
A Plant Strategy: Irrigation, Nitrogen Fertilization, and Climatic Conditions Regulated the Carbon Allocation and Yield of Oilseed Flax in Semi-Arid Area


Abstract
1. Introduction
2. Results
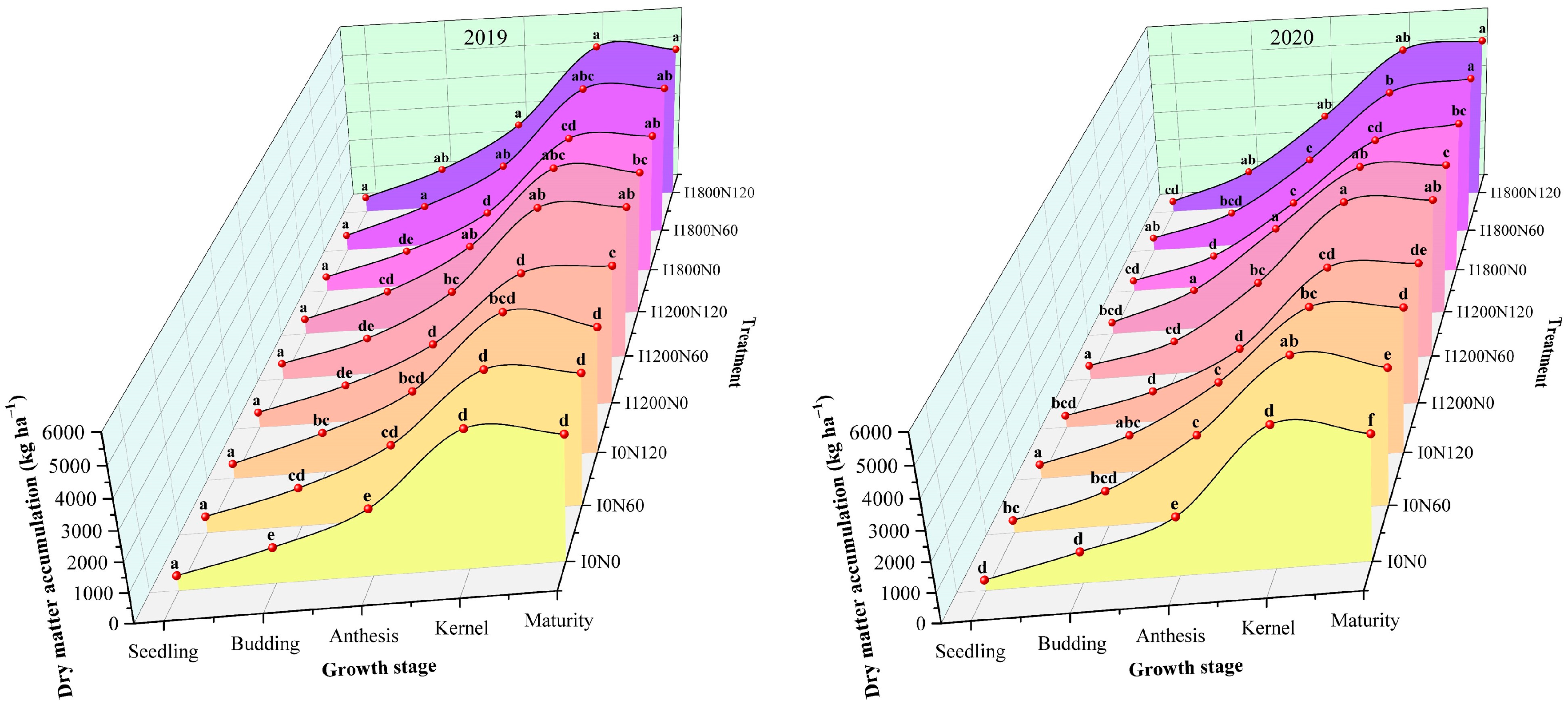
2.1. Dry Matter Accumulation under Different Treatments
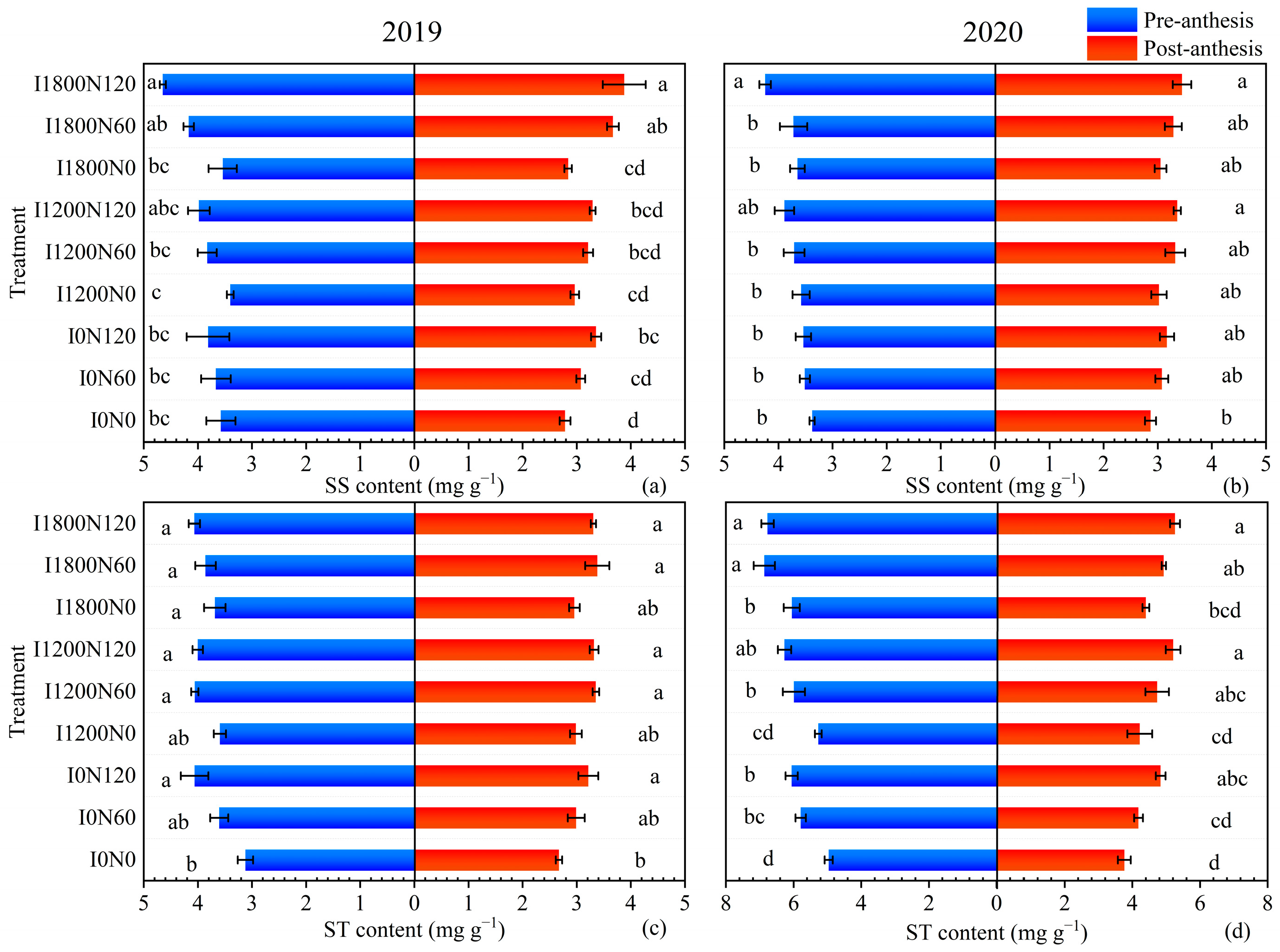
2.2. Soluble Sugar and Starch Content under Different Treatments
2.2.1. Variance Analysis of Soluble Sugar and Starch Content in Various Organs of Oilseed Flax at Pre-Anthesis and Post-Anthesis
2.2.2. Soluble Sugar and Starch Content in Leaves
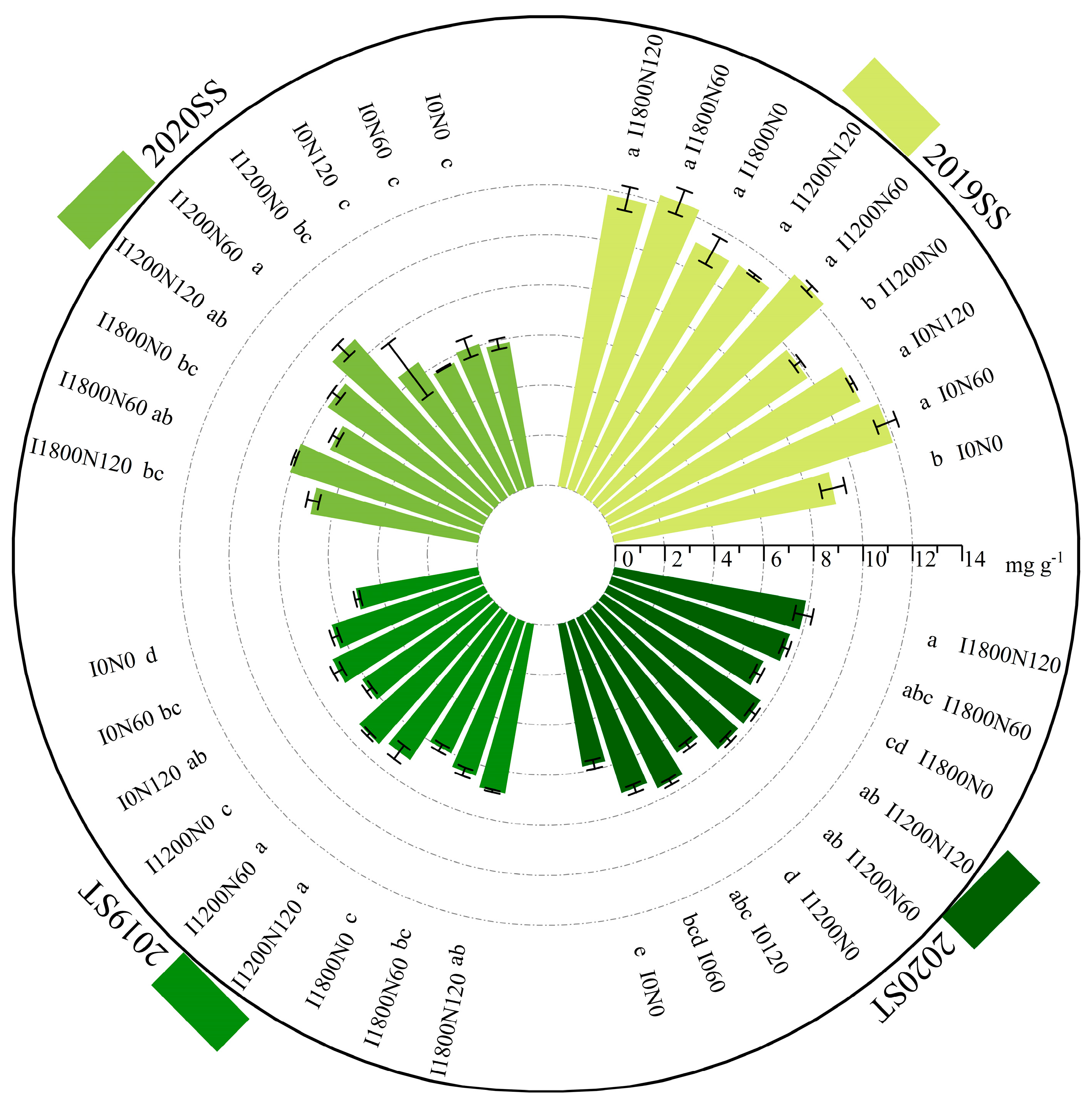
2.2.3. Soluble Sugar and Starch Content in Stems
