
Shading and Water Addition Alleviate the Elemental Limitations of the Early Restoration Community in a Stressful Environment

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Concentrations and Ratios of Nutrient Elements in the Communities
2.2. Effects of Shading and Water Addition on Stoichiometric Traits in the Communities
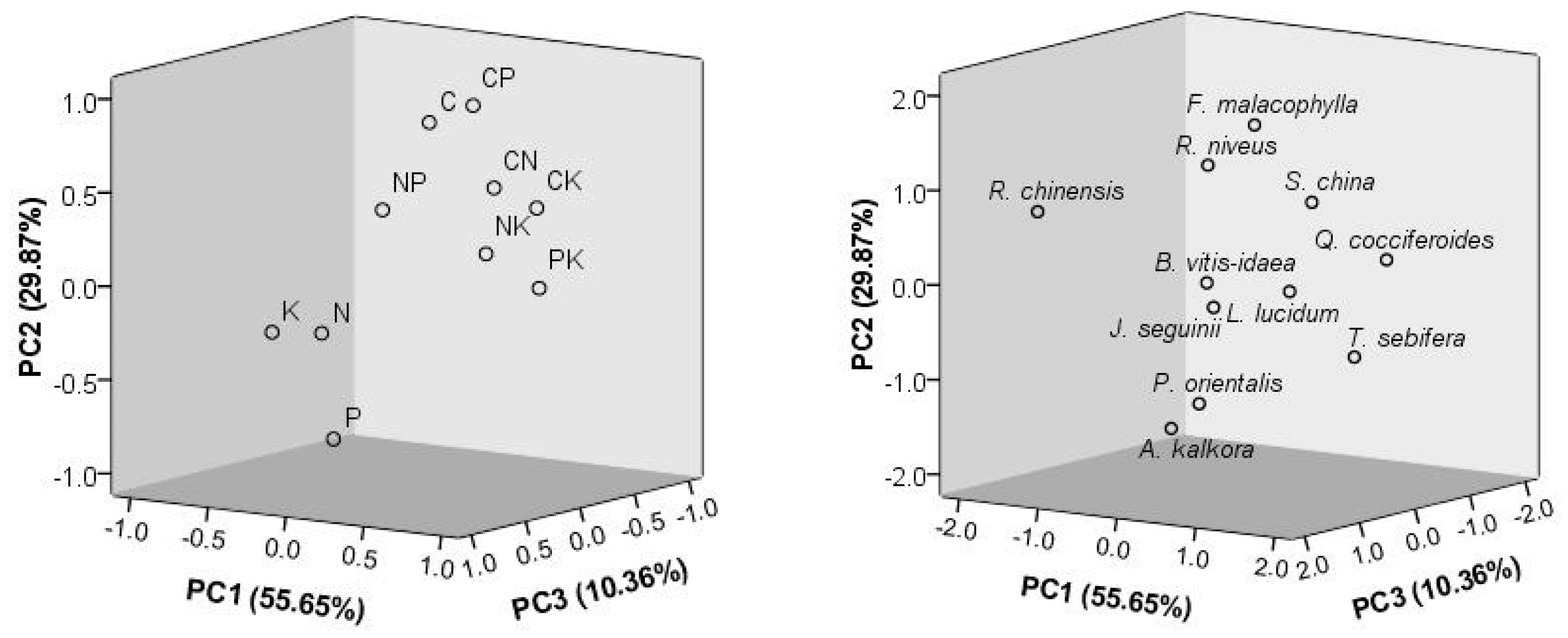
2.3. Species-Specific Variation and Comparisons with the Control and Natural Forest
3. Discussion
3.1. Elemental Characteristics and Their Responses to Shading and Water Addition
3.2. Interactive Effects of Shading and Water Addition on Leaf Stoichiometry
3.3. Limitation of Nutrient Elements and Implications for Vegetation Restoration
4. Materials and Methods
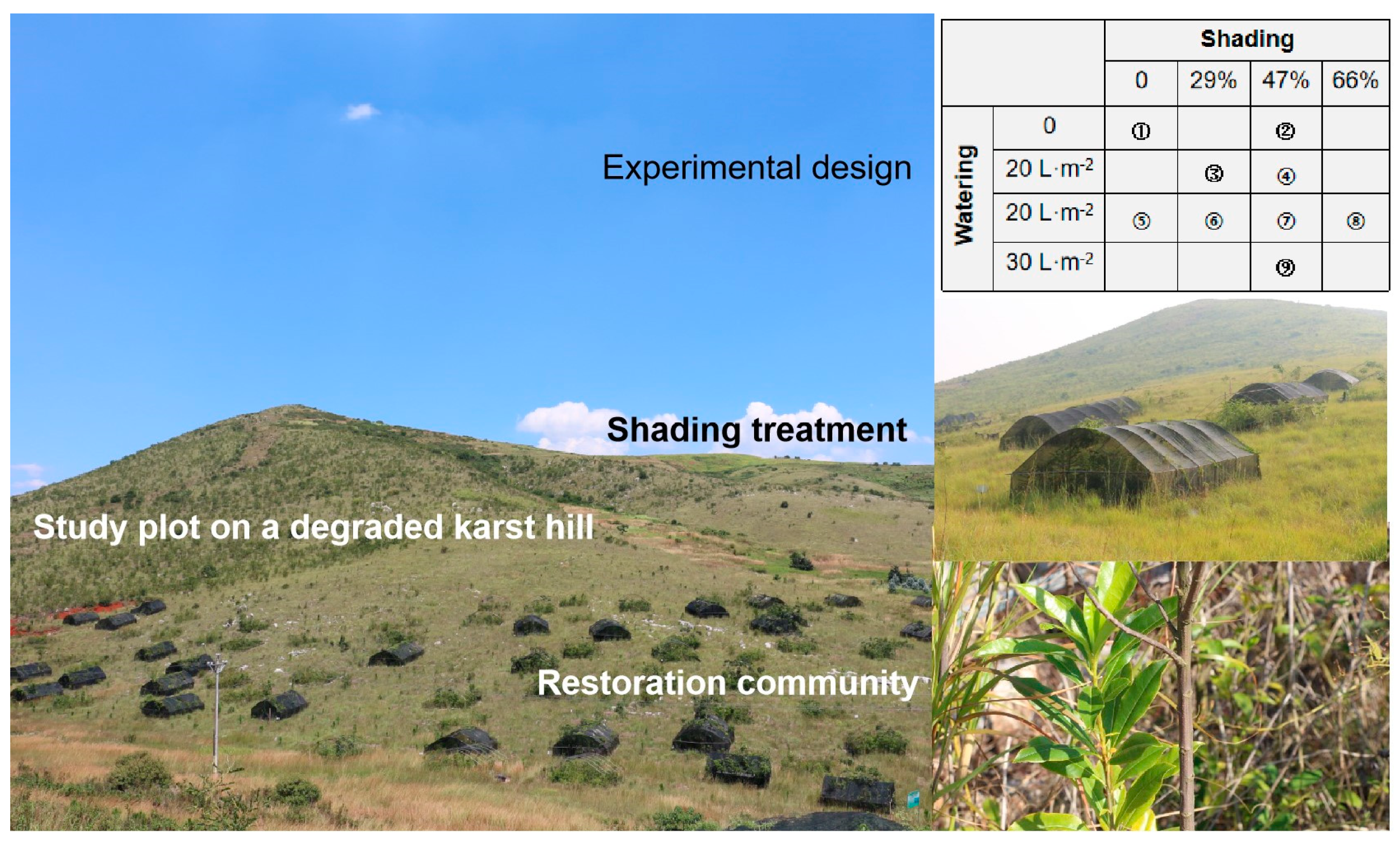
4.1. Study Site
4.2. Experimental Design
4.3. Sampling and Measurement
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoekstra, J.M.; Boucher, T.M.; Ricketts, T.H.; Roberts, C. Confronting a biome crisis: Global disparities of habitat loss and protection. Ecol. Lett. 2005, 8, 23–29. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment 2020: Main Report; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Matricardi, E.A.T.; Skole, D.L.; Costa, O.B.; Pedlowski, M.A.; Samek, J.H.; Miguel, E.P. Long-term forest degradation surpasses deforestation in the Brazilian Amazon. Science 2020, 369, 1378–1382. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.J.; Liu, Q.M.; Zhang, D.F. Karst rocky desertification in southwestern China: Geomorphology, landuse, impact and rehabilitation. Land Degrad. Dev. 2004, 15, 115–121. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Why seedlings grow: Influence of plant attributes. New Forest. 2018, 49, 1–34. [Google Scholar] [CrossRef]
- Zhao, G.J.; Shen, Y.X.; Liu, W.Y.; Tan, B.L.; Li, Z.J.; Wang, Q.H. Quantifying the effect of shading and watering on seed germination in translocated forest topsoil at a subtropical karst of China. For. Ecol. Manag. 2020, 459, 117811. [Google Scholar] [CrossRef]
- Lin, J.; Harcombe, P.A.; Fulton, M.R.; Hall, R.W. Sapling growth and survivorship as a function of light in a mesic forest of southeast Texas, USA. Oecologia 2002, 132, 428–435. [Google Scholar] [CrossRef]
