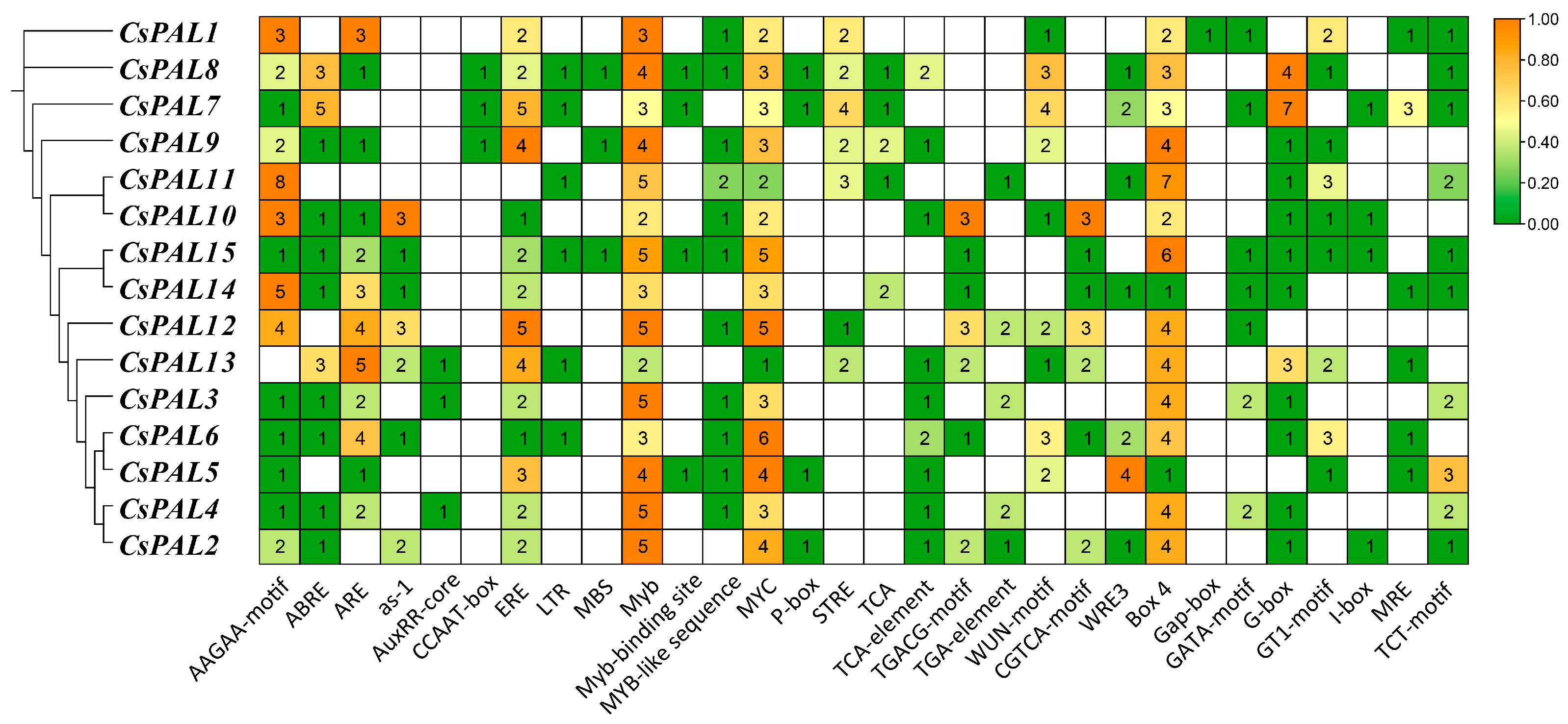
2.2. Analysis of Cis-Regulatory Elements in CsPAL
2.3. Chromosomal Location, Tandem Duplication, and Ka/Ks Ratio of CsPAL Genes
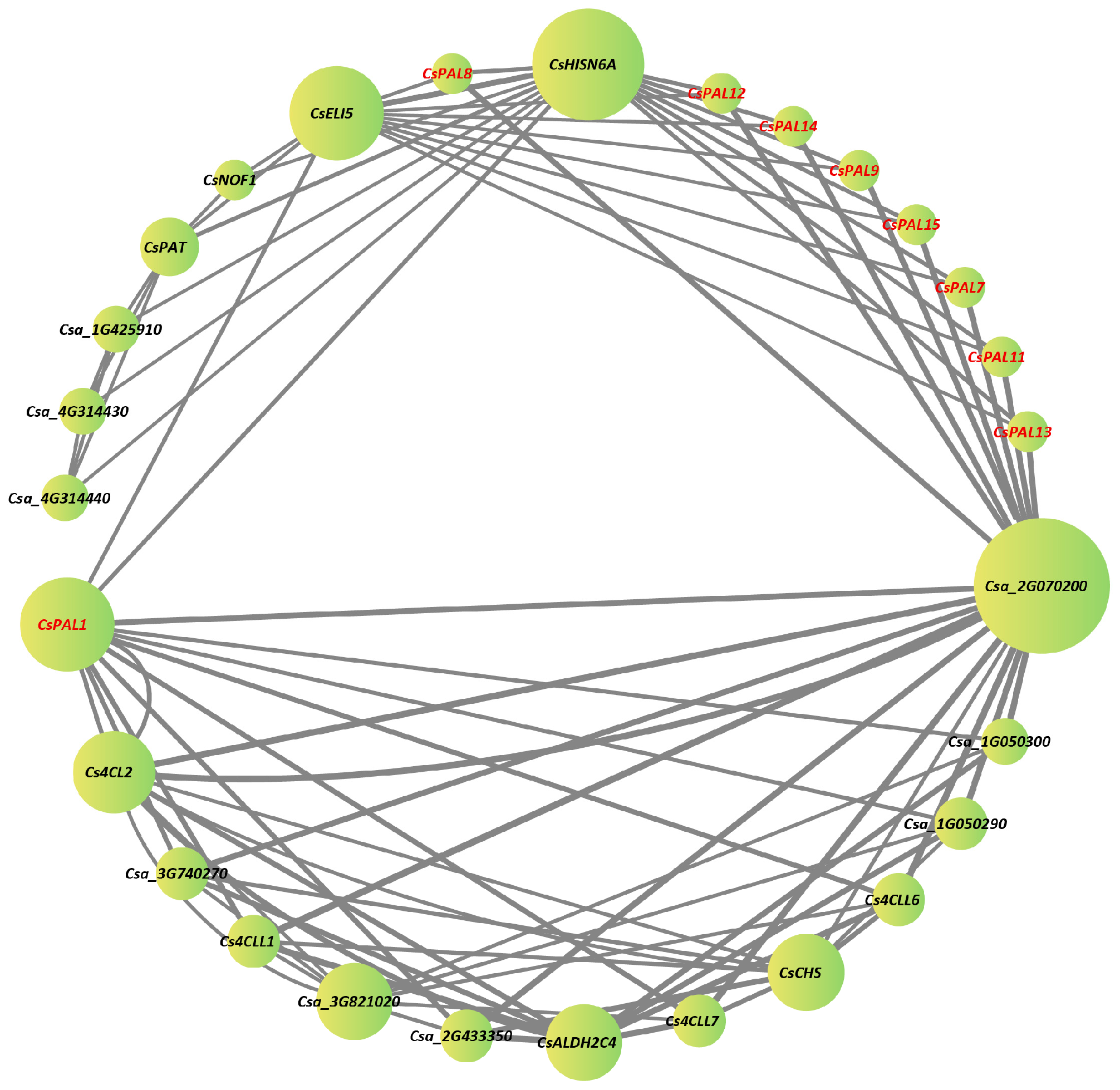
2.4. Protein-Protein Interaction Network Analysis
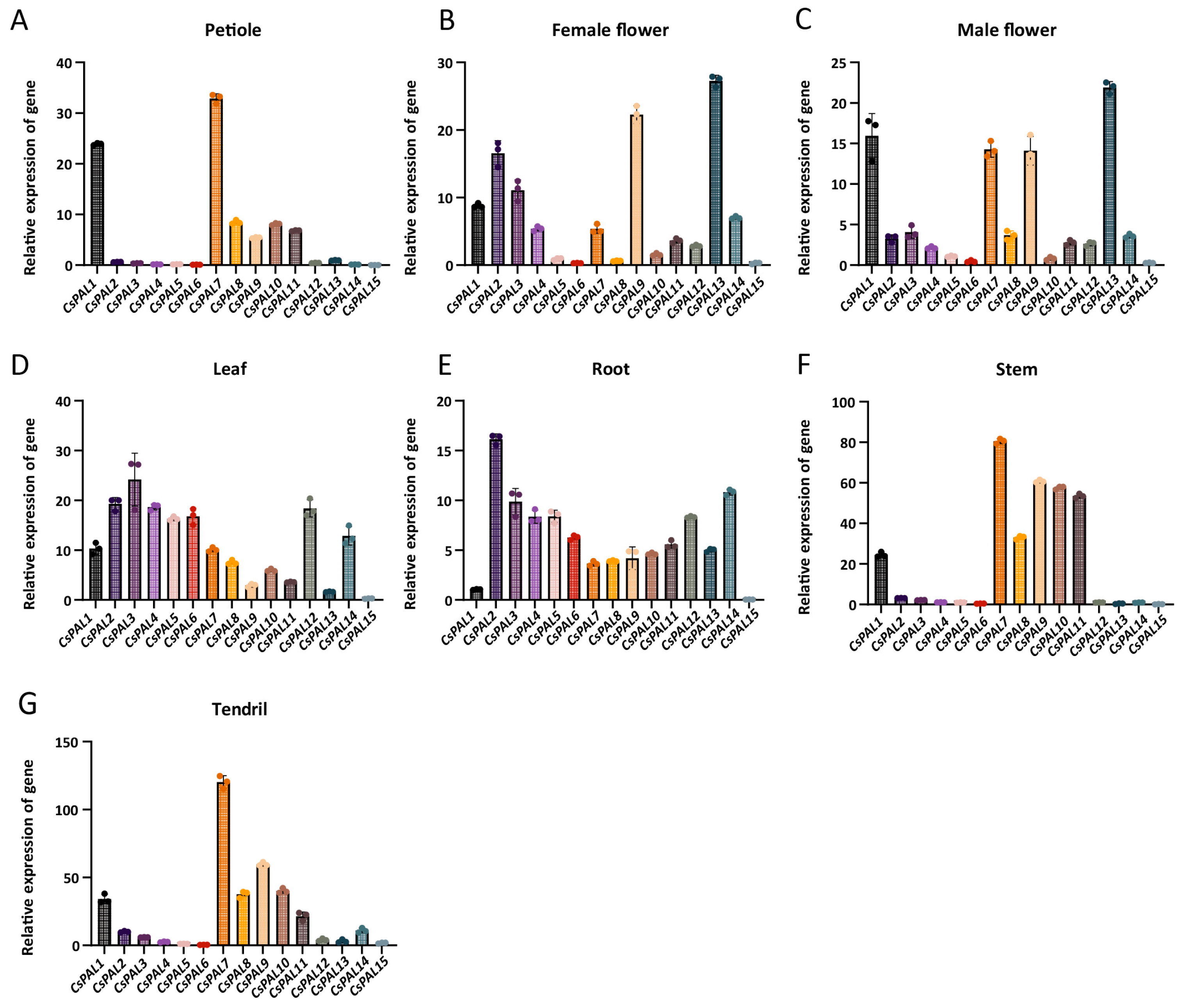
2.5. Expression Patterns of CsPAL in Different Tissues
2.6. Expression Patterns of CsPAL under Abiotic Stress Conditions
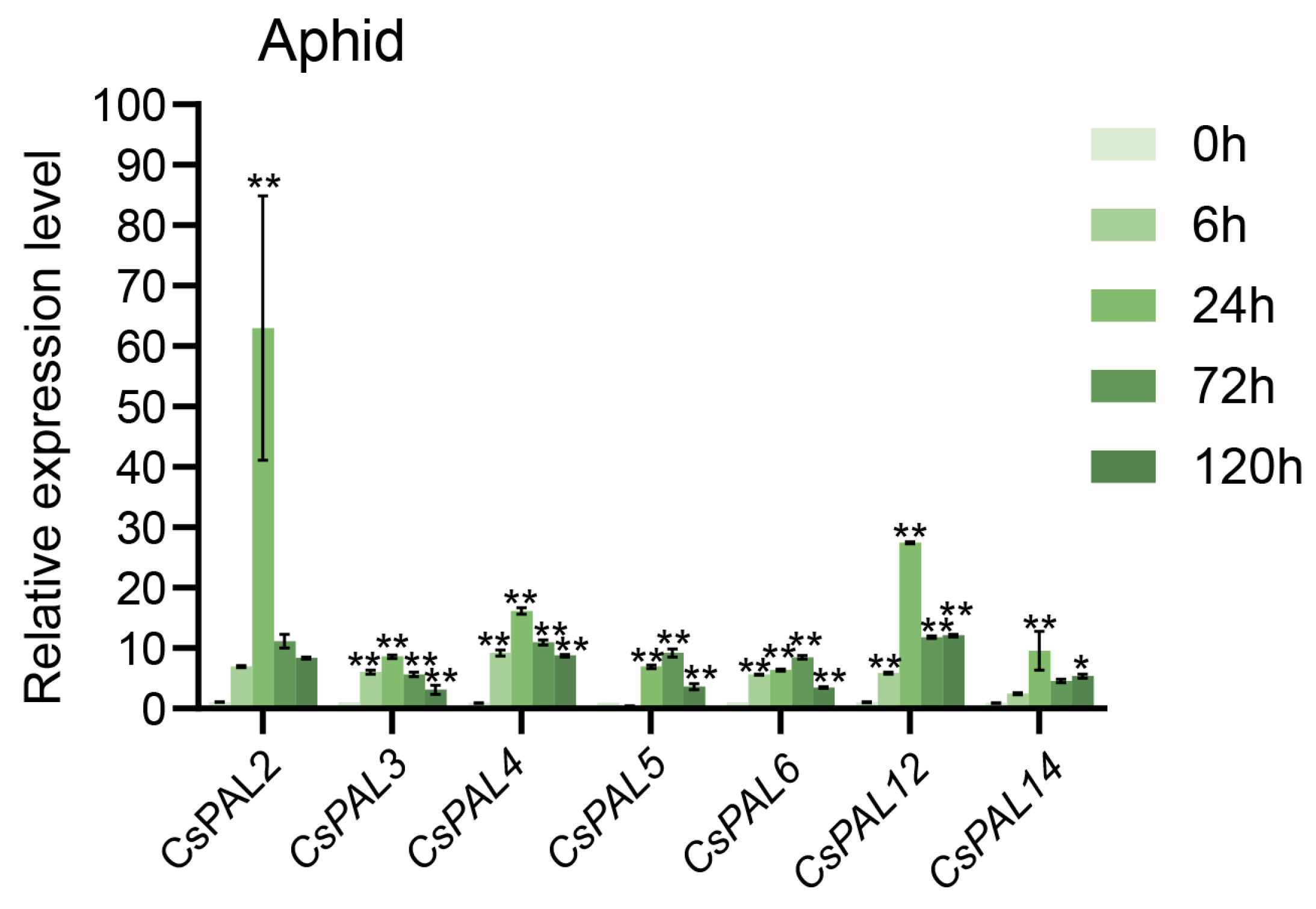
2.7. Expression Patterns of CsPAL under Aphid Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growing Conditions
4.2. Aphid and Abiotic Stress Treatment
4.3. Identification of CsPAL Gene Family Members in Cucumber
4.4. Physicochemical Properties of CsPAL Family Proteins
4.5. Analysis of Conserved Motifs, Gene Structure, Cis-Acting Elements, Multiple Sequence Alignment, and Phylogenetic Analysis
4.6. Chromosomal Localization and Ka/Ks Analysis
4.7. Protein-Protein Interaction Prediction
4.8. RT-qPCR Expression Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacDonald, M.J.; D’Cunha, G.B. A modern view of phenylalanine ammonia lyase. Biochem. Cell Biol. 2007, 85, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Gharibi, S.; Sayed Tabatabaei, B.E.; Saeidi, G.; Talebi, M.; Matkowski, A. The effect of drought stress on polyphenolic compounds and expression of flavonoid biosynthesis related genes in Achillea pachycephala Rech.f. Phytochemistry 2019, 162, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Gho, Y.S.; Kim, S.J.; Jung, K.H. Phenylalanine ammonia-lyase family is closely associated with response to phosphate deficiency in rice. Genes Genom. 