2.2.4. Soluble Sugar and Starch Content in Capsules
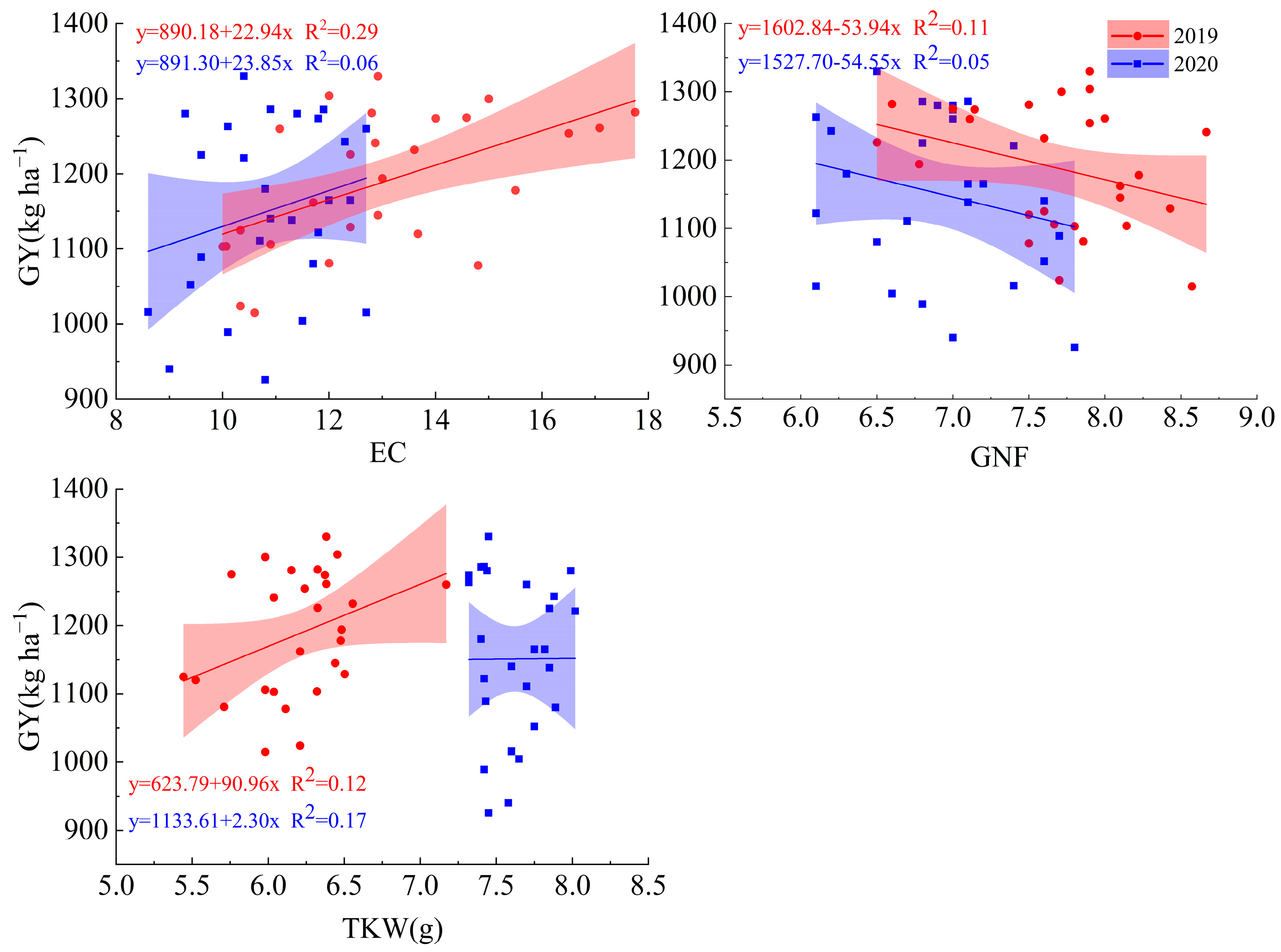
2.2.5. Correlation of Soluble Sugar and Starch Content with Yield
2.3. Yield Performance
2.3.1. Yield Components and Grain Yield
2.3.2. Yield Increase Rate and Contribution to Yield Increase
2.4. Water and Nitrogen Productivity under Different Treatments
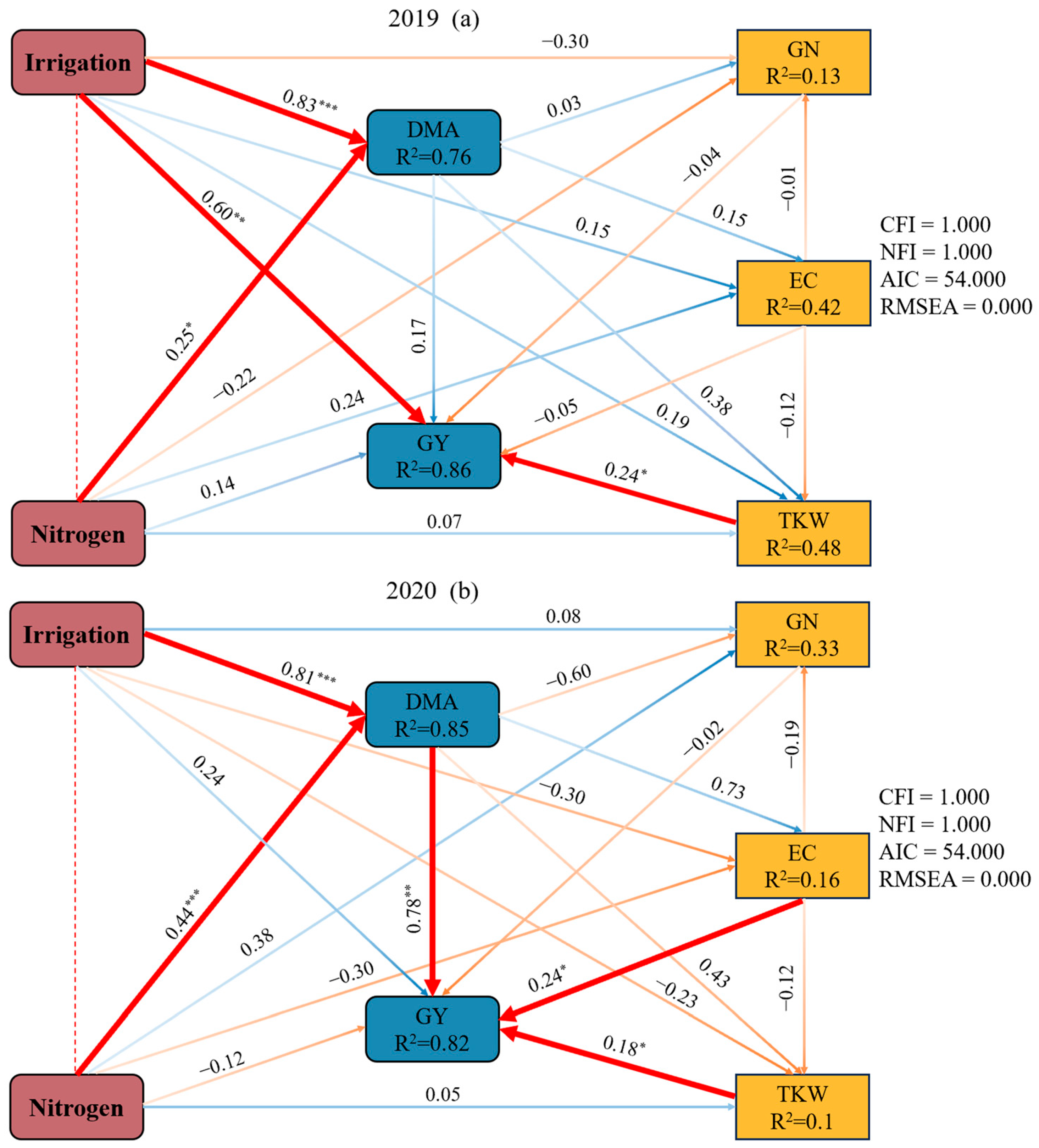
2.5. The Effects of Irrigation and Nitrogen on Dry Matter Accumulation at the Maturity Stage, Grain Yield, and Yield Components Based on a Structure Equation Model
3. Discussions
3.1. Dry Matter Accumulation
3.2. Non-Structural Carbohydrates
3.3. Grain Yield and Yield Components
