
Response of Native and Non-Native Subarctic Plant Species to Continuous Illumination by Natural and Artificial Light

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Plants under Artificial Lighting
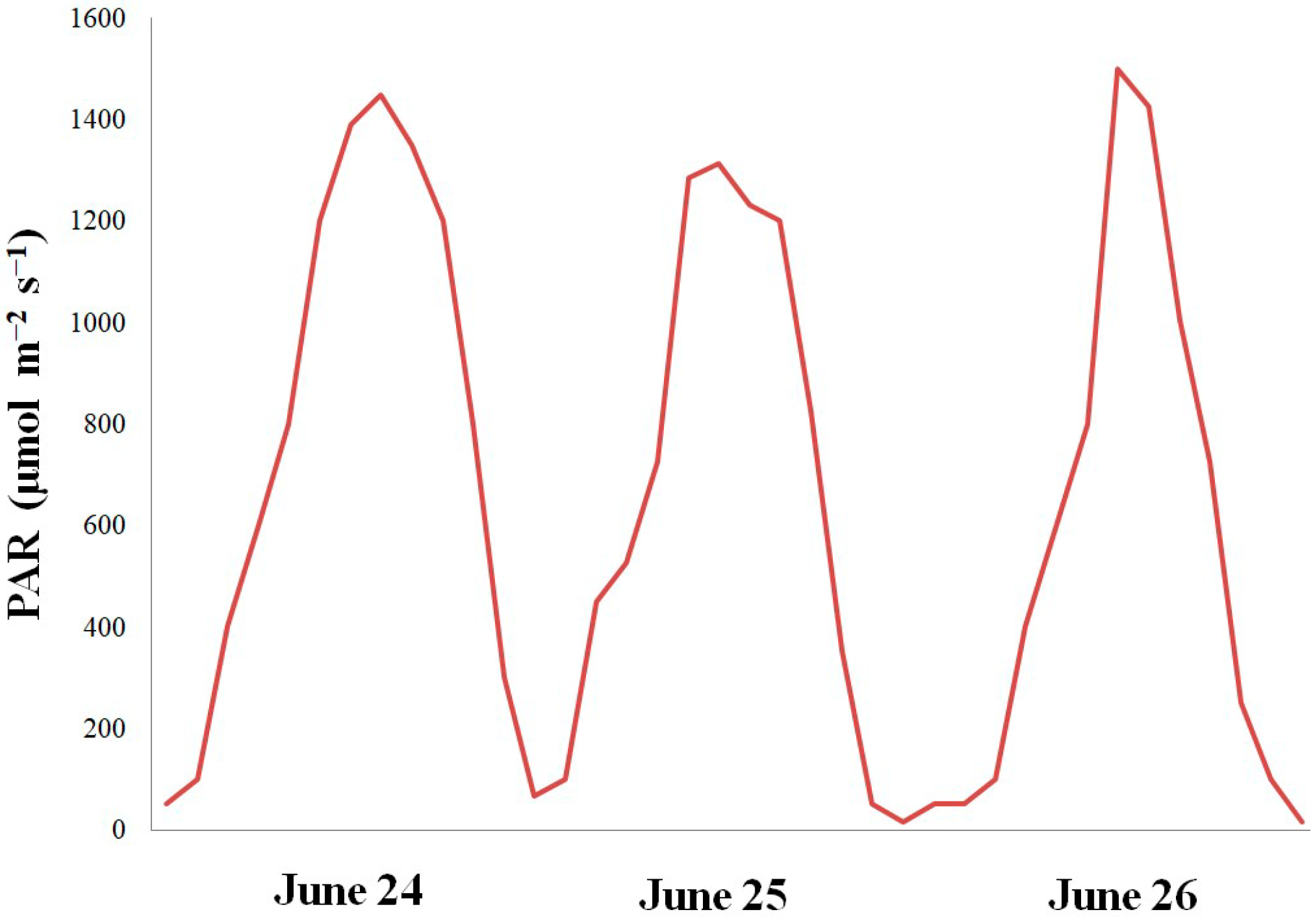
2.2. Plants under Natural Lighting
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Experimental Design
4.3. Leaf Mass Per Area
4.4. Chlorophyll Fluorescence Measurements
4.5. Stomatal Conductance Measurements
4.6. Photosynthetic Pigment Analysis
4.7. Anthocyanins and Flavonoids Content
4.8. Malondialdehyde (MDA) Content
4.9. Hydrogen Peroxide Content
4.10. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barber, J.; Andersson, B. Too much of a good thing: Light can be bad for photosynthesis. Trends Biochem. Sci. 1992, 17, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Sysoeva, M.I.; Markovskaya, E.F.; Shibaeva, T.G. Plants under continuous light: A review. Plant Stress 2010, 4, 5–17. [Google Scholar]
- Velez-Ramirez, A.I.; van Ieperen, W.; Vreugdenhil, D.; Millenaar, F.F. Plants under continuous light. Trends Plant Sci. 2011, 16, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Velez-Ramirez, A.I.; Heuvelink, E.; van Ieperen, W.; Vreugdenhil, D.; Millenaar, F. Continuous light as a way to increase greenhouse tomato production: Expected challenges. ISHS Acta Hortic. 2012, 956, 51–57. [Google Scholar] [CrossRef]
- Shibaeva, T.G.; Sherudilo, E.G.; Rubaeva, A.A.; Titov, A.F. Continuous LED lighting enhances yield and nutritional value of four genotypes of Brassicaceae microgreens. Plants 2022, 11, 176. [Google Scholar] [CrossRef]
- Lanoue, J.; St. Louis, S.; Little, C.; Hao, X. Photosynthetic adaptation strategies in peppers under continuous lighting: Insights into photosystem protection. Front Plant Sci. 2024, 15, 1372886. [Google Scholar] [CrossRef] [PubMed]
- Velez-Ramirez, A.I.; Carreño-Quintero, N.; Vreugdenhil, D.; Millenaar, F.F.; van Ieperen, W. Sucrose and Starch Content Negatively Correlates with PSII Maximum Quantum Efficiency in Tomato (Solanum lycopersicum) Exposed to Abnormal Light/Dark Cycles and Continuous Light. Plant Cell Physiol. 2017, 58, 1339–1349. [Google Scholar] [CrossRef]
- Velez-Ramirez, A.I.; Dunner-Planella, G.; Vreugdenhil, D.; Millenaar, F.F.; van Ieperen, W. On the induction of injury in tomato under continuous light: Circadian asynchrony as the main triggering factor. Funct. Plant Biol. 2017, 44, 597–611. [Google Scholar] [CrossRef]
- Marie, T.R.J.G.; Leonardos, E.D.; Lanoue, J.; Hao, X.; Micallef, B.J.; Grodzinski, B. A perspective emphasizing circadian rhythm entrainment to ensure sustainable crop production in controlled environment agriculture: Dynamic use of LED cues. Front. Sustain. Food Syst. 2022, 6, 856162. [Google Scholar] [CrossRef]