- Semchenko, M.; Lepik, M.; Götzenberger, L.; Zobel, K. Positive effect of shade on plant growth: Amelioration of stress or active regulation of growth rate? J. Ecol. 2012, 100, 459–466. [Google Scholar] [CrossRef]
- Held, I.M.; Soden, B.J. Robust responses of the hydrological cycle to global warming. J. Clim. 2006, 19, 5686–5699. [Google Scholar] [CrossRef]
- Xiong, J.L.; Dong, L.W.; Lu, J.L.; Hu, W.G.; Gong, H.Y.; Xie, S.B.; Zhao, D.M.; Zhang, Y.H.; Wang, X.T.; Deng, Y.; et al. Variation in plant carbon, nitrogen and phosphorus contents across the drylands of China. Funct. Ecol. 2022, 36, 174–186. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, Ü.; Ntagkas, N.; Siebenkäs, A.; Mäenpää, M.; Matsubara, S.; Pons, T.L. A meta-analysis of plant responses to light intensity for 70 traits ranging from molecules to whole plant performance. New Phytol. 2019, 223, 1073–1105. [Google Scholar] [CrossRef]
- Kamiyama, C.; Oikawa, S.; Hikosaka, K. Seasonal change in light partitioning among coexisting species of different functional groups along elevation gradient in subalpine moorlands. New Phytol. 2014, 204, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Hachemi, A.; Ali, O.S.; Belghazi, T.; Lahrouni, A.; Mercht, S.E.; Hassan, C.E.; Messoussi, S.E. Effect of hydric and light stress on biomass, nutrient uptake and enzymatic antioxidants of Argania spinosa seedlings. Arch. Biol. Sci. 2021, 73, 145–153. [Google Scholar] [CrossRef]
- Poorter, L. Growth responses of 15 rain-forest tree species to a light gradient: The relative importance of morphological and physiological traits. Funct. Ecol. 1999, 13, 396–410. [Google Scholar] [CrossRef]
- Lin, J.; Zhang, R.; Hu, Y.; Song, Y.; Hänninen, H.; Wu, J. Interactive effects of drought and shading on Torreya grandis seedlings: Physiological and growth responses. Trees 2019, 33, 951–961. [Google Scholar] [CrossRef]
- Sack, L. Responses of temperate woody seedlings to shade and drought: Do trade-offs limit potential niche differentiation? Oikos 2004, 107, 110–127. [Google Scholar] [CrossRef]
- Quero, J.L.; Villar, R.; Marañón, T.; Zamora, R. Interactions of drought and shade effects on seedlings of four quercus species: Physiological and structural leaf responses. New Phytol. 2006, 170, 819–834. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, T.; Ma, W.; Tian, C.; Sha, Z.; Li, J. Morphological and physiological response of Acer catalpifolium rehd. seedlings to water and light stresses. Glob. Ecol. Conserv. 2019, 19, e00660. [Google Scholar] [CrossRef]