2020, 42, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Khakdan, F.; Alizadeh, H.; Ranjbar, M. Molecular cloning, functional characterization and expression of a drought inducible phenylalanine ammonia-lyase gene (ObPAL) from Ocimum basilicum L. Plant Physiol. Biochem. 2018, 130, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Koukol, J.; Conn, E.E. The metabolism of aromatic compounds in higher plants. IV. Purification and properties of the phenylalanine deaminase of Hordeum vulgare. J. Biol. Chem. 1961, 236, 2692–2698. [Google Scholar] [CrossRef]
- Ohl, S.; Hedrick, S.A.; Chory, J.; Lamb, C.J. Functional properties of a phenylalanine ammonia-lyase promoter from Arabidopsis. Plant Cell 1990, 2, 837–848. [Google Scholar] [CrossRef]
- Costa, M.A.; Collins, R.E.; Anterola, A.M.; Cochrane, F.C.; Davin, L.B.; Lewis, N.G. An in silico assessment of gene function and organization of the phenylpropanoid pathway metabolic networks in Arabidopsis thaliana and limitations thereof. Phytochemistry 2003, 64, 1097–1112. [Google Scholar] [CrossRef]
- Rohde, A.; Morreel, K.; Ralph, J.; Goeminne, G.; Hostyn, V.; De Rycke, R.; Kushnir, S.; Van Doorsselaere, J.; Joseleau, J.P.; Vuylsteke, M.; et al. Molecular phenotyping of the pal1 and pal2 mutants of Arabidopsis thaliana reveals far-reaching consequences on phenylpropanoid, amino acid, and carbohydrate metabolism. Plant Cell 2004, 16, 2749–2771. [Google Scholar] [CrossRef]
- Wong, J.H.; Namasivayam, P.; Abdullah, M.P. The PAL2 promoter activities in relation to structural development and adaptation in Arabidopsis thaliana. Planta 2012, 235, 267–277. [Google Scholar] [CrossRef]