- Velez-Ramirez, A.I.; Van Ieperen, W.; Vreugdenhil, D.; van Poppel, P.M.; Heuvelink, E.; Millenaar, F.F. A single locus confer tolerance to continuous light and allows substantial yield increase in tomato. Nat. Commun. 2014, 5, 4549. [Google Scholar] [CrossRef]
- Inoue, K.; Araki, T.; Endo, M. Circadian clock during plant development. J. Plant Res. 2018, 131, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Shibaeva, T.G.; Mamaev, A.V.; Titov, A.F. Possible Physiological Mechanisms of Leaf Photodamage in Plants Grown under Continuous Lighting. Russ. J. Plant Physiol. 2023, 70, 15. [Google Scholar] [CrossRef]
- Pham, M.D.; Hwang, H.; Park, S.W.; Cui, M.; Lee, H.; Chun, C. Leaf chlorosis, epinasty, carbohydrate contents and growth of tomato show different responses to the red/blue wavelength ratio under continuous light. Plant Physiol. Biochem. 2019, 141, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Vialet-Chabrand, S.; Matthews, J.S.A.; Brendel, O.; Blatt, M.R.; Wang, Y.; Hills, A.; Griffiths, H.; Rogers, S.; Lawson, T. Modelling water use efficiency in a dynamic environment: An example using Arabidopsis thaliana. Plant Sci. 2016, 251, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Vialet-Chabrand, S.; Matthews, J.S.A.; Simkin, A.J.; Raines, C.A.; Lawson, T. Importance of fluctuations in light on plant photosynthetic acclimation. Plant Physiol. 2017, 173, 2163–2179. [Google Scholar] [CrossRef] [PubMed]
- Jänkänpää, H.J.; Mishra, Y.; Schröder, W.P.; Jansson, S. Metabolic profiling reveals metabolic shifts in Arabidopsis plants grown under different light conditions. Plant Cell Environ. 2012, 35, 1824–1836. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Fiorani, F.; Pieruschka, R.; Wojciechowski, T.; Van der Putten, W.H.; Kleyer, M.; Schurr, U.; Postma, J. Pampered inside, pestered outside? Differences and similarities between plants growing in controlled conditions and in the field. New Phytol. 2016, 212, 838–855. [Google Scholar] [CrossRef] [PubMed]
- Demers, D.A.; Gosselin, A. Growing greenhouse tomato and sweet pepper under supplemental lighting: Optimal photoperiod, negative effects of long photoperiod and their causes. Acta Hortic. 2002, 580, 83–88. [Google Scholar] [CrossRef]
- Hillman, W.S. Injury of tomato plants by continuous light and unfavorable photoperiodic cycles. Amer. J. Bot. 1956, 43, 89. [Google Scholar] [CrossRef]
- Demers, D.A.; Dorais, M.; Wien, H.C.; Gosselin, A. Effects of supplemental light duration on greenhouse tomato (Lycopersicon exculentum Mill.) plants and fruit yields. Sci. Hortic. 1998, 74, 295. [Google Scholar] [CrossRef]
- Matsuda, R.; Ozawa, N.; Fujiwara, K. Effects of continuous lighting with or without a diurnal temperature difference on photosynthetic characteristics of tomato leaves. Acta Hortic. 2012, 956, 165. [Google Scholar] [CrossRef]
