Abstract
Taraxacum kok-saghyz Rodin (TKS) is a recognized alternative source of natural rubber comparable to the rubber tree. The geranylgeranyl pyrophosphate synthase (GGPS) catalyzed the synthesis of geranylgeranyl pyrophosphate (GGPP), which is an important enzyme in the secondary metabolism pathway. In this study, we present the first analysis of the GGPS gene family in TKS, where a total of seven TkGGPS family members were identified. Their core motifs, conserved structural domains, gene structures, and cis-acting elements were described. In addition, two phylogenetic trees were constructed based on the Neighbor-Joining and Maximum-Likelihood methods, and the TkGGPSs were highly conserved and exhibited good collinearity with the other species. Transcriptome data showed that seven TkGGPS gene members were expressed in all the 12 tissues measured, and TkGGPS1, TkGGPS3, and TkGGPS6 were highly expressed in latex, suggesting that they may be associated with natural rubber synthesis. Meanwhile, quantitative real-time PCR (qRT-PCR) showed that the expression levels of the TkGGPS genes were regulated by the ethylene and methyl jasmonate (MeJA) pathways. Subcellular localization results indicated that all the TkGGPS proteins were also located in chloroplasts involved in photosynthesis in plants. This study will provide valuable insights into the selection of candidate genes for molecular breeding and natural rubber biosynthesis in TKS.
1. Introduction
Natural rubber (NR) is a kind of natural polymer compound with cis-1,4-polyisoprene as the main component [1,2]. More than 2500 plants in the world can produce natural rubber, but only a few of these species can produce high-molecular-weight rubber [3,4]. It is widely used in the fields of industry, agriculture, national defense, transportation, medicine, and daily life [5,6,7] because of its high strength, toughness, elasticity, and other excellent physical properties [8,9], such as conveyor belts, sponge products, gas masks, various tires, medical gloves, warm bags printing special parts, and other latex products [10,11,12]. With the rapid development of the economy and the improvement in human living standards, the demand for natural rubber is increasing [13]. At present, Hevea brasiliensis is almost the primary commercial source of natural rubber [14]. However, the rubber tree as a single species is at risk, with a long breeding season, limited planting area, and vulnerability to potential threats from climate change and disease [15,16,17]. Therefore, to increase the yield of natural rubber, it is necessary to develop an alternative source of natural rubber [18,19].
Taraxacum kok-saghyz Rodin (TKS), also known as Russian dandelion, is a self-incompatible perennial herbaceous plant of the family Asteraceae [20]. As a promising rubber-producing crop, it has the advantages of a short growth cycle, high adaptability, wide cultivation area, relatively simple genome, easy reproduction, and mature genetic transformation system [21,22]. Furthermore, high molecular weight natural rubber can be produced in the laticifers of TKS roots [23,24], and its rubber production is comparable to the yield of the H. brasiliensis [25]. In addition, TKS can also produce some valuable compounds, such as inulin, pentacyclic triterpene, and sterol; the change in their content has a certain effect on the synthesis of natural rubber [26].
Plants have the following two main metabolic pathways for isoprenoid biosynthesis: mevalonate (MVA) and methylerythritol (MEP). The MVA pathway mainly occurs in the cytoplasm, while the MEP pathway operates in the plastid [27]. These two pathways provide precursors for isoprene biosynthesis, changes in gene expression and enzyme activity in either pathway will affect the metabolic balance of cells and plant growth and development [28]. Geranylgeranyl pyrophosphate synthase (GGPS) serves as an essential enzyme in the metabolism of isoprenoids [29], catalyzing the synthesis of 20-carbon geranylgeranyl diphosphate (GGPP) from isopentenyl pyrophosphate (IPP) and its isomer dimethylallyl pyrophosphate (DMAPP), with the substrate being provided by the MVA or MEP pathways [30]. In addition, GGPP is a significant intermediate compound in the bio-production of various isoprenoids, such as natural rubber, carotene, gibberellin, tocopherol, and cholesterol [31,32]. The GGPS genes have been studied in certain plants so far. Heterologous expression of the sunflower genes (HaGGPS) on chloroplasts in tobacco and Arabidopsis thaliana results in rapid growth, early flowering, and an increase in gibberellin (GA) content and the number of flowers and seed pods [33]. The identification of the GGPS gene family in several cotton varieties and the expression analysis of GGPSs in plant development and abiotic stress indicated that GGPS participated in chlorophyll synthesis of cotton, promoted fiber development, and responded to abiotic and hormonal stress. [34,35]. The cloning and expression analysis of the GGPS1 gene in Salvia miltiorrhiza showed that the GGPS1 gene was expressed in all tested tissues, but its expression level was higher in leaves at the flowering stage [36]. The gene cloning, expression, and function analysis of GGPS synthase from H. brasiliensis indicated that GGPS synthase in H. brasiliensis rubber latex may be involved in natural rubber and β-carotene biosynthesis [32]. Twelve GGPS synthase gene family members were predicted in A. thaliana, suggesting that some key genes are provided for specific tissues, developmental stages, or metabolic pathways [37]. However, the identification and characterization of GGPS gene family members have not been reported in T. kok-saghyz.