- Cochrane, F.C.; Davin, L.B.; Lewis, N.G. The Arabidopsis phenylalanine ammonia lyase gene family: Kinetic characterization of the four PAL isoforms. Phytochemistry 2004, 65, 1557–1564. [Google Scholar] [CrossRef]
- Mo, F.; Li, L.; Zhang, C.; Yang, C.; Chen, G.; Niu, Y.; Si, J.; Liu, T.; Sun, X.; Wang, S.; et al. Genome-Wide Analysis and Expression Profiling of the Phenylalanine Ammonia-Lyase Gene Family in Solanum tuberosum. Int. J. Mol. Sci. 2022, 23, 6833. [Google Scholar] [CrossRef]
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Chen, Z. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010, 153, 1526–1538. [Google Scholar] [CrossRef]
- Zhang, R.; Li, L.; Xue, H.; Bi, Y.; Raza, H.; Si, M.; Peng, H.; Nan, M.; Zong, Y.; Prusky, D. Ca2+ applications affect the phenylpropanoid metabolism in potato tubers induced by T-2 toxin. Postharvest Biol. Technol. 2021, 180, 111616. [Google Scholar] [CrossRef]
- Wang, W.; Wang, X.; Zhang, X.; Wang, Y.; Huo, Z.; Huang, M.; Cai, J.; Zhou, Q.; Jiang, D. Involvement of salicylic acid in cold priming-induced freezing tolerance in wheat plants. Plant Growth Regul. 2021, 93, 117–130. [Google Scholar] [CrossRef]
- Koramutla, M.K.; Tuan, P.A.; Ayele, B.T. Salicylic Acid Enhances Adventitious Root and Aerenchyma Formation in Wheat under Waterlogged Conditions. Int. J. Mol. Sci. 2022, 23, 1243. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Jiang, T.; Du, K.; Chen, H.; Cao, Y.; Xie, J.; Li, M.; Carr, J.P.; Wu, B.; Fan, Z.; et al. Maize phenylalanine ammonia-lyases contribute to resistance to Sugarcane mosaic virus infection, most likely through positive regulation of salicylic acid accumulation. Mol. Plant Pathol. 2019, 20, 1365–1378. [Google Scholar] [CrossRef]