- Matsuda, R.; Yamano, T.; Murakami, K.; Fujiwara, K. Effects of spectral distribution and photosynthetic photon flux density for overnight LED light irradiation on tomato seedling growth and leaf injury. Sci. Hortic. 2016, 198, 363–369. [Google Scholar] [CrossRef]
- Haque, M.S.; Kjaer, K.H.; Rosenqvist, E.; Ottosen, C.O. Continuous light increases growth, daily carbon gain, antioxidants, and alters carbohydrate metabolism in a cultivated and a wild tomato species. Front. Plant Sci. 2015, 6, 522. [Google Scholar] [CrossRef]
- Haque, M.; de Sousa, A.; Soares, C.; Kjaer, K.H.; Fidalgo, F.; Rosenqvist, E.; Ottosen, C.-O. Temperature variation under continuous light restores tomato leaf photosynthesis and maintains the diurnal pattern in stomatal conductance. Front. Plant Sci. 2017, 8, 1602. [Google Scholar] [CrossRef] [PubMed]
- Ikkonen, E.N.; Shibaeva, T.G.; Rosenqvist, E.; Ottosen, C.O. Daily temperature drop prevents inhibition of photosynthesis in tomato plants under continuous light. Photosynthetica 2015, 53, 114. [Google Scholar] [CrossRef]
- Shibaeva, T.G.; Mamaev, A.V.; Sherudilo, E.G.; Titov, A.F. The role of photosynthetic daily light integral in plant response to extended photoperiods. Russ. J. Plant Physiol. 2022, 69, 7. [Google Scholar] [CrossRef]
- Murage, E.; Watashiro, N.; Masuda, M. Leaf chlorosis and carbon metabolism of eggplant in response to continuous light and carbon dioxide. Sci. Hortic. 1996, 67, 27. [Google Scholar] [CrossRef]
- Murage, E.N.; Sato, Y.; Masuda, M. Influence of light quality, PPFD and temperature on leaf chlorosis of eggplants grown under continuous illumination. Sci. Hortic. 1997, 68, 73. [Google Scholar] [CrossRef]
- Murage, E.N.; Masuda, M. Response of pepper and eggplant to continuous light in relation to leaf chlorosis and activites of antioxidative enzymes. Sci. Hortic. 1997, 70, 269–279. [Google Scholar] [CrossRef]
- Lanoue, J.; Little, C.; Hao, X. The power of far-red light at night: Photomorphogenic, physiological, and yield response in pepper during dynamic 24 hour lighting. Front. Plant Sci. 2022, 13, 857616. [Google Scholar] [CrossRef] [PubMed]
- Wolff, S.A.; Langerud, A. Fruit yield, starch content and leaf chlorosis in cucumber exposed to continuous lighting. Eur. J. Hortic. Sci. 2006, 71, 259. [Google Scholar]
- Wheeler, R.M.; Tibbitts, T.W. Utilization of potatoes for life support systems in space. I. Cultivar-photoperiod interaction. Am. Potato J. 1986, 63, 315. [Google Scholar] [CrossRef]
- Wheeler, R.M.; Tibbitts, T.W. Growth and tuberization of potato (Solanum tuberosum L.) under continuous light. Plant Physiol. 1986, 80, 801. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Tibbitts, T.W. Physiological responses in potato plants under continuous irradiation. J. Am. Soc. Hortic. Sci. 1991, 116, 525. [Google Scholar] [CrossRef] [PubMed]
- Dorais, M.; Gosselin, A. Physiological response of greenhouse vegetable crops to supplemental lighting. Acta Hortic. 2002, 580, 59. [Google Scholar] [CrossRef]