In this study, we comprehensively identified seven GGPS family members in TKS. First of all, we analyzed the physicochemical properties, secondary and tertiary structures of TkGGPS proteins, and performed bioinformatics analysis of conserved motif and domain, gene structure, chromosome localization, inter-specific phylogenetic relationship, and collinearity features. Then, we investigated the expression of TkGGPS genes in different tissues and under hormone treatments by analyzing transcriptomic data and qRT-PCR. Finally, we determined the subcellular localization of all TkGGPS proteins. The results obtained in this study will contribute to a deeper understanding of the role of TkGGPSs in the growth and development and the biosynthesis of natural rubber in T. kok-saghyz.
2. Results
2.1. Identification of the GGPS Gene Family in T. kok-saghyz
A total of seven GGPS genes were identified based on the latest genomic datas by local BLAST analysis and HMM search with the GGPS protein sequence of Arabidopsis thaliana as a query. We also selected Arabidopsis thaliana, a closely related and rubber-producing related species to further characterize the TKGGPS gene family using an orthology-based method. As a result, six TkGGPS members were identified, which did not include TkGGPS5 (Supplementary Tables S1 and S2), so we compared the structure and physicochemical properties of the TkGGPS5 protein with those of several other members and found that this member shared similar structure and properties with the others; therefore, we classified TkGGPS5 as a member of the TkGGPS gene family.
The seven TkGGPS members were named according to the length of their amino acid sequences, and their physicochemical properties were analyzed, as shown in Table 1. The length of the TkGGPS amino acid sequences ranged from 262 aa (TkGGPS1) to 436 aa (TkGGPS7), the molecular weights ranged from 28.53 kDa (TkGGPS2) to 48.34 kDa (TkGGPS7), their predicted isoelectric points ranged from 5.14 (TkGGPS3) to 9.76 (TkGGPS5), and the instability index ranged from 34.27 (TkGGPS2) to 62.03 (TkGGPS1). All GGPS proteins were expected to be localized on chloroplasts, and most members of this family had an instability index of more than 40, indicating that they are unstable. The GRAVY ranged from −0.247 (TkGGPS5) to 0.096 (TkGGPS6), most TkGGPS members have more hydrophilic regions than hydrophobic regions (Supplementary Figure S1), while the average hydrophobicity of TkGGPS 2 and TkGGPS 6 was 0.038 and 0.096, respectively, suggesting that they may be hydrophobic proteins. In addition, the trans-membrane domain analysis of the amino acid sequence of TkGGPSs (Supplementary Figure S2) showed that all the amino acids of these seven TkGGPS members were outside the membrane, indicating that TkGGPS protein synthesis could play a direct role without transmembrane transport. The prediction and analysis of signal peptides of seven TkGGPS proteins (Supplementary Figure S3) showed that there were no signal peptides in TkGGPS proteins, and they were non-secreted proteins.

Table 1.
Basic information of GGPS genes identified in T. kok-saghyz.
2.2. Secondary and Three-Dimensional Structures Analysis of the TkGGPSs
The secondary structure of the seven TkGGPS proteins was predicted and analyzed, and the results are shown in Table 2. The α-helices and random coils are the main structures; there is no β-folding structure, but the extension chain and β-turn are scattered throughout the protein. Among them, the α-helix occupies 42.12% (TkGGPS5)–63.26% (TkGGPS2), while the β-turn occupies 3.67% (TkGGPS7)–9.09% (TkGGPS5), the extended strand occupies 4.92% (TkGGPS2)–16.97% (TkGGPS5), and the random coil chain accounts for 25.37% (TkGGPS3)–38.30% (TkGGPS7).
Table 2.
Secondary structure predictions of TkGGPSs.
The three-dimensional structure of the seven TkGGPS proteins was predicted by homology modeling, and the results are shown in Figure 1. The similarity between TkGGPS proteins and the template sequence ranges from 34.05% (TkGGPS5) to 87.12% (TkGGPS7). It can be observed that the three-dimensional structures of the TkGGPS proteins are predominantly composed of α-helices and random coils, resulting from further twisting and folding processes. From a three-dimensional structure perspective, there is a significant resemblance among the different members of the TkGGPSs. Therefore, it can be inferred that these members likely share similar protein functions in this plant. Only TkGGPS5 has a different three-dimensional structure from the other members, which may be due to its differentiation during evolution.