- He, J.; Liu, Y.; Yuan, D.; Duan, M.; Liu, Y.; Shen, Z.; Yang, C.; Qiu, Z.; Liu, D.; Wen, P.; et al. An R2R3 MYB transcription factor confers brown planthopper resistance by regulating the phenylalanine ammonia-lyase pathway in rice. Proc. Natl. Acad. Sci. USA 2020, 117, 271–277. [Google Scholar] [CrossRef]
- Luo, K.; Yao, X.J.; Luo, C.; Hu, X.S.; Hu, Z.Q.; Zhang, G.S.; Zhao, H.Y. Previous Aphid Infestation Induces Different Expression Profiles of Genes Associated with Hormone-Dependent Responses in Near-Isogenic Winter Wheat Lines. J. Econ. Entomol. 2020, 113, 461–470. [Google Scholar] [CrossRef]
- Gebretsadik, K.; Qiu, X.; Dong, S.; Miao, H.; Bo, K. Molecular research progress and improvement approach of fruit quality traits in cucumber. Theor. Appl. Genet. 2021, 134, 3535–3552. [Google Scholar] [CrossRef]
- Gou, C.; Zhu, P.; Meng, Y.; Yang, F.; Xu, Y.; Xia, P.; Chen, J.; Li, J. Evaluation and Genetic Analysis of Parthenocarpic Germplasms in Cucumber. Genes 2022, 13, 225. [Google Scholar] [CrossRef]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef]
- Li, Q.; Li, H.; Huang, W.; Xu, Y.; Zhou, Q.; Wang, S.; Ruan, J.; Huang, S.; Zhang, Z. A chromosome-scale genome assembly of cucumber (Cucumis sativus L.). GigaScience 2019, 8, giz072. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Duan, L.; Yu, J.; Su, C.; Li, J.; Chen, D.; Zhang, X.; Song, H.; Pan, Y. Characterization analysis and heavy metal-binding properties of CsMTL3 in Escherichia coli. FEBS Open Bio 2018, 8, 1820–1829. [Google Scholar] [CrossRef]