3.4. Water Use Efficiency and Nitrogen Partial Factor Productivity
4. Materials and Methods
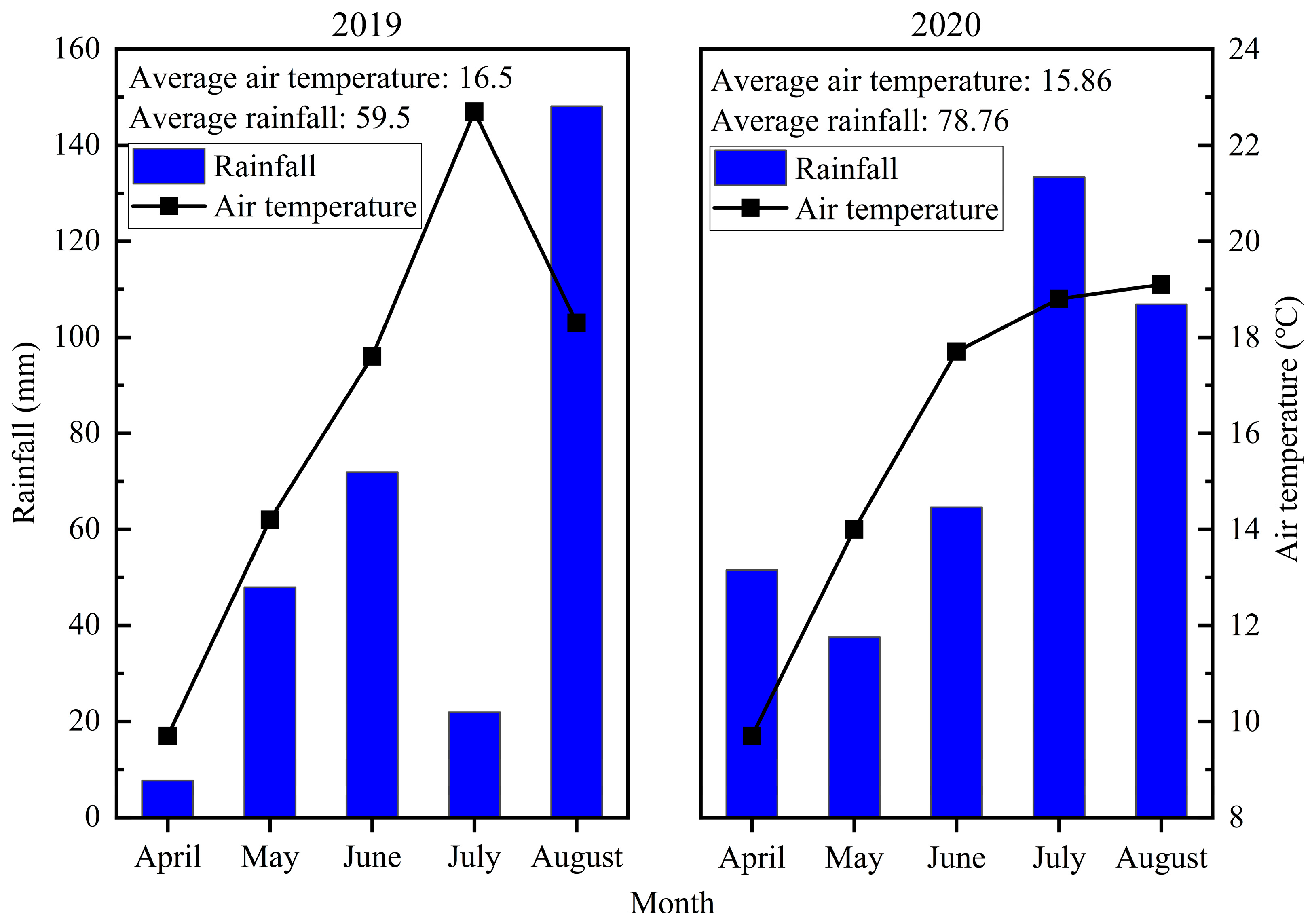
4.1. Site Description
4.2. Experimental Design and Field Management
4.3. Date Collection
4.3.1. Dry Matter Accumulation
4.3.2. Soluble Sugar and Starch
4.3.3. Grain Yield and Yield Composition of Oilseed Flax
4.3.4. The Contribution to Yield Increase of Oilseed Flax
4.3.5. Water and Nitrogen Productivity
4.3.6. Nitrogen Partial Factor Productivity (NPFP)
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. 2021. Available online: https://www.fao.org/land-water/solaw2021/facts/en/ (accessed on 2 July 2024).