- Tang, Z.Y.; Xu, W.T.; Zhou, G.Y.; Bai, Y.F.; Li, J.X.; Tang, X.L.; Chen, D.M.; Liu, Q.; Ma, W.H.; Xiong, G.M.; et al. Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 2018, 115, 4033–4038. [Google Scholar] [CrossRef]
- Sardans, J.; Janssens, I.A.; Ciais, P.; Obersteiner, M.; Peñuelas, J. Recent advances and future research in ecological stoichiometry. Perspect. Plant Ecol. 2021, 50, 125611. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, J.; Pan, F.; Li, D.; Chen, H.; Wang, K. Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in southwest China. Plant Soil 2015, 391, 77–91. [Google Scholar] [CrossRef]
- Shen, Y.X.; Yu, Y.; Lucas-Borja, M.E.; Chen, F.J.; Chen, Q.Q.; Tang, Y.Y. Change of soil K, N and P following forest restoration in rock outcrop rich karst area. Catena 2020, 186, 104395. [Google Scholar] [CrossRef]
- Güsewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Peñuelas, J.; Coll, M.; Vayreda, J.; Rivas-Ubach, A. Stoichiometry of potassium is largely determined by water availability and growth in Catalonian forests. Funct. Ecol. 2012, 26, 1077–1089. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium control of plant functions: Ecological and agricultural implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, A.E.; Llano-Sotelo, J.M.; Machado-Encinas, L.I.; López-Piña, J.E.; Romo-Leon, J.R.; Sardans, J.; Peñuelas, J. Foliar C, N, and P stoichiometry characterize successful plant ecological strategies in the Sonoran Desert. Plant Ecol. 2018, 219, 775–788. [Google Scholar] [CrossRef]
- Koerselman, W.; Meuleman, A.F.M. The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Tessier, J.T.; Raynal, D.J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. J. Appl. Ecol. 2003, 40, 523–534. [Google Scholar] [CrossRef]
- Meier, I.C.; Leuschner, C. Nutrient dynamics along a precipitation gradient in European beech forests. Biogeochemistry 2014, 120, 51–69. [Google Scholar] [CrossRef]
- He, P.; Fontana, S.; Sardans, J.; Peñuelas, J.; Gessler, A.; Schaub, M.; Rigling, A.; Li, H.; Jiang, Y.; Li, M.H. The biogeochemical niche shifts of pinus sylvestris var. mongolica along an environmental gradient. Environ. Exp. Bot. 2019, 167, 103825. [Google Scholar] [CrossRef]