- Xu, X.; Liu, M.; Hu, Q.; Yan, W.; Pan, J.; Yan, Y.; Chen, X. A CsEIL3-CsARN6.1 module promotes waterlogging-triggered adventitious root formation in cucumber by activating the expression of CsPrx5. Plant J. 2023, 114, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Cao, M.; Li, Y.; Wang, J.; He, L.; Li, P.; Lin, X.; Li, X.; Yuan, X.; Zhao, B.; et al. RING finger ubiquitin E3 ligase CsCHYR1 targets CsATAF1 for degradation to modulate the drought stress response of cucumber through the ABA-dependent pathway. Plant Physiol. Biochem. 2023, 202, 107928. [Google Scholar] [CrossRef] [PubMed]
- El-Remaly, E. Morphological, physio-biochemical, and molecular indications of heat stress tolerance in cucumber. Sci. Rep. 2023, 13, 18729. [Google Scholar] [CrossRef]
- Li, Y.; Fan, H.; Si, Y.; Guo, Y.; Wu, Z.; Du, C. The phloem lectin PP2-A1 enhances aphid resistance by affecting aphid behavior and maintaining ROS homeostasis in cucumber plants. Int. J. Biol. Macromol. 2023, 229, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Raes, J.; Rohde, A.; Christensen, J.H.; Van de Peer, Y.; Boerjan, W. Genome-wide characterization of the lignification toolbox in Arabidopsis. Plant Physiol. 2003, 133, 1051–1071. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Paiva, N.L. Stress-Induced Phenylpropanoid Metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Weisshaar, B.; Jenkins, G.I. Phenylpropanoid biosynthesis and its regulation. Curr. Opin. Plant Biol. 1998, 1, 251–257. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, C.J. Multifaceted regulations of gateway enzyme phenylalanine ammonia-lyase in the biosynthesis of phenylpropanoids. Mol. Plant 2014, 8, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.M.; Li, L.; Dong, C.J. Multiple tandem duplication of the