- Zhang, Y.; Ma, Q.; Liu, D.; Sun, L.; Ren, X.; Ali, S.; Zhang, P.; Jia, Z. Effects of different fertilizer strategies on soil water utilization and maize yield in the ridge and furrow rainfall harvesting system in semiarid regions of China. Agric. Water Manag. 2018, 208, 414–421. [Google Scholar] [CrossRef]
- Xu, J.; Cai, H.; Wang, X.; Ma, C.; Lu, Y.; Ding, Y.; Wang, X.; Chen, H.; Wang, Y.; Saddique, Q. Exploring optimal irrigation and nitrogen fertilization in a winter wheat-summer maize rotation system for improving crop yield and reducing water and nitrogen leaching. Agric. Water Manag. 2020, 228, 105904. [Google Scholar] [CrossRef]
- Zheng, M.; Chen, J.; Shi, Y.; Li, Y.; Yin, Y.; Yang, D.; Luo, Y.; Pang, D.; Xu, X.; Li, W.; et al. Manipulation of lignin metabolism by plant densities and its relationship with lodging resistance in wheat. Sci. Rep. 2017, 7, 41805. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, X.; Zhang, X.; Zhang, L.; Li, Y.; Huang, G. Evaluation on the responses of maize (Zea mays L.) growth, yield and water use efficiency to drip irrigation water under mulch condition in the Hetao irrigation District of China. Agric. Water Manag. 2016, 179, 144–157. [Google Scholar] [CrossRef]
- Behera, S.K.; Panda, R.K. Integrated management of irrigation water and fertilizers for wheat crop using field experiments and simulation modeling. Agric. Water Manag. 2009, 96, 1532–1540. [Google Scholar] [CrossRef]
- Song, F.; Zhuge, Y.; Guo, X.; Lou, Y.; Wang, H.; Pan, T.; Feng, H. Optimizing irrigation and fertilization can improve degraded saline soils and increase wheat grain yield. Land Degrad. Dev. 2021, 32, 494–504. [Google Scholar] [CrossRef]
- Gao, R.; Pan, Z.; Zhang, J.; Chen, X.; Qi, Y.; Zhang, Z.; Chen, S.; Jiang, K.; Ma, S.; Wang, J.; et al. Optimal cooperative application solutions of irrigation and nitrogen fertilization for high crop yield and friendly environment in the semi-arid region of North China. Agric. Water Manag. 2023, 283, 108326. [Google Scholar] [CrossRef]
- Yagioka, A.; Hayashi, S.; Kimiwada, K.; Kondo, M. Kitagenki, a high-yielding rice variety, exhibits a high yield potential under optimum crop management practices. Eur. J. Agron. 2022, 140, 126606. [Google Scholar] [CrossRef]
- Gao, Y.H.; Jiang, H.Y.; Wu, B.; Niu, J.Y.; Li, Y.J.; Guo, F.; Wang, L.; Blakney, A.J. The effects of planting density on lodging resistance, related enzyme activities, and grain yield in different genotypes of oilseed flax. Crop Sci. 2018, 58, 2613–2625. [Google Scholar] [CrossRef]
- Du, J.; Yang, P.; Li, Y.; Ren, S.; Wang, Y.; Li, X.; He, X. Influence of the irrigation, fertilization and groundwater depth on wheat yield and nitrate nitrogen leaching. Trans. Chin. Soc. Agric. Eng. 2011, 27, 57–64. [Google Scholar]
- Tan, Y.C.; Xu, C.; Liu, D.X.; Wu, W.L.; Lal, R.; Meng, F.Q. Effects of optimized N fertilization on greenhouse gas emission and crop production in the North China Plain. Field Crops Res. 2017, 205, 135–146. [Google Scholar] [CrossRef]
- West, P.C.; Gerber, J.S.; Engstrom, P.M.; Mueller, N.D.; Brauman, K.A.; Carlson, K.M.; Cassidy, E.S.; Johnston, M.; MacDonald, G.K.; Ray, D.K.; et al. Leverage points for improving global food security and the environment. Science 2014, 345, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, G.H.; Chen, Z.J.; Xiong, Y.W.; Huang, Q.Z.; Xu, X.; Huo, Z. Effects of irrigation and fertilization on grain yield, water and nitrogen dynamics and their use efficiency of spring wheat farmland in an arid agricultural watershed of Northwest China. Agric. Water Manag. 2022, 260, 107277. [Google Scholar] [CrossRef]
- Fu, Q.P.; Wang, Q.J.; Shen, X.L.; Fan, J. Optimizing water and nitrogen inputs for winter wheat cropping system on the Loess Plateau, China. J. Arid. Land 2014, 6, 230–242. [Google Scholar] [CrossRef]
- Wang, X.; Xing, Y. Evaluation of the effects of irrigation and fertilization on tomato fruit yield and quality: A principal component analysis. Sci. Rep. 2017, 7, 350–358. [Google Scholar] [CrossRef]
- Gheysari, M.; Miriatifi, S.M.; Bannayan, M.; Homaee, M.; Hoogenboom, G. Interaction of water and nitrogen on maize grown for silage. Agric. Water Manag. 2009, 96, 809–821. [Google Scholar] [CrossRef]
- Wang, X.Y.; He, M.R.; Li, F.; Liu, Y.H.; Zhang, H.H.; Liu, C.G. Coupling effects of irrigation and nitrogen fertilization on grain protein and starch quality of strong-gluten winter wheat. Front. Agric. China 2008, 2, 274–280. [Google Scholar] [CrossRef]
- Liu, Y.; Liao, Y.C.; Liu, W.Z. High nitrogen application rate and planting density reduce wheat grain yield by reducing filling rate of inferior grain in middle spikelets. Crop J. 2021, 9, 412–426. [Google Scholar] [CrossRef]
- Yang, X.; Mcmaster, G.S.; Yu, Q. Spatial patterns of relationship between wheat yield and yield components in China. Int. J. Plant Prod. 2018, 12, 61–71. [Google Scholar] [CrossRef]
- Grafius, J.E. Multiple characters and correlated response. Crop Sci. 1978, 18, 931–934. [Google Scholar] [CrossRef]
- Kong, J.J.; Liu, W.Q.; Huang, F.; Hua, L.; Yu, F.; He, Q.; Su, Y.; Li, J.Y.; Qiu, Q. Spatial Patterns of Non-Structural Carbohydrates in Eucalyptus urophylla × E. grandis under Dry-Season Irrigation with Fertilization. Forests 2021, 12, 1049. [Google Scholar] [CrossRef]
- Carpenter, L.T.; Pezeshki, S.R.; Shields, F.D. Responses of nonstructural carbohydrates to shoot removal and soil moisture treatments in Salix nigra. Trees 2008, 22, 737–748. [Google Scholar] [CrossRef]