- Matsuda, R.; Ozawa, N.; Fujiwara, K. Leaf photosynthesis, plant growth, and carbohydrate accumulation of tomato under different photoperiods and diurnal temperature differences. Sci. Hortic. 2014, 170, 150–158. [Google Scholar] [CrossRef]
- Foyer, C.H.; Neukermans, J.; Queval, G.; Noctor, G.; Harbinson, J. Photosynthetic control of electron transport and the regulation of gene expression. J. Exp. Bot. 2012, 63, 1637–1661. [Google Scholar] [CrossRef]
- Zhu, M.-D.; Chen, X.-L.; Zhu, X.-Y.; Xing, Y.-D.; Du, D.; Zhang, Y.-Y.; Lu, X.; Peng, S.-S.; He, G.-H.; Zhang, T.-Q. Identification and gene mapping of the starch accumulation and premature leaf senescence mutant ossac4 in rice. J. Integr. Agric. 2020, 19, 2150–2164. [Google Scholar] [CrossRef]
- Pammenter, N.W.; Loreto, F.; Sharkey, T.D. End product feedback effects on photosynthetic electron transport. Photosynth. Res. 1993, 35, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Van Gestel, N.C.; Nesbit, A.D.; Gordon, E.P.; Green, C.; Pare, P.W.; Thompson, L.; Peffley, E.B.; Tissue, D.T. Continuous light may induce photosynthetic downregulation in onion—Consequences for growth and biomass partitioning. Physiol. Plant. 2005, 125, 235–246. [Google Scholar] [CrossRef]
- Smith, A.M.; Stitt, M. Coordination of carbon supply and plant growth. Plant Cell Environ. 2007, 30, 1126–1149. [Google Scholar] [CrossRef] [PubMed]
- Stitt, M.; Lunn, J.; Usadel, B. Arabidopsis and primary photosynthetic metabolism—More than the icing on the cake. Plant J. 2010, 61, 1067–1091. [Google Scholar] [CrossRef] [PubMed]
- Cushman, K.E.; Tibbitts, T.W. The role of ethylene in the development of constant-light injury of potato and tomato. J. Am. Soc. Hortic. Sci. 1998, 123, 239. [Google Scholar] [CrossRef] [PubMed]
- Cushman, K.E.; Tibbitts, T.W.; Sharkey, T.D.; Wise, R.R. Constant-light injury of potato: Temporal and spatial patterns of carbon dioxide assimilation, starch content, chloroplast Integrity, and necrotic lesions. J. Am. Soc. Hortic. Sci. 1995, 120, 1032. [Google Scholar] [CrossRef]
- Lim, P.O.; Kim, H.J.; Nam, H.G. Leaf senescence. Annu. Rev. Plant Biol. 2007, 58, 115. [Google Scholar] [CrossRef] [PubMed]
- Krüll, F. The synchronizing effect of slight oscillations of light intensity on activity period of birds. Oecologia 1976, 25, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, M.G.; Apelt, F.; Carillo, P.; Krause, U.; Feil, R.; Mengin, V.; Lauxmann, M.A.; Köhl, K.; Nikoloski, Z.; Stitt, M.; et al. Getting back to nature: A reality check for experiments in controlled environments. J. Exp. Bot. 2017, 68, 4463–4477. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.M.; Guthrie, J.D.; Newell, J.M. Some effects of artificial climates on the growth and chemical composition of plants. Am. J. Bot. 1930, 17, 416–482. [Google Scholar] [CrossRef]