phenylalanine ammonia-lyase genes in Cucumis sativus L. Planta 2012, 236, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- Qian, M.; Kalbina, I.; Rosenqvist, E.; Jansen, M.A.K.; Teng, Y.; Strid, Å. UV regulates the expression of phenylpropanoid biosynthesis genes in cucumber (Cucumis sativus L.) in an organ and spectrum dependent manner. Photochem. Photobiol. Sci. 2019, 18, 424–433. [Google Scholar] [CrossRef]
- Nawaz, S.; Bano, A. Effects of PGPR (Pseudomonas sp.) and Ag-nanoparticles on Enzymatic Activity and Physiology of Cucumber. Recent Pat. Food Nutr. Agric. 2020, 11, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.Y.; Sattler, S.A.; Cortez, G.S.; Vermerris, W.; Sattler, S.E.; Kang, C. Biochemical and Structural Analysis of Substrate Specificity of a Phenylalanine Ammonia-Lyase. Plant Physiol. 2018, 176, 1452–1468. [Google Scholar] [CrossRef]
- Olsen, K.M.; Lea, U.S.; Slimestad, R.; Verheul, M.; Lillo, C. Differential expression of four Arabidopsis PAL genes; PAL1 and PAL2 have functional specialization in abiotic environmental-triggered flavonoid synthesis. J. Plant Physiol. 2008, 165, 1491–1499. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, S.; Chu, D.; Wang, X. Exploring the evolution of CHS gene family in plants. Front. Genet. 2024, 15, 1368358. [Google Scholar] [CrossRef]
- Glagoleva, A.Y.; Ivanisenko, N.V.; Khlestkina, E.K. Organization and evolution of the chalcone synthase gene family in bread wheat and relative species. BMC Genet. 2019, 20, 30. [Google Scholar] [CrossRef]
- Deng, Y.; Li, C.; Li, H.; Lu, S. Identification and Characterization of Flavonoid Biosynthetic Enzyme Genes in Salvia miltiorrhiza (Lamiaceae). Molecules 2018, 23, 1467. [Google Scholar] [CrossRef]