- Cannell, M.G.R.; Dewar, R.C. Carbon allocation in trees: A review of concepts for modelling. Adv. Ecol. Res. 1994, 25, 59–104. [Google Scholar]
- Yan, B.; Wu, B.; Gao, Y.H.; Wu, J.M.; Niu, J.Y.; Xie, Y.P.; Cui, Z.; Zhang, Z. Effects of nitrogen and phosphorus on the regulation of nonstructural carbohydrate accumulation, translocation and the yield formation of oilseed flax. Field Crops Res. 2018, 219, 229–241. [Google Scholar] [CrossRef]
- Hiroshi, W.; Chisato, M.K.; Koichi, T.; Hiroshi, N.; Fukuyo, T.; Haruka, O.; Rosa, E.B.; Kenzo, H.; Taiken, N.; Makoto, H.; et al. Turgor-responsive starch phosphorylation in Oryza sativa stems: A primary event of starch degradation associated with grain-filling ability. PLoS ONE 2017, 12, e0181272. [Google Scholar]
- Wu, W.; Ma, B.L.; Fan, J.J.; Sun, M.; Yi, Y.; Guo, W.S.; Voldeng, H.D. Management of nitrogen fertilization to balance reducing lodging risk and increasing yield and protein content in spring wheat. Field Crops Res. 2019, 241, 107584. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, B.; Li, Q.; Kong, F.; Du, L.; Zhou, F.; Shi, H.; Ke, Y.; Liu, Q.; Feng, D.; et al. Non-structural carbohydrates in maize with different nitrogen tolerance are affected by nitrogen addition. PLoS ONE 2019, 14, e0225753. [Google Scholar] [CrossRef]
- Lu, J.; Hu, T.; Geng, C.; Cui, X.; Fan, J.; Zhang, F. Response of yield, yield components and water-nitrogen use efficiency of winter wheat to different drip fertigation regimes in Northwest China. Agric. Water Manag. 2021, 255, 107034. [Google Scholar] [CrossRef]
- Latt, C.R.; Nair, P.K.R.; Kang, B.T. Reserve carbohydrate levels in the boles and structural roots of five multipurpose tree species in a seasonally dry tropical climate. For. Ecol. Manag. 2001, 146, 145–158. [Google Scholar] [CrossRef]
- von Arx, G.; Arzac, A.; Fonti, P.; Frank, D.; Zweifel, R.; Rigling, A.; Galiano, L.; Gessler, A.; Olano, J.M. Responses of sapwood ray parenchyma and non-structural carbohydrates of Pinus sylvestris to drought and long-term irrigation. Funct. Ecol. 2017, 31, 1371–1382. [Google Scholar] [CrossRef]
- Torres, N.; Yu, R.; Martínez-Lüscher, J.; Kostaki, E.; Kurtural, S.K. Application of fractions of crop evapotranspiration affects carbon partitioning of grapevine differentially in a hot climate. Front. Plant Sci. 2021, 12, 633600. [Google Scholar] [CrossRef] [PubMed]
- Dordas, C.A. Nitrogen and dry matter dynamics in linseed as affected by the nitrogen level and genotype in a Mediterranean environment. Biomass Bioenergy 2012, 43, 1–11. [Google Scholar] [CrossRef]
- Xie, Y.P.; Gan, Y.T.; Li, Y.; Niu, J.Y.; Gao, Y.H.; An, H.H.; Li, A. Effect of Nitrogen Fertilizer on Nitrogen Accumulation, Translocation, and Use Efficiency in Dryland Oilseed Flax. Agron. J. 2015, 107, 1931–1939. [Google Scholar] [CrossRef]
- Zhang, J.P.; Long, Y.; Wang, L.M.; Dang, Z.; Zhang, T.B.; Song, X.X.; Dang, Z.; Pei, X. Consensus genetic linkage map construction and QTL mapping for plant height-related traits in linseed flax (Linum usitatissimum L.). BMC Plant Biol. 2018, 18, 160. [Google Scholar] [CrossRef]
- Dzuvor, C.K.O.; Taylor, J.T.; Acquah, C.; Pan, S.; Agyei, D. Bioprocessing of functional ingredients from flaxseed. Molecules 2018, 23, 2444. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef]
- Zhao, B.X.; Wang, L.; Yu, H.L.; Li, Y.Y.; Tan, M.L.; Shi, X.H.; Yang, J.Z.; Ma, Q.F. Effects of irrigation period and frequency on yield and quality of Linseed. Guangdong Agric. Sci. 2023, 50, 19–26. [Google Scholar]
- Cui, H.Y.; Hu, F.L.; Fang, Z.S.; Niu, J.Y. Effects of irrigation amount and stage on water consumption characteristics and grain yield of oil flax. J. Nucl. Agric. Sci. 2015, 29, 812–819. [Google Scholar]
- Cui, H.Y.; Fang, Z.S. Effect of nitrogen and irrigation interaction on dry matter production and grain yield of oil flax under different irrigation modes. Acta Bot. Boreali-Occident. Sin. 2016, 36, 156–164. [Google Scholar]
- Cui, Z.J.; Liu, D.; Wu, B.; Yan, B.; Ma, J.; Zhao, B.Q.; Gao, Y.H.; Niu, J.Y. Effects of water and nitrogen coupling on grain yield formation and nitrogen accumulation, transportation of oil flax in dryland. Chin. J. Appl. Ecol. 2020, 31, 909–918. [Google Scholar]
- Ma, Q.; Wang, M.Y.; Zheng, G.L.; Yao, Y.; Tao, R.R.; Zhu, M.; Ding, J.; Li, C.; Guo, W.; Zhu, X. Twice-split application of controlled-release nitrogen fertilizermet the nitrogen demand of winter wheat. Field Crops Res. 2021, 267, 108163. [Google Scholar] [CrossRef]
- Sun, L.; Wang, R.; Li, J.; Wang, Q.; Lyu, W.; Wang, X.L.; Cheng, K.; Mao, H.; Zhang, X. Reasonable fertilization improves the conservation tillage benefit for soil water use and yield of rain-fed winter wheat: A case study from the Loess Plateau, China. Field Crop Res. 2019, 242, 107589. [Google Scholar] [CrossRef]
- Li, Y.; Song, H.; Zhou, L.; Xu, Z.; Zhou, G. Vertical distributions of chlorophyll and nitrogen and their associations with photosynthesis under drought and rewatering regimes in a maize field. Agric. For. Meteorol. 2019, 272, 40–54. [Google Scholar] [CrossRef]
- Wang, X.; Xiang, Y.Z.; Guo, J.J.; Tang, Z.J.; Zhao, S.T.; Wang, H.; Li, Z.; Zhang, F. Coupling effect analysis of drip irrigation and mixed slow-release nitrogen fertilizer on yield and physiological characteristics of winter wheat in Guanzhong area. Field Crops Res. 2023, 302, 109103. [Google Scholar] [CrossRef]
- Hamid, R.; Najme, Y.; Hamidreza, G.H.Y.; Mojtaba, T.; Majid, M. Spinach growth regulation due to interactive salinity, water, and nitrogen stresses. J. Plant Growth Regulation 2021, 41, 1654–1671. [Google Scholar]