- Darrow, G.M. Tomatoes, berries and other crops under continuous light in Alaska. Science 1933, 78, 370. [Google Scholar] [CrossRef] [PubMed]
- Tibbitts, T.W.; Bennett, S.M.; Cao, W. Control of continuous irradiation injury on potatoes with daily temperature cycling. Plant Physiol. 1990, 93, 409–411. [Google Scholar] [CrossRef] [PubMed]
- Cushman, K.E.; Tibbitts, T.W. Root-zone temperature effects on continuous irradiation injury on potato. HortScience 1991, 26, 745. [Google Scholar]
- Cao, W.; Tibbitts, T.W. Temperature cycling periods affect growth and tuberization in potatoes under continuous light. HortScience 1992, 27, 344–345. [Google Scholar] [CrossRef]
- Yamashino, T.; Ito, S.; Niwa, Y.; Kunihiro, A.; Nakamichi, N.; Mizuno, T. Involvement of Arabidopsis clock-associated pseudo-response regulators in diurnal oscillations of gene expression in the presence of environmental time cues. Plant Cell Physiol. 2008, 49, 1839–1850. [Google Scholar] [CrossRef] [PubMed]
- Servaites, J.C.; Geiger, D.R.; Tucci, M.A.; Fondy, B.R. Leaf carbon metabolism and metabolite levels during a period of sinusoidal light. Plant Physiol. 1989, 89, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Geiger, D.R.; Shieh, W.J.; Lu, L.S.; Servaites, J.C. Carbon assimilation and leaf water status in sugar beet leaves during a simulated natural light regimen. Plant Physiol. 1991, 97, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.J.; Aradottir, G.I.; Smart, L.E.; Martin, J.L.; Caulfield, J.C.; Doherty, A.; Spark, C.A.; Woodcock, C.M.; Birkett, M.A.; Napier, J.A.; et al. The first crop plant genetically engineered to release an insect pheromone for defence. Sci. Rep. 2015, 5, 11183. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ullah, F.; Zhou, D.X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Meskauskiene, R.; Apel, K.; Laloi, C. No single way to understand singlet oxygen signalling in plants. EMBO Rep. 2008, 9, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Yonekura-Sakakibara, K.; Higashi, Y.; Nakabayashi, R. The Origin and Evolution of Plant Flavonoid Metabolism. Front. Plant Sci. 2019, 10, 943. [Google Scholar] [CrossRef]
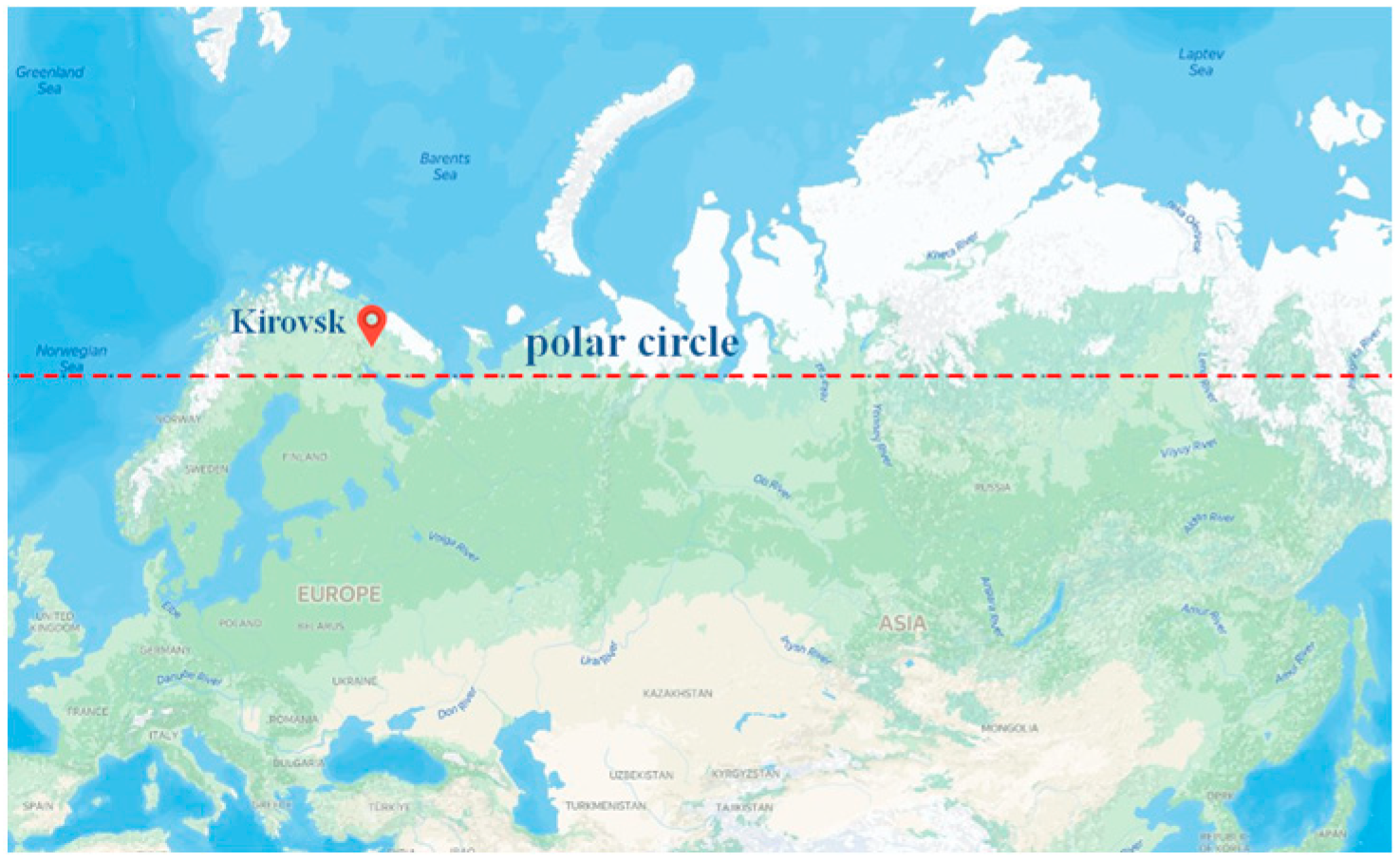
- Chupina, V.I. Natural and Antropogenic Soils of the Polar-Alpine Botanical Garden (Kirovsk). Soil Genes. Geogr. 2020, 75, 55–60. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 603, 591–592. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigment of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Kang, J.H.; Sugumaran, K.; Atulba, S.L.S.; Jeong, B.R.; Hwang, S.J. Light intensity and photoperiod influence the growth and development of hydroponically grown leaf lettuce in a closed-type plant factory system. Hortic. Environ. Biotechnol. 2013, 54, 501–509. [Google Scholar] [CrossRef]