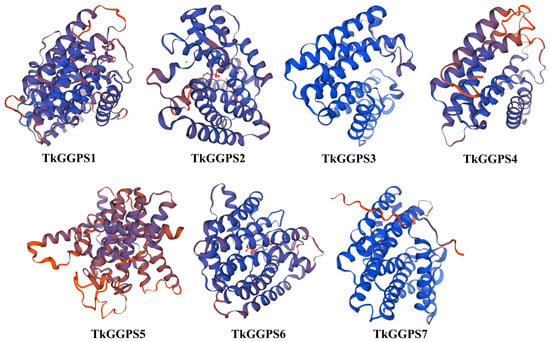
Figure 1.
Prediction of the three-dimensional structure of TkGGPS proteins in T. kok-saghyz. The blue chain represents consistency with the template sequence and a higher score, while the red represents a lower score.
2.3. Conserved Motif, Domain and Gene Structure of the TkGGPS in T. kok-saghyz
The MEME and Pfam websites were employed to identify the conserved motifs and domains of these TkGGPSs (Figure 2). A total of ten motifs were identified. TkGGPS2, TkGGPS3, and TkGGPS6, TkGGPS5 and TkGGPS7, TkGGPS1 and TkGGPS4, respectively, from the same clade shared common motif compositions (Figure 2A). Conserved motifs in the TkGGPSs indicated that almost all the TkGGPSs contained motifs 1 and 2. However, TkGGPS6 lacked motif 10, TkGGPS7 lacked motifs 5 and 7, while TkGGPS2 and TkGGPS3 lacked motifs 6, 8, and 10. TkGGPS1 and TkGGPS4 lacked motifs 5, 6, 7, and 9. Motif 9 was present in TkGGPS2, TkGGPS3, TkGGPS6, and TkGGPS7. TkGGPS5 contained motifs 1, 2 and 6. All TkGGPSs contained one polyprenyl_synt superfamily domain, which was typical of a GGPS protein (Figure 2B).
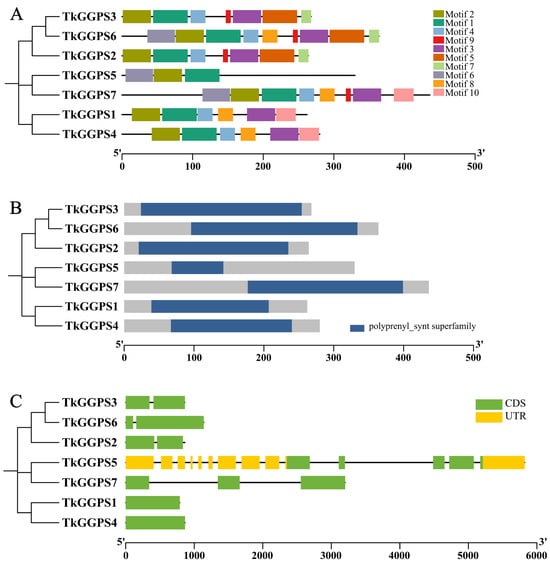
Figure 2.
Phylogenetic relationship, motif composition, conserved domains, and gene structure of the TkGGPS family proteins. (A) The phylogenetic tree and the motif composition in TkGGPSs. Different colored boxes represent putative motifs, and the lengths of motifs in each protein are presented proportionally. The phylogenetic tree was constructed using the Neighbor-Joining (NJ) method. (B) The polyprenyl_synt superfamily domain of TkGGPSs. (C) The exon–intron structure of the TkGGPS genes. Yellow boxes indicate 5′ UTR and 3′ UTR, green boxes indicate exons, black lines indicate introns.
To gain further insights into the structural diversity of TkGGPSs, the exon/intron organizations of TkGGPS genes were determined by Tbtools software. Most TkGGPS genes contained exons and introns, but TkGGPS1 and TkGGPS4 have no intron (Figure 2C). The multiple sequence alignment results revealed a partial or total variation in the five key residues of all seven TkGGPSs (Supplementary Figure S4). The members of the TkGGPS include five conserved amino acid regions named I, II, III, IV, and V. The first conserved region is GKXXR (X is an arbitrary amino acid), the third region is GQ, and the fourth region is KT. In addition, the second and fifth regions are two aspartate-rich regions, DDXXXXD and DDXXD, that are called the FARM (the first aspartate-rich motif) and SARM (the second aspartate-rich motif) regions, which are the binding sites of allyl substrate and IPP, respectively.
2.4. Cis-Acting Elements Analysis of the TkGGPS Gene Family
Cis-acting elements are widely recognized as crucial factors in gene adaptation to environmental changes and the regulation of growth and development. Therefore, to gain preliminary insights into the role of the seven TkGGPS genes, we analyzed the potential cis-acting elements in their promoter regions. A comprehensive examination identified a total of 33 types of cis-acting elements (Figure 3).