- Zhao, D.; Oosterhuis, D.M. Dynamics of non-structural carbohydrates in developing leaves, bracts and floral buds of cotton. Environ. Exp. Bot. 2000, 43, 185–195. [Google Scholar] [CrossRef]
- Yang, J.R.; Yang, K.; Lv, C.H.; Wang, Y.J. Effects of Moderate Water Deficit on the Accumulation and Translocation of Stem Non-Structural Carbohydrates, Yield and Yield Components in a Sink-Limited Rice Variety Under Elevated CO2 Concentration. J. Plant Growth Regul. 2023, 42, 4350–4359. [Google Scholar] [CrossRef]
- Yang, J.C.; Zhang, J.H. Grain filling of cereals under soil drying. New Phytol. 2006, 169, 223–236. [Google Scholar] [CrossRef]
- Liu, J.; Wu, N.; Wang, H.; Sun, J.; Peng, B.; Jiang, P.; Bai, E. Nitrogen addition affects chemical compositions of plant tissues, litter and soil organic matter. Ecology 2016, 97, 1796–1806. [Google Scholar] [CrossRef]
- Guo, X.; Peng, C.; Li, T.; Huang, J.; Song, H.; Zhu, Q.; Wang, M. The effects of drought and re-watering on non-structural carbohydrates of Pinus tabulaeformis seedlings. Biology 2021, 10, 281. [Google Scholar] [CrossRef]
- Hoch, G. Cell wall hemicelluloses as mobile carbon stores in nonreproductive plant tissues. Funct. Ecol. 2007, 21, 823–834. [Google Scholar] [CrossRef]
- Serena, M.; Schiavon, M.; Sallenave, R.; Leinauer, B. Nitrogen fertilization of warm-season turfgrasses irrigated from varying irrigation systems: 2. Carbohydrate and protein content. J. Agron. Crop Sci. 2018, 204, 265–273. [Google Scholar] [CrossRef]
- Bazargani, M.M.; Hajirezaei, M.R.; Salekdeh, G.H.; Bushehri, A.A.S.; Falahati-Anbaran, M.; Moradi, F.; Naghavi, M.R.; Ehdaie, B. A view on the role of metabolites in enhanced stem reserves remobilization in wheat under drought during grain filling. Aust. J. Crop Sci. 2012, 6, 1613–1623. [Google Scholar]
- Xia, G.H.; Cheng, L.L. Foliar urea application in the fall affects both nitrogen and carbon storage in young ‘Concord’ grapevines grown under a wide range of nitrogen supply. HortScience 2004, 39, 827–828. [Google Scholar] [CrossRef]
- Dayer, S.; Prieto, J.A.; Galat, E.; Peña, J.P. Leaf carbohydrate metabolism in M albec grapevines: Combined effects of regulated deficit irrigation and crop load. Aust. J. Grape Wine Res. 2016, 22, 115–123. [Google Scholar] [CrossRef]
- Pan, J.F.; Cui, K.H.; Wei, D.; Huang, J.L.; Xing, J.; Nie, L. Relationships of non-structural carbohydrates accumulation and translocation with yield formation in rice recombinant inbred lines under two nitrogen levels. Physiol. Plant. 2011, 141, 321–331. [Google Scholar] [CrossRef]
- Gibson, S.I. Plant sugar-response pathways. Part of a complex regulatory web. Plant Physiol. 2000, 124, 1532–1539. [Google Scholar] [CrossRef]
- Li, P.; Zhou, H.M.; Feng, Z.Z. Ozone pollution, nitrogen addition, and drought stress interact to affect non-structural carbohydrates in the leaves and fine roots of poplar. Environ. Sci. 2021, 42, 1004–1012. [Google Scholar]
- Hoch, G.; Körner, C. The carbon charging of pines at the climatic treeline: A global comparison. Oecologia 2003, 135, 10–21. [Google Scholar] [CrossRef]
- Solfjeld, I.; Johnsen, Ø. The influence of root-zone temperature on growth of Betula pendula Roth. Trees 2006, 20, 320–328. [Google Scholar] [CrossRef]
- Okamura, M.; Arai-Sanoh, Y.; Yoshida, H.; Mukouyama, T.; Adachi, S.; Yabe, S.; Nakagawa, H.; Tsutsumi, K.; Taniguchi, Y.; Kobayashi, N.; et al. Characterization of high-yielding rice cultivars with different grain-filling properties to clarify limiting factors for improving grain yield. Field Crops Res. 2018, 219, 139–147. [Google Scholar] [CrossRef]
- Pan, J.; Cui, K.H.; Xiang, J.; Wei, D.; Wang, K.; Huang, J.L.; Nie, L.X. Characteristics of non-structural carbohydrate accumulation and translocation in rice genotypes with various sink-capacity. J. Huazhong Agric. Univ. 2015, 34, 9–15. [Google Scholar]
- Körner, C. Significance of temperature in plant life. In Plant Growth and Climate Change; Blackwell Publishing Ltd.: Oxford, UK, 2006; pp. 48–69. [Google Scholar]
- Horie, T.; Shiraiwa, T.; Homma, K.; Katsura, K.; Maeda, Y.; Yoshida, H. Can yields of lowland rice resumes the increases that showed in the 1980s. In Proceedings of the International Crop Science Congress, Queensland, Australia, 26 Septemper–1 October 2004; Volume 8, pp. 259–274. [Google Scholar]
- Fu, J.; Huang, Z.H.; Wang, Z.Q.; Yang, J.C.; Zhang, J.H. Preanthesis non-structural carbohydrate reserve in the stem enhances the sink strength of inferior spikelets during grain filling of rice. Field Crops Res. 2011, 123, 170–182. [Google Scholar] [CrossRef]
- Ho, L.C. Metabolism and compartmentation of imported sugars in sink organs in relation to sink strength. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1988, 39, 355–378. [Google Scholar] [CrossRef]
- Xue, H.; Han, Y.; Li, Y.; Wang, G.; Feng, L.; Fan, Z.; Du, W.; Yang, B.; Cao, C.; Mao, S. Spatial distribution of light interception by different plant population densities and its relationship with yield. Field Crops Res. 2015, 184, 17–27. [Google Scholar] [CrossRef]
- Zhang, L.; He, X.; Liang, Z.; Zhang, W.; Zou, C.; Chen, X. Tiller development affected by nitrogen fertilization in a high-yielding wheat production system. Crop Sci. 2020, 60, 1034–1047. [Google Scholar] [CrossRef]
- Royo, C.; Abaza, M.; Blanco, R.; del Moral, L.F. Triticale grain growth and morphometry as affected by drought stress, late sowing and simulated drought stress. Funct. Plant Biol. 2000, 27, 1051–1059. [Google Scholar] [CrossRef]