- Meng, X.; Xing, T.; Wang, X. The role of light in the regulation of anthocyanin accumulation in Gerbera hybrid. Plant Growth Regul. 2004, 44, 243–250. [Google Scholar] [CrossRef]
- Nogues, S.; Backer, N.R. Effect of drought on photosynthesis in Mediterranean plants under UV-B radiation. J. Exp. Bot. 2000, 51, 1309–1317. [Google Scholar] [PubMed]
- Baroowa, B.; Gogoi, N. Biochemical changes in two Vigna spp. during drought and subsequent recovery. Ind. J. Plant Physiol. 2013, 18, 319–325. [Google Scholar] [CrossRef]
- Kolupaev, Y.E.; Fisova, E.N.; Yastreb, T.O.; Ryabchun, N.I.; Kirichenko, V.V. Effect of hydrogen sulfide donor on antioxidant state of wheat plants and their resistance to soil drought. Russ. J. Plant Physiol. 2019, 66, 59–66. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperioxidation in isolated chloroplasts I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant system in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
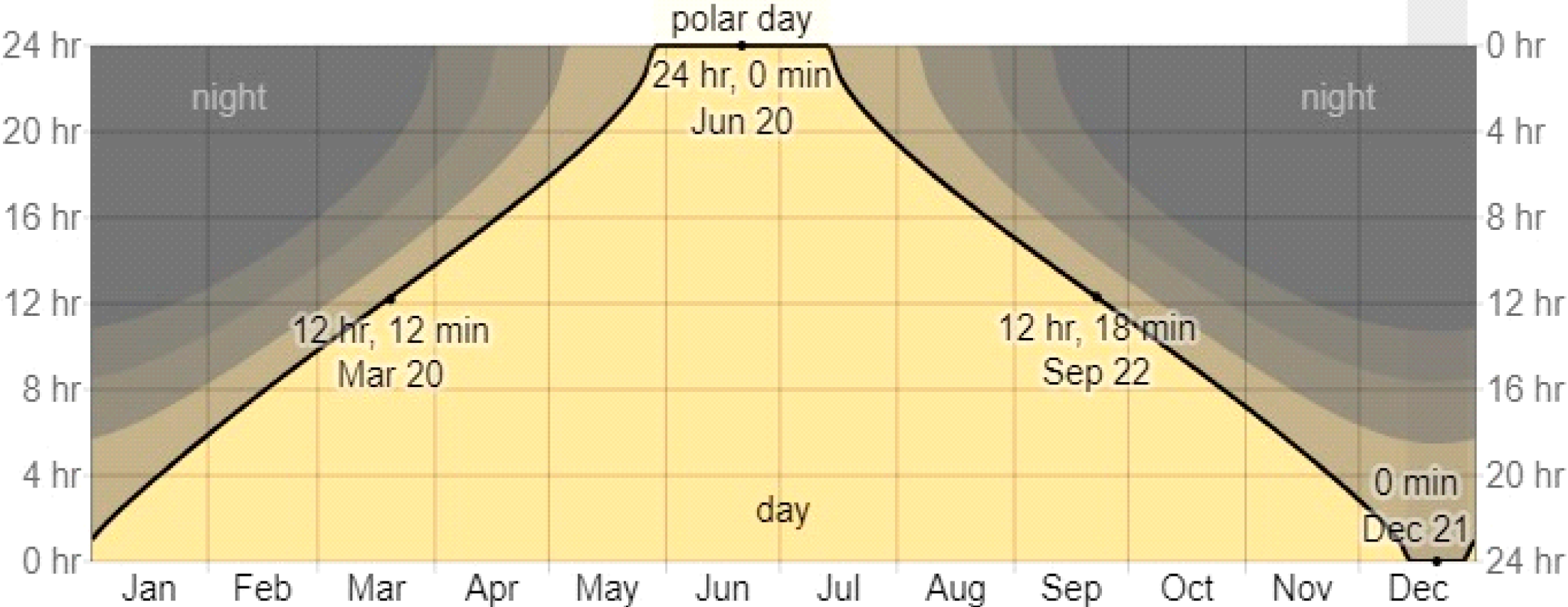
| Days | May | June | July | August | September |
|---|---|---|---|---|---|
| 1–5 | 17–42 | 23–07 | 24–00 | 18–41 | 14–38 |
| 6–10 | 18–24 | 24–00 | 23–59 | 17–57 | 14–00 |
| 11–15 | 19–08 | 24–00 | 22–44 | 17–18 | 13–22 |
| 16–20 | 19–53 | 24–00 | 21–27 | 16–40 | 12–42 |
| 21–25 | 20–46 | 24–00 | 20–29 | 16–02 | 12–04 |
| 26–30 | 21–52 | 24–00 | 19–35 | 15–09 | 11–28 |
| Average by month | 19–38 | 24–00 | 22–02 | 17–00 | 13–02 |
| Parameter | Photoperiod | Geranium sylvaticum | Geranium himalayense | Geum rivale | Geum coccineum | Potentilla erecta | Potentilla atrosanguinea |
|---|---|---|---|---|---|---|---|
| LMA, mg/cm2 | A-16 h | 3.52 b | 3.00 b | 4.78 b | 4.63 b | 4.94 b | 5.25 b |
| A-CL | 5.37 a | 4.98 a | 6.87 a | 7.3 a | 5.29 a | 7.96 a | |
| Fv/Fm | A-16 h | 0.766 a | 0.778 a | 0.783 a | 0.782 a | 0.823 a | 0.804 a |
| A-CL | 0.691 b | 0.755 b | 0.773 a | 0.753 b | 0.806 b | 0.741 b | |
| Chl, SPAD | A-16 h | 28.3 a | 37.9 a | 44.6 a | 40.2 a | 50.4 a | 49.6 a |
| A-CL | 26.0 a | 33.8 a | 43.9 a | 22.5 b | 49.8 a | 37.2 b | |
| Chl a + b, mg/g DW | A-16 h | 3.60 a | 3.87 a | 8.21 a | 7.96 a | 8.98 a | 7.11 a |
| A-CL | 2.64 b | 3.14 a | 5.12 b | 2.39 b | 8.13 a | 3.32 b | |
| Chl a/b | A-16 h | 2.3 b | 2.9 a | 2.5 a | 2.4 a | 2.0 a | 3.15 b |
| A-CL | 2.6 a | 3.0 a | 2.6 a | 2.6 a | 2.2 a | 3.34 a | |
| Carotenoids, mg/g DW | A-16 h | 0.51 a | 0.68 a | 1.0 a | 0.98 a | 0.72 a | 1.02 a |
| A-CL | 0.49 a | 0.69 a | 0.7 a | 0.35 b | 0.56 a | 0.61 b | |
| Chl/Carotenoids | A-16 h | 7.1 a | 5.7 a | 8.6 a | 8.1 a | 12.5 a | 7.0 a |
| A-CL | 5.4 b | 4.6 a | 6.8 b | 6.8 b | 14.5 a | 5.0 b | |
| LHCII, % | A-16 h | 69.9 a | 56.4 a | 64.0 a | 67.9 a | 79.3 a | 54.4 a |
| A-CL | 60.4 b | 54.8 a | 51.6 b | 64.7 a | 81.7 a | 48.4 b | |
| Anthocyanins, mg/mg FW | A-16 h | 0.80 b | 1.98 b | 0.98 b | 1.06 b | 0.59 b | 0.78 b |