- Umair, M.; Sun, N.; Du, H.; Chen, K.; Tao, H.; Yuan, J.; Abbasi, A.M.; Liu, C. Differential stoichiometric responses of shrubs and grasses to increased precipitation in a degraded karst ecosystem in southwestern china. Sci. Total Environ. 2020, 700, 134421. [Google Scholar] [CrossRef]
- Fernández-Martínez, M. From atoms to ecosystems: Elementome diversity meets ecosystem functioning. New Phytol. 2022, 234, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Yan, Z.B.; Niklas, K.J.; Han, W.X.; Kattge, J.; Reich, P.B.; Luo, Y.K.; Chen, Y.H.; Tang, Z.Y.; Hu, H.F.; et al. Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent. Natl. Sci. Rev. 2018, 5, 728–739. [Google Scholar] [CrossRef]
- Pan, F.J.; Zhang, W.; Liu, S.J.; Li, D.J.; Wang, K.L. Leaf N:P stoichiometry across plant functional groups in the karst region of southwestern China. Trees 2015, 29, 883–892. [Google Scholar] [CrossRef]
- Han, W.X.; Fang, J.Y.; Guo, D.L.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef]
- Zhang, C.; Tian, H.; Liu, J.; Wang, S.; Liu, M.; Pan, S.; Shi, X. Pools and distributions of soil phosphorus in China. Glob. Biogeochem. Cycles 2005, 19, GB1020. [Google Scholar] [CrossRef]
- Tian, D.; Reich, P.B.; Chen, H.Y.H.; Xiang, Y.; Luo, Y.; Shen, Y.; Meng, C.; Han, W.; Niu, S. Global changes alter plant multi-element stoichiometric coupling. New Phytol. 2019, 221, 807–817. [Google Scholar] [CrossRef]
- Wang, Z.; Silva, L.C.R.; Sun, G.; Luo, P.; Mou, C.X.; Horwath, W.R. Quantifying the impact of drought on soil-plant interactions: A seasonal analysis of biotic and abiotic controls of carbon and nutrient dynamics in high-altitudinal grasslands. Plant Soil 2015, 389, 59–71. [Google Scholar] [CrossRef]
- He, M.; Dijkstra, F.A. Drought effect on plant nitrogen and phosphorus: A meta-analysis. New Phytol. 2014, 204, 924–931. [Google Scholar] [CrossRef]
- Niu, D.; Zhang, C.; Ma, P.; Fu, H.; Elser, J.J. Responses of leaf C:N:P stoichiometry to water supply in the desert shrub Zygophyllum xanthoxylum. Plant Biol. 2019, 21, 82–88. [Google Scholar] [CrossRef]
- Zhang, Q.; Luo, D.; Yang, L.; Xie, J.; Yang, Z.; Zhou, J.; Li, X.; Xiong, D.; Chen, Y.; Yang, Y. Variations in rainfall affect the responses of foliar chemical properties of Cunninghamia lanceolata seedlings to soil warming. Front. Plant Sci. 2021, 12, 705861. [Google Scholar] [CrossRef]