- Costa, M.A.; Bedgar, D.L.; Moinuddin, S.G.A.; Kim, K.-W.; Cardenas, C.L.; Cochrane, F.C.; Shockey, J.M.; Helms, G.L.; Amakura, Y.; Takahashi, H.; et al. Characterization in vitro and in vivo of the putative multigene 4-coumarate:CoA ligase network in Arabidopsis: Syringyl lignin and sinapate/sinapyl alcohol derivative formation. Phytochemistry 2005, 66, 2072–2091. [Google Scholar] [CrossRef]
- Li, Y.; Kim, J.I.; Pysh, L.; Chapple, C. Four Isoforms of Arabidopsis 4-Coumarate:CoA Ligase Have Overlapping yet Distinct Roles in Phenylpropanoid Metabolism. Plant Physiol. 2015, 169, 2409–2421. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Li, Y.; Feng, S.; Zou, W.; Guo, K.; Fan, C.; Si, S.; Peng, L. Analysis of five rice 4-coumarate:coenzyme A ligase enzyme activity and stress response for potential roles in lignin and flavonoid biosynthesis in rice. Biochem. Biophys. Res. Commun. 2013, 430, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Cesarino, I. Structural features and regulation of lignin deposited upon biotic and abiotic stresses. Curr. Opin. Biotechnol. 2019, 56, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Du, L.; Cao, Q.; Yang, Z.; Liu, Y.; Yang, H.; Duan, X.; Meng, Z. Salt Tolerance Evaluation of Cucumber Germplasm under Sodium Chloride Stress. Plants 2023, 12, 2927. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wu, X.; Liu, S.; Zhang, L.; Li, T.; Cao, Y.; Duan, Q. Comprehensive Analysis of BrHMPs Reveals Potential Roles in Abiotic Stress Tolerance and Pollen-Stigma Interaction in Brassica rapa. Cells 2023, 12, 1096. [Google Scholar] [CrossRef]