| A-CL | 4.23 a | 3.94 a | 1.95 a | 3.73 a | 0.70 a | 6.23 a | |
| Flavonoids, mg/mg FW | A-16 h | 271.60 b | 404.3 b | 191.2 b | 189.9 b | 85.0 b | 205.1 b |
| A-CL | 497.40 a | 632.5 a | 243.1 a | 233.7 a | 140.5 a | 215.9 a | |
| MDA, μmol/g FW | A-16 h | 53.9 b | 51.4 b | 45.6 b | 93.1 b | 52.9 b | 45.4 b |
| A-CL | 79.0 a | 76.4 a | 88.0 a | 175.3 a | 67.3 a | 65.9 a | |
| H2O2, μmol/g FW | A-16 h | 1.36 b | 1.10 b | 0.78 b | 1.47 b | 1.06 b | 0.89 b |
| A-CL | 2.58 a | 1.85 a | 1.86 a | 2.02 a | 1.47 a | 1.44 a |
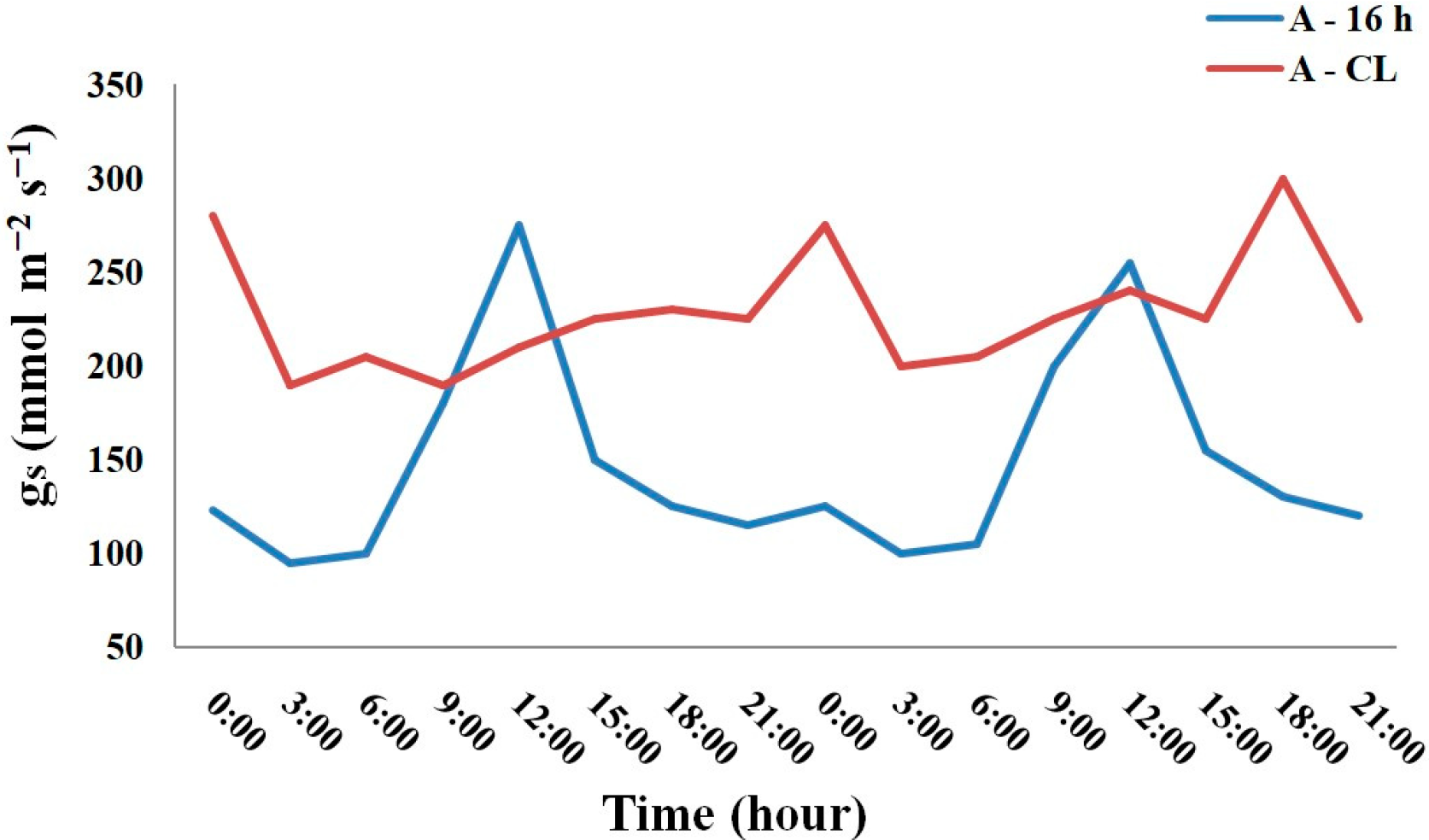
| Plant Name | Maximum/Minimum Daily gs, mmol/(m2 s) | |||
|---|---|---|---|---|
| A-16 h | A-CL | N-16 h | N-CL | |
| Geranium sylvaticum | 229/142 | 312/275 | 996/330 | 686/244 |
| Geranium himlayense | 363/90 | 297/188 | 711/267 | 645/248 |
| Geum rivale | 440/250 | 395/320 | 845/304 | 776/286 |
| Geum coccineum | 215/98 | 123/101 | 477/183 | 450/197 |
| Potentilla erecta | 507/160 | 521/223 | 812/195 | 790/217 |
| Potentilla atrosanguinea | 342/114 | 297/2 60 | 995/302 | 806/124 |
| Parameter | Photoperiod | Geranium sylvaticum | Geranium himalayense | Geum rivale | Geum coccineum | Potentilla erecta | Potentilla atrosanguinea |
|---|---|---|---|---|---|---|---|
| LMA, mg/cm2 | N-16 h | 3.83 b | 3.77 b | 3.23 a | 4.82 b | 3.95 b | 5.57 b |
| N-CL | 4.74 a | 4.23 a | 3.47 a | 7.50 a | 5.99 a | 6.81 a | |
| Fv/Fm | N-16 h | 0.805 a | 0.821 a | 0.820 a | 0.813 a | 0.820 a | 0.808 a |
| N-CL | 0.795 a | 0.802 b | 0.818 a | 0.794 b | 0.821 a | 0.800 a | |
| Chl, SPAD | N-16 h | 22.8 a | 41.3 a | 43.3 a | 40.2 a | 40.8 a | 45.1 a |
| N-CL | 20.6 a | 33.4 b | 38.3 a | 26.5 b | 45.1 a | 38.9 b | |
| Chl a + b, mg/g DW | N-16 h | 3.21 a | 5.24 a | 3.03 a | 6.43 a | 7.28 a | 4.51 a |
| N-CL | 2.71 b | 2.26 b | 1.98 b | 2.27 b | 6.47 b | 1.24 b | |
| Chl a/b | N-16 h | 2.6 b | 3.0 b | 3.0 a | 1.9 b | 1.8 b | 2.9 b |
| N-CL | 3.4 a | 3.6 a | 3.4 a | 2.5 a | 2.5 a | 3.7 a | |
| Carotenoids, mg/g DW | N-16 h | 0.52 a | 0.68 a | 0.50 a | 0.57 a | 0.82 a | 0.77 b |
| N-CL | 0.44 b | 0.41 b | 0.37 a | 0.28 b | 0.62 b | 0.66 a | |
| Chl/Carotenoids | N-16 h | 6.2 a | 7.8 a | 6.1 a | 11.3 a | 8.9 a | 5.9 a |
| N-CL | 6.3 a | 5.5 b | 5.4 a | 8.1 b | 10.4 a | 1.9 b | |
| LHCII, % | N-16 h | 61.2 a | 6.1 a | 55.0 a | 77.9 a | 78.5 a | 56.8 a |
| N-CL | 50.5 b | 47.8 b | 49.7 a | 51.2 b | 61.0 b | 42.6 b | |
| Anthocyanins, mg/mg FW | N-16 h | 1.18 a | 0.84 a | 0.87 a | 0.82 b | 1.17 a | 0.98 a |
| N-CL | 1.04 a | 0.95 a | 1.02 a | 1.17 a | 1.07 a | 0.76 b | |
| Flavonoids, mg/mg FW | N-16 h | 254.7 a | 354.1 a | 279.8 b | 334.8 a | 82.1 b | 131.4 b |
| N-CL | 277.5 a | 283.6 b | 343.1 a | 351.3 a | 139.0 a | 215.7 a | |
| MDA, μmol/g FW | N-16 h | 57.1 b | 37.0 b | 101.2 b | 61.3 a | 52.0 a | 38.1 a |
| N-CL | 69.7 a | 45.0 a | 120.9 a | 72.8 b | 55.7 a | 40.3 a | |