- Guo, J.; Fan, J.; Xiang, Y.; Zhang, F.; Yan, S.; Zhang, X.; Zheng, J.; Hou, X.; Tang, Z.; Li, Z. Maize leaf functional responses to blending urea and slow-release nitrogen fertilizer under various drip irrigation regimes. Agric. Water Manag. 2022, 262, 107396. [Google Scholar] [CrossRef]
- Wang, Z.H.; Bian, Q.Y.; Zhang, J.Z.; Zhou, B. Optimized water and fertilizer management of mature jujube in Xinjiang arid Area using drip irrigation. Water 2018, 10, 1467. [Google Scholar] [CrossRef]
- Cui, Z.J.; Effah, Z.; Yan, B.; Gao, Y.H.; Wu, B.; Wang, Y.F.; Xu, P.; Wang, H.D.; Zhao, B.Q.; Wang, Y.Z. Water and nitrogen coupling increased the water-nitrogen use efficiency of oilseed flax. Plants 2022, 12, 51. [Google Scholar] [CrossRef]
- Huang, M.; Wang, C.; Qi, W.; Zhang, Z.; Xu, H. Modelling the integrated strategies of deficit irrigation, nitrogen fertilization, and biochar addition for winter wheat by AquaCrop based on a two-year field study. Field Crops Res. 2022, 282, 108510. [Google Scholar] [CrossRef]
- Hamani, A.K.M.; Abubakar, S.A.; Si, Z.; Kama, R.; Gao, Y.; Duan, A. Responses of grain yield and water-nitrogen dynamic of drip-irrigated winter wheat (Triticum aestivum L.) to different nitrogen fertigation and water regimes in the North China Plain. Agric. Water Manag. 2023, 288, 108494. [Google Scholar] [CrossRef]
- Lin, E.; Liu, H.G.; He, X.L.; Li, X.X.; Gong, P.; Li, L. Water-nitrogen coupling effect on drip-irrigated dense planting of dwarf jujube in an extremely arid area. Agronomy 2019, 9, 561. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, P.; Wang, L.; Ma, G.; Li, Z.; Wang, C. Interaction of nitrogen and phosphorus on wheat yield, N use efficiency and soil nitrate nitrogen distribution in the North China Plain. Int. J. Plant Prod. 2020, 14, 415–426. [Google Scholar] [CrossRef]
- Li, X.; Leng, X.; Zhang, J.; Guo, Y.; Ding, Z.; Hu, X.; Zhu, K.L. Simulation and optimization of maize growth and nitrogen utilization under degradation film mulching in arid areas of North China. Trans. Chin. Soc. Agric. Eng. 2020, 36, 113–121. [Google Scholar]
- Liu, C.X.; Rubaek, G.H.; Liu, F.L.; Andersen, M.N. Effect of partial root zone drying and deficit irrigation on nitrogen and phosphorus uptake in potato. Agric. Water Manag. 2015, 159, 66–76. [Google Scholar] [CrossRef]
- Hao, T.; Zhu, Z.; Zhang, Y.; Liu, S.; Xu, Y.; Xu, X.; Zhao, C. Effects of Drip Irrigation and Fertilization Frequency on Yield, Water and Nitrogen Use Efficiency of Medium and Strong Gluten Wheat in the Huang-Huai-Hai Plain of China. Agronomy 2023, 13, 1564. [Google Scholar] [CrossRef]
- Chen, B.; Guo, L.; Tang, J.; Li, Y.; Li, C. Comprehensive impacts of different integrated rice-animal co-culture systems on rice yield, nitrogen fertilizer partial factor productivity and nitrogen losses: A global meta-analysis. Sci. Total Environ. 2024, 915, 169994. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Stage | 2019 | 2020 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Growth Stage | Seedling | Budding | Anthesis | Kernel | Maturity | Seedling | Budding | Anthesis | Kernel | Maturity | |
| Nitrogen | N0 | 504.48 b | 1220.50 b | 2345.71 b | 4612.74 b | 4473.88 a | 371.40 b | 1049.21 c | 2327.29 c | 4697.71 c | 4643.31 b |
| N60 | 542.48 a | 1299.38 a | 2579.98 a | 5007.56 a | 4803.13 a | 443.97 a | 1154.82 b | 2867.69 b | 5179.61 a | 5049.63 a | |
| N120 | 510.99 b | 1333.75 a | 2663.09 a | 5117.65 a | 4707.42 a | 417.54 a | 1297.71 a | 3071.07 a | 5153.45 b | 5098.69 a | |
| Irrigation | I0 | 515.06 a | 1263.13 b | 2390.54 b | 4668.92 b | 4210.73 b | 413.10 ab | 1155.51 a | 2511.67 b | 4914.18 b | 4477.33 c |
| I1200 | 516.50 a | 1252.50 b | 2568.71 a | 4957.55 a | 4816.59 a | 426.63 a | 1166.18 a | 2826.38 a | 5097.24 a | 5006.27 b | |
| I1800 | 526.38 a | 1338.00 a | 2629.53 a | 5111.48 a | 4957.11 a | 393.18 b | 1180.06 a | 2928.00 a | 5019.35 ab | 5308.03 a | |
| Irrigation (I) | ns | ** | ** | ** | ** | ns | ns | ** | ns | ** | |
| Nitrogen (N) | * | ** | ** | ** | ** | ** | ** | ** | ** | ** | |
| I × N | ns | * | ns | ns | ns | ** | ns | ** | ns | ** | |
| 2019 | 2020 | |||||||
|---|---|---|---|---|---|---|---|---|
| Irrigation (I) | Nitrogen (N) | I × N | Irrigation (I) | Nitrogen (N) | I × N | |||
| SS | Pre-A | Stems | ** | ** | * | ns | ** | ns |
| Leaves | ns | ** | ns | * | * | ns | ||
| Post-A | Stems | ns | * | ns | * | ** | ns | |
| Leaves | * | ** | ns | ns | * | ns | ||
| Capsules | * | ** | ns | ** | * | ns | ||
| ST | Pre-A | Stems | ** | * | ns | ** | ** | ns |
| Leaves | ns | ** | ns | ** | ** | ns | ||
| Post-A | Stems | ** | ** | ns | ** | ** | ** | |
| Leaves | * | ** | ns | ** | ** | ns | ||
| Capsules | ** | ** | * | ** | ** | ** |
| Item | EC | GN | TKW | GY | ||
|---|---|---|---|---|---|---|
| SS | Pre-A | Stems | 0.597 ** | −0.077 | −0.112 | 0.517 ** |
| Leaves | 0.440 ** | −0.076 | −0.120 | 0.429 ** | ||
| Post-A | Stems | 0.287 * | 0.051 | −0.019 | 0.282 * | |
| Leaves | 0.523 ** | −0.071 | 0.014 | 0.411 ** | ||
| Capsules | 0.597 ** | 0.450 ** | −0.758 ** | 0.457 ** | ||
| ST | Pre-A | Stems | 0.545 ** | 0.460 ** | −0.739 ** | 0.405 ** |
| Leaves | −0.372 ** | −0.643 ** | 0.903 ** | 0.107 | ||
| Post-A | Stems | 0.583 ** | 0.325 * | −0.621 ** | 0.531 ** | |
| Leaves | −0.278 * | −0.602 ** | 0.850 ** | 0.084 | ||
| Capsules | −0.050 | −0.468 ** | 0.575 ** | 0.387 ** | ||