- Dynarski, K.A.; Soper, F.M.; Reed, S.C.; Wieder, W.R.; Cleveland, C.C. Patterns and controls of foliar nutrient stoichiometry and flexibility across United States forests. Ecology 2023, 104, e3909. [Google Scholar] [CrossRef] [PubMed]
- Callaway, R.M. Positive Interactions and Interdependence in Plant Communities; Springer: Dordrecht, The Netherlands, 2007; pp. 23–48. [Google Scholar]
- Pájaro-Esquivia, Y.S.; Domínguez-Haydar, Y.; Tinoco-Ojanguren, C. Intraspecific variation in morpho-functional traits and plastic response to water and light in seedlings of Aspidosperma polyneuron (Apocynaceae). Flora 2021, 282, 151903. [Google Scholar] [CrossRef]
- Elser, J.J.; Dobberfuhl, D.R.; MacKay, N.A.; Schampel, J.H. Organism size, life history, and N:P stoichiometry. Bioscience 1996, 46, 674–684. [Google Scholar] [CrossRef]
- Ashton, M.S.; Singhakumara, B.M.P.; Gamage, H.K. Interaction between light and drought affect performance of Asian tropical tree species that have differing topographic affinities. For. Ecol. Manag. 2006, 221, 42–51. [Google Scholar] [CrossRef]
- Rentería, L.Y.; Jaramillo, V.J. Rainfall drives leaf traits and leaf nutrient resorption in a tropical dry forest in Mexico. Oecologia 2011, 165, 201–211. [Google Scholar] [CrossRef]
- Liu, Q.; Huang, Z.; Wang, Z.; Chen, Y.; Wen, Z.; Liu, B.; Tigabu, M. Responses of leaf morphology, NSCs contents and C:N:P stoichiometry of Cunninghamia lanceolata and Schima superba to shading. BMC Plant Biol. 2020, 20, 354. [Google Scholar] [CrossRef]
- Zhang, Q.Z.; Zhang, J.Y.; Shi, Z.L.; Kang, B.Y.; Tu, H.K.; Zhu, J.Y.; Li, H.Y. Nitrogen addition and drought affect nitrogen uptake patterns and biomass production of four urban greening tree species in North China. Sci. Total Environ. 2023, 893, 164893. [Google Scholar] [CrossRef]
- Chen, F.; Zhao, G.; Shen, Y.; Li, Z.; Tan, B.; Zhu, H.; Wang, Q.; Fu, X. Shading gradients drive differential responses of leaf traits in an early community germinated by forest topsoil. Diversity 2022, 14, 600. [Google Scholar] [CrossRef]