- Sohail, H.; Noor, I.; Nawaz, M.A.; Ma, M.; Shireen, F.; Huang, Y.; Yang, L.; Bie, Z. Genome-wide identification of plasma-membrane intrinsic proteins in pumpkin and functional characterization of CmoPIP1-4 under salinity stress. Environ. Exp. Bot. 2022, 202, 104995. [Google Scholar] [CrossRef]

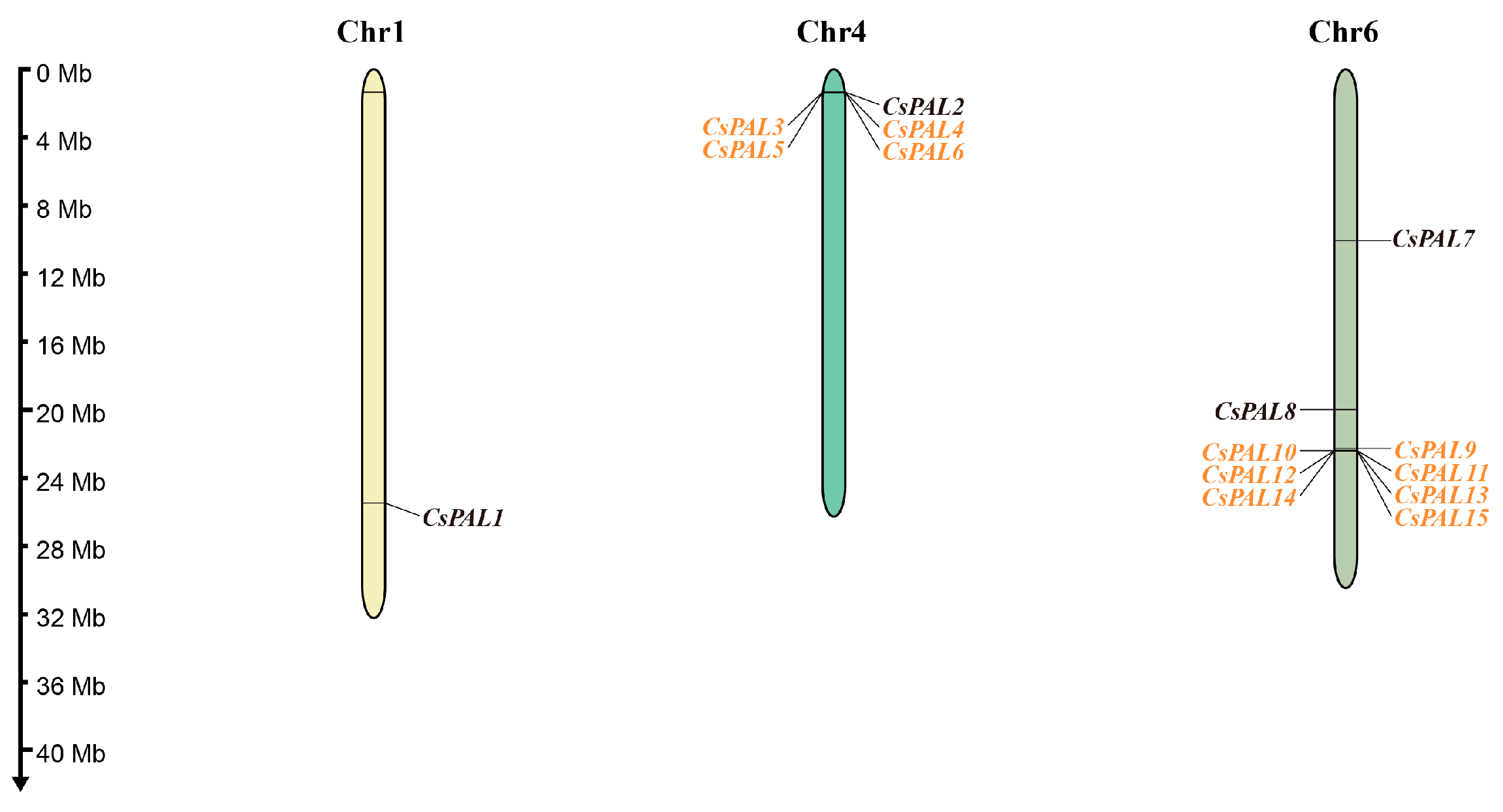

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, J.; Sohail, H.; Qiu, L.; Chen, C.; Yue, H.; Li, Z.; Yang, X.; Zhang, L. Genome-Wide Characterization and Expression Analysis of CsPALs in Cucumber (Cucumis sativus L.) Reveal Their Potential Roles in Abiotic Stress and Aphid Stress Tolerance. Plants 2024, 13, 2537. https://doi.org/10.3390/plants13182537
Gu J, Sohail H, Qiu L, Chen C, Yue H, Li Z, Yang X, Zhang L. Genome-Wide Characterization and Expression Analysis of CsPALs in Cucumber (Cucumis sativus L.) Reveal Their Potential Roles in Abiotic Stress and Aphid Stress Tolerance. Plants. 2024; 13(18):2537. https://doi.org/10.3390/plants13182537
Chicago/Turabian StyleGu, Jieni, Hamza Sohail, Lei Qiu, Chaoyan Chen, Haoyu Yue, Ziyi Li, Xiaodong Yang, and Lili Zhang. 2024. "Genome-Wide Characterization and Expression Analysis of CsPALs in Cucumber (Cucumis sativus L.) Reveal Their Potential Roles in Abiotic Stress and Aphid Stress Tolerance" Plants 13, no. 18: 2537. https://doi.org/10.3390/plants13182537
APA StyleGu, J., Sohail, H., Qiu, L., Chen, C., Yue, H., Li, Z., Yang, X., & Zhang, L. (2024). Genome-Wide Characterization and Expression Analysis of CsPALs in Cucumber (Cucumis sativus L.) Reveal Their Potential Roles in Abiotic Stress and Aphid Stress Tolerance. Plants, 13(18), 2537. https://doi.org/10.3390/plants13182537