| 2019 | 2020 | |||||||
|---|---|---|---|---|---|---|---|---|
| Treatment | EC (Number Plant−1) | GN (Number Capsule−1) | TKW (g) | GY (kg ha−1) | EC (Number Plant−1) | GN (Number Capsule−1) | TKW (g) | GY (kg ha−1) |
| I0N0 | 10.61 c | 7.97 a | 5.56 e | 1048.33 e | 9.47 b | 7.40 a | 7.54 a | 960.57 e |
| I0N60 | 12.42 bc | 8.02 a | 6.31 abc | 1103.50 de | 10.77 ab | 7.17 ab | 7.65 a | 1121.67 bc |
| I0N120 | 12.00 c | 7.65 a | 5.99 d | 1108.67 de | 10.33 ab | 7.00 ab | 7.61 a | 1015.10 de |
| I1200N0 | 11.52 c | 8.19 a | 6.10 bcd | 1168.6 cd | 11.77 a | 6.73 ab | 7.56 a | 1078.43 cd |
| I1200N60 | 12.93 abc | 7.20 a | 6.36 a | 1246.33 ab | 12.1 a | 6.70 ab | 7.70 a | 1260.90 a |
| I1200N120 | 12.64 bc | 7.60 a | 6.06 cd | 1214.33 bc | 11.37 ab | 7.23 ab | 7.43 a | 1174.77 b |
| I1800N0 | 13.52 abc | 7.49 a | 6.34 ab | 1237.33 abc | 10.73 ab | 6.40 b | 7.36 a | 1175.67 b |
| I1800N60 | 15.92 a | 7.50 a | 6.46 a | 1291.00 a | 11.07 ab | 6.80 ab | 7.87 a | 1300.57 a |
| I1800N120 | 15.36 ab | 7.54 a | 6.41 a | 1276.33 ab | 10.40 ab | 6.70 ab | 7.86 a | 1272.23 a |
| I | ** | ns | ** | ** | * | ns | ns | ** |
| N | ns | ns | ** | * | ns | ns | ns | ** |
| I × N | ns | ns | ** | ns | ns | ns | ns | ns |
| Treatment | 2019 | Changes (%) | 2020 | Changes (%) | ||
|---|---|---|---|---|---|---|
| GY (kg ha−1) | GY (kg ha−1) | |||||
| Nitrogen | N0 | 1151.44 b | – | 1071.56 c | – | |
| N60 | 1213.61 a | 5.40 | 1227.71 a | 14.57 | ||
| N120 | 1199.78 a | – | 1154.03 b | -6.00 | ||
| Irrigation | I0 | 1086.83 c | – | 1032.44 c | – | |
| I1200 | 1209.78 b | 11.31 | 1171.37 b | 13.45 | ||
| I1800 | 1268.22 a | 4.83 | 1249.49 a | 6.67 | ||
| Increase in percentage from I0 to I1800 | 16.69 | 21.02 | ||||
| Treatment | 2019 | 2020 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| GY (kg ha−1) | Yield Increase Rate (%) | Contribution to Yield Increase (%) | GY (kg ha−1) | Yield Increase Rate (%) | Contribution to Yield Increase (%) | |||||
| N | I | I × N | N | I | I × N | |||||
| I0N0 | 1048.33 d | – | – | – | – | 960.57 d | – | – | – | – |
| I0N60 | 1103.50 cd | 5.63 | 100.00 | – | – | 1121.67 bc | 16.77 | 100.00 | – | – |
| I0N120 | 1108.67 cd | 5.76 | 100.00 | – | – | 1015.10 d | 5.68 | 100.00 | – | – |
| I1200N0 | 1168.67 bc | 11.48 | – | 100.00 | – | 1078.43 cd | 12.27 | – | 100.00 | – |
| I1200N60 | 1246.33 ab | 18.89 | 35.18 | 68.52 | -3.70 | 1260.90 a | 31.27 | 54.11 | 39.70 | 6.20 |
| I1200N120 | 1214.33 ab | 15.84 | 24.67 | 60.17 | 15.17 | 1174.77 b | 22.30 | 40.06 | 70.54 | -10.60 |
| I1800N0 | 1237.33 ab | 18.03 | – | 100.00 | – | 1175.67 b | 22.39 | – | 100.00 | – |
| I1800N60 | 1291.00 a | 23.15 | 18.74 | 73.40 | 7.87 | 1300.57 a | 35.40 | 30.01 | 45.05 | 24.93 |
| I1800N120 | 1276.33 a | 21.75 | 14.49 | 69.53 | 15.98 | 1272.23 a | 32.45 | 25.31 | 78.06 | −3.37 |
| 2019 | 2020 | |||||||
|---|---|---|---|---|---|---|---|---|
| ETa (mm) | WUE (kg ha−1 mm−1) | NPFP (kg kg−1) | WIUE (kg ha−1 mm−1) | ETa (mm) | WUE (kg ha−1 mm−1) | NPFP (kg kg−1) | WIUE (kg ha−1 mm−1) | |
| I0N0 | 266.42 d | 3.94 a | — | — | 298.72 d | 3.24 b | — | — |
| I0N60 | 284.60 d | 3.88 a | 18.39 b | — | 302.78 d | 3.73 a | 18.69 b | — |
| I0N120 | 273.15 d | 4.06 a | 9.24 d | — | 296.48 d | 3.44 ab | 8.46 e | — |
| I1200N0 | 318.57 c | 3.67 ab | — | 9.74 a | 419.04 c | 2.59 c | — | 8.99 c |
| I1200N60 | 336.48 c | 3.71 ab | 20.77 a | 10.39 a | 460.79 bc | 2.75 c | 21.02 a | 10.51 a |
| I1200N120 | 328.00 c | 3.71 ab | 10.12 c | 10.12 a | 442.58 bc | 2.68 c | 9.79 d | 9.79 b |
| I1800N0 | 360.57 b | 3.44 b | — | 6.87 b | 487.01 ab | 2.42 c | 6.53 e | |
| I1800N60 | 375.07 ab | 3.44 b | 21.52 a | 7.17 b | 528.13 a | 2.47 c | 21.68 a | 7.23 d |
| I1800N120 | 381.11 a | 3.35 b | 10.64 c | 7.09 b | 503.49 ab | 2.54 c | 10.60 c | 7.07 d |
| Irrigation (I) | ** | ** | ** | ** | ** | ** | ** | ** |
| Nitrogen (N) | * | ns | ** | ns | ns | ns | ** | ** |
| I × N | ns | ns | * | ns | ns | ns | * | ns |
| Year | pH | Sulfate | Chloride | Sulfide | Nitrate | Nitrite | Ammonia Nitrogen | Sodium |
|---|---|---|---|---|---|---|---|---|
| 2019 | 7.82 | 486.00 | 419.00 | 0.005 | 19.05 | 0.005 | 0.04 | 180.50 |
| 2020 | 8.21 | 392.00 | 330.00 | 0.005 | 19.6 | 0.005 | 0.04 | 174.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Zhao, B.; Gao, Y.; Yan, B.; Wu, B.; Cui, Z.; Wang, Y.; Wen, M.; Ma, X. A Plant Strategy: Irrigation, Nitrogen Fertilization, and Climatic Conditions Regulated the Carbon Allocation and Yield of Oilseed Flax in Semi-Arid Area. Plants 2024, 13, 2553. https://doi.org/10.3390/plants13182553
Wang H, Zhao B, Gao Y, Yan B, Wu B, Cui Z, Wang Y, Wen M, Ma X. A Plant Strategy: Irrigation, Nitrogen Fertilization, and Climatic Conditions Regulated the Carbon Allocation and Yield of Oilseed Flax in Semi-Arid Area. Plants. 2024; 13(18):2553. https://doi.org/10.3390/plants13182553
Chicago/Turabian StyleWang, Haidi, Bangqing Zhao, Yuhong Gao, Bin Yan, Bing Wu, Zhengjun Cui, Yifan Wang, Ming Wen, and Xingkang Ma. 2024. "A Plant Strategy: Irrigation, Nitrogen Fertilization, and Climatic Conditions Regulated the Carbon Allocation and Yield of Oilseed Flax in Semi-Arid Area" Plants 13, no. 18: 2553. https://doi.org/10.3390/plants13182553
APA StyleWang, H., Zhao, B., Gao, Y., Yan, B., Wu, B., Cui, Z., Wang, Y., Wen, M., & Ma, X. (2024). A Plant Strategy: Irrigation, Nitrogen Fertilization, and Climatic Conditions Regulated the Carbon Allocation and Yield of Oilseed Flax in Semi-Arid Area. Plants, 13(18), 2553. https://doi.org/10.3390/plants13182553