- Huang, J.B.; Liu, W.Y.; Li, S.; Song, L.; Lu, H.Z.; Shi, X.M.; Chen, X.; Hu, T.; Liu, S.; Liu, T. Ecological stoichiometry of the epiphyte community in a subtropical forest canopy. Ecol. Evol. 2019, 9, 14394–14406. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T. C (g·kg−1) | T. N (g·kg−1) | T. P (g·kg−1) | T. K (g·kg−1) | C:N | C:P | C:K | N:P | N:K | P:K | |
|---|---|---|---|---|---|---|---|---|---|---|
| Community mean, arithmetic | ||||||||||
| Sha × sha | 2.115 ** | −0.883 | −1.649 ** | −1.554 * | 1.007 | 2.214 ** | 1.580* | 1.836 ** | 0.984 | −0.268 |
| Shading | −1.873 * | 0.935 | 1.703 ** | 1.827 ** | −0.939 | −2.273 ** | −1.924 ** | −1.869 ** | −1.375 * | 0.080 |
| Wat × wat | 0.864 | −1.317 * | 1.472 ** | −0.426 | 1.568 | −1.200 ** | 0.886 | −2.077 ** | 0.924 | 1.942 ** |
| Watering | −0.324 | 1.114 | −0.818 | −0.223 | −1.310 | 0.726 | −0.450 | 1.396 ** | −0.393 | −1.034 * |
| Sha × wat | −0.582 | 0.712 | −0.065 | 0.314 | −0.563 | 0.065 | −0.107 | 0.405 | 0.104 | −0.216 |
| F | 2.414 | 3.932 | 10.617 | 3.439 | 1.887 | 11.542 | 3.628 | 18.431 | 3.074 | 10.306 |
| p | 0.098 | 0.024 | 0.000 | 0.037 | 0.171 | 0.000 | 0.031 | 0.000 | 0.051 | 0.001 |
| Community mean, weighted by abundance | ||||||||||
| Sha × sha | 0.409 | −0.538 | −1.373 ** | −1.263 * | 0.264 | 1.768 ** | 1.294 * | 1.720 ** | 0.875 | 0.039 |
| Shading | 0.976 | −0.670 | 1.223 * | 1.996 ** | 1.093 | −1.405 * | −1.924 ** | −2.133 ** | −1.758 * | −0.511 |
| Wat × wat | 0.769 | −0.439 | 1.811 ** | 0.037 | 0.690 | −1.550 * | 0.326 | −2.072 ** | 0.491 | 1.472 ** |
| Watering | 0.085 | 0.235 | −1.141 * | −0.339 | −0.320 | 1.098 * | 0.034 | 1.338 ** | −0.169 | −0.701 |
| Sha × wat | −1.338 * | 1.085 | −0.06 | −0.142 | −1.128 * | −0.050 | 0.046 | 0.622 | 0.267 | −0.084 |
| F | 3.462 | 2.872 | 8.911 | 5.961 | 4.508 | 6.400 | 4.688 | 15.390 | 9.144 | 6.425 |
| p | 0.036 | 0.062 | 0.001 | 0.005 | 0.015 | 0.004 | 0.013 | 0.000 | 0.001 | 0.004 |
| Community mean, weighted by leaf biomass | ||||||||||
| Sha × sha | −0.016 | 0.091 | −1.602 * | −0.976 | −0.055 | 1.696 * | 1.076 | 1.954 ** | 1.007 * | 0.013 |
| Shading | 1.189 * | −0.727 | 1.379 * | 1.224 * | 0.988 | −1.260 * | −1.236 * | −2.179 ** | −1.451 ** | −0.382 |
| Wat × wat | 1.203 * | −1.227 | 0.745 | −0.409 | 1.495 * | −0.451 | 0.479 | −1.580 ** | −0.152 | 0.786 |
| Watering | −0.174 | 1.652 * | −0.059 | 0.162 | −1.564 ** | −0.101 | −0.054 | 1.072 * | 0.619 | −0.013 |