| H2O2, μmol/g FW | N-16 h | 4.48 b | 2.10 b | 1.9 b | 1.62 b | 1.82 b | 1.76 a |
| N-CL | 6.42 a | 4.01 a | 2.8 a | 2.54 a | 2.22 a | 1.82 a |
| Parameter | Year | ||
|---|---|---|---|
| 2021 | 2022 | ||
| Average temperature, °C | June | 13.3 | 11.9 |
| July | 14.8 | 16.5 | |
| August | 11.4 | 14.1 | |
| Effective temperature sum (base 5 °C) | June | From May 30 to Sept 11 401.0 | From May 28 to Sept 29 356.8 |
| July | 457.4 | 508.6 | |
| August | 350.9 | 436.9 | |
| Effective temperature sum (base 10 °C) | June | From June 2 to Aug 28 375.8 | From May 29 to Aug 28 282.5 |
| July | 407.3 | 499.6 | |
| August | 265.2 | 415.8 | |
| Relative air humidity, % | June | 7.6 | 79.9 |
| July | 76.9 | 85.0 | |
| August | 88.3 | 85.9 | |
| Amount of precipitation during the vegetation period, mm | 375 | 353 | |
| Vegetation period length, days | 66 | 75 | |
| Horizon, Depth (sm) | pH | Humus, (Cx1.724), % | TiO2 | Fe2O3 | CaO | Al2O3 | SiO2 | P2O5 | K2O | MgO | MnO |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Humus-illuvial podzols (Native plants) | |||||||||||
| O 0–10 | 5.2 | 49.0 | 1.17 | 4.5 | 3.3 | 7.2 | 61.0 | 1.43 | 1.43 | 0.73 | 0.08 |
| E 10–20 | 4.9 | 2.5 | 1.66 | 2.6 | 2.9 | 15.2 | 78/0 | 0.26 | 2.15 | 0.67 | 0.06 |
| BHF 20–25 | 5.6 | 16.5 | 0.68 | 8.7 | 1.1 | 21.3 | 61.1 | 0.38 | 0.61 | 0.68 | 0.05 |
| BHF 25–30 | 5.3 | 23.4 | 0.58 | 9.7 | 0.9 | 18.0 | 59.8 | 0.54 | 0.58 | 0.78 | 0.03 |
| BC 30–50 | 5.9 | 7.5 | 0.85 | 5.7 | 1.6 | 26.9 | 64.5 | 0.4 | 2.02 | 0.66 | 0.15 |
| Agrozem alfe–humus-stratified soils (Non-native plants) | |||||||||||
| Prh 0–15 | 6.7 | 4.7 | 1.87 | 9.6 | 2.6 | 28.0 | 64.0 | 0.59 | 2.22 | 1.11 | 0.39 |
| Ccr 15–26 | 6.5 | 2.4 | 1.76 | 9.1 | 1.9 | 32.3 | 67.0 | 0.27 | 2.23 | 0.95 | 0.39 |
| [E] 26–30 | 6.1 | 4.1 | 1.96 | 6.1 | 3.5 | 18.3 | 67.6 | 0.19 | 2/01 | 1.26 | 0.12 |
| [BH] 30–38 | 5.9 | 23.2 | 0.78 | 8.4 | 1.2 | 20.5 | 59.7 | 0.50 | 0.6 | 0.79 | 0.05 |
| [BHF] 30–50 | 5.9 | 23.3 | 0.53 | 6.7 | 1.1 | 21.2 | 60.6 | 0.51 | 0.58 | 0.69 | 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shibaeva, T.G.; Sherudilo, E.G.; Rubaeva, A.A.; Shmakova, N.Y.; Titov, A.F. Response of Native and Non-Native Subarctic Plant Species to Continuous Illumination by Natural and Artificial Light. Plants 2024, 13, 2742. https://doi.org/10.3390/plants13192742
Shibaeva TG, Sherudilo EG, Rubaeva AA, Shmakova NY, Titov AF. Response of Native and Non-Native Subarctic Plant Species to Continuous Illumination by Natural and Artificial Light. Plants. 2024; 13(19):2742. https://doi.org/10.3390/plants13192742
Chicago/Turabian StyleShibaeva, Tatjana G., Elena G. Sherudilo, Alexandra A. Rubaeva, Natalya Yu. Shmakova, and Alexander F. Titov. 2024. "Response of Native and Non-Native Subarctic Plant Species to Continuous Illumination by Natural and Artificial Light" Plants 13, no. 19: 2742. https://doi.org/10.3390/plants13192742