| Sha × wat | −1.007 * | 0.091 | 0.000 | 0.665 | −0.463 | −0.125 | −0.822 | 0.165 | −0.624 | −0.700 |
| F | 9.853 | 3.852 | 5.426 | 4.297 | 5.203 | 4.345 | 4.047 | 12.929 | 8.816 | 3.913 |
| p | 0.001 | 0.026 | 0.008 | 0.018 | 0.009 | 0.017 | 0.022 | 0.000 | 0.001 | 0.025 |
| No. | Species | Family | Genus | Growth Form | Leaf Habit |
|---|---|---|---|---|---|
| 1 | Albizia kalkora (Roxb.) Prain | Fabaceae | Albizia | Tree | Deciduous |
| 2 | Breynia vitis-idaea (Burm. F.) C. E. C. Fischer | Phyllanthaceae | Breynia | Shrub | Deciduous |
| 3 | Fraxinus malacophylla Hemsl. | Oleaceae | Fraxinus | Tree | Deciduous |
| 4 | Jasminum seguinii Lévl. | Oleaceae | Jasminum | Shrub | Evergreen |
| 5 | Ligustrum lucidum Ait. | Oleaceae | Ligustrum | Tree | Evergreen |
| 6 | Paliurus orientalis (Franch.) Hemsl. | Rhamnaceae | Paliurus | Tree | Deciduous |
| 7 | Quercus cocciferoides Hand.-Mazz. | Fagaceae | Quercus | Tree | Semievergreen |
| 8 | Rhus chinensis Mill. | Anacardiaceae | Rhus | Tree | Deciduous |
| 9 | Rubus niveus Thunb. | Rosaceae | Rubus | Shrub | Deciduous |
| 10 | Smilax china L. | Smilacaceae | Smilax | Liana | Evergreen |
| 11 | Triadica sebifera (Linnaeus) Small | Euphorbiaceae | Triadica | Tree | Deciduous |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.; Zhao, G.; Shen, Y.; Zhu, H.; Li, Z.; Tan, B. Shading and Water Addition Alleviate the Elemental Limitations of the Early Restoration Community in a Stressful Environment. Plants 2024, 13, 2626. https://doi.org/10.3390/plants13182626
Chen F, Zhao G, Shen Y, Zhu H, Li Z, Tan B. Shading and Water Addition Alleviate the Elemental Limitations of the Early Restoration Community in a Stressful Environment. Plants. 2024; 13(18):2626. https://doi.org/10.3390/plants13182626
Chicago/Turabian StyleChen, Fajun, Gaojuan Zhao, Youxin Shen, Hong Zhu, Zhenjiang Li, and Beilin Tan. 2024. "Shading and Water Addition Alleviate the Elemental Limitations of the Early Restoration Community in a Stressful Environment" Plants 13, no. 18: 2626. https://doi.org/10.3390/plants13182626
APA StyleChen, F., Zhao, G., Shen, Y., Zhu, H., Li, Z., & Tan, B. (2024). Shading and Water Addition Alleviate the Elemental Limitations of the Early Restoration Community in a Stressful Environment. Plants, 13(18), 2626. https://doi.org/10.3390/plants13182626