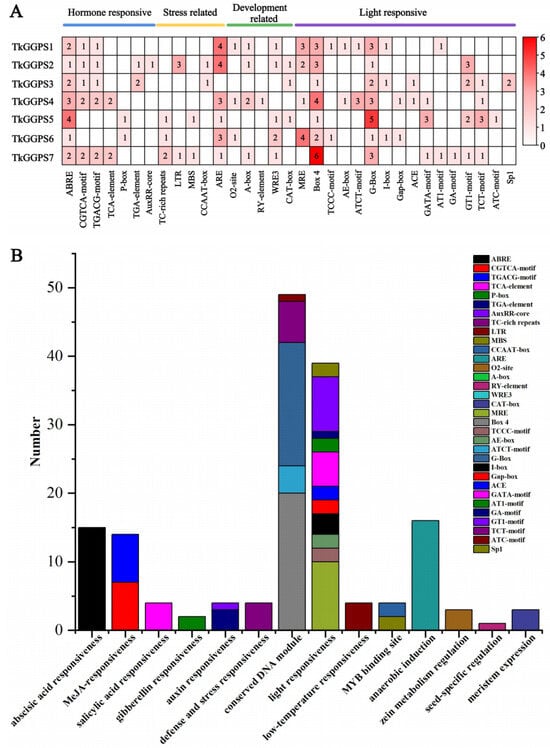
Figure 3.
Predicted cis-acting elements of the promoters of TkGGPSs. (A) Classification and heatmap expression analysis of all cis-acting elements on promoters of TkGGPSs. The 2.0 kb promoter sequences of seven TkGGPS genes were analyzed using the PlantCARE database. The different colors represent the number of each cis-acting element. (B) Statistics of all cis-acting elements on promoters of TkGGPSs.
These cis-acting elements were further divided into the following four functional categories: hormone response, stress response, growth and development, and light response (Figure 3A). For the hormone-responsive category, the abscisic acid-responsive element accounted for the largest proportion (ABRE7/7), followed by the MeJA-responsive elements (CGTGA-motif, 5/7, TGACG-motif, 5/7), auxin-responsive elements (TGA-element, 2/7, AuxRR-core, 1/7), salicylic acid-responsive element (TCA-element, 2/7), and gibberellin-responsive element (P-box, 2/7). For the stress-responsive category, six TkGGPSs contained the anaerobic induction element (ARE), three TkGGPSs owned defense and stress responsiveness elements (TC-rich repeats), two TkGGPSs possessed the low-temperature responsiveness element (LTR), two TkGGPSs had the binding site MYBHv1 element (CCAAT-box), and two TkGGPSs had the drought-inducibility element (MBS). For the developmental regulation category, WRE3 related to developmental regulation was the most abundant element (5/7), followed by A-box related to developmental regulation (4/7), CAT-box involved in meristem expression (3/7), O2-site involved in zein metabolism regulation (3/7), and RY-element involved in seed-specific regulation (1/7). Notably, within the light-responsive element category, the conserved DNA module subcategory mainly exhibited motifs corresponding to 16 types of cis-acting elements, including MRE, Box 4, TCCC-motif, AE-box, ATCT-motif, G-Box, I-box, Gap-box, ACE, GATA-motif, AT1-motif, GA-motif, GT1-motif, TCT-motif, ATC-motif, and Sp1 (Figure 3B). Interestingly, all seven TkGGPSs contained a large number of light-responsive elements (Figure 3A), suggesting a close association between the TkGGPSs and photosynthesis in TKS.
2.5. Phylogenetic Analysis of TkGGPS Proteins
To investigate the phylogenetic relationship among GGPS proteins upon representative monocotyledonous and dicotyledonous plants, we constructed phylogenetic trees with 7 TkGGPS, 5 HbGGPS, 5 LsGGPS, 5 TmGGPS, 13 NtGGPS, 6 OsGGPS, 12 AtGGPS and 9 HaGGPS proteins from eight species, namely T. kok-saghyz, H. brasiliensis, Lactuca sativa, Taraxacum mongolicum, Nicotiana tabacum, Oryza sativa, A. thaliana, and Helianthus annuus. The protein names used for AtGGPS are those known from TAIR, and the remaining GGPS names for each species are based on the length of the amino acid sequence (Supplementary Table S3). The GGPS of all the above eight species were classified into six clades using the Neighbor-Joining (NJ) and Maximum-Likelihood methods (ML), respectively (Figure 4). In the phylogenetic tree constructed by the NJ method, TkGGPS1 and TkGGPS4 belonged to clade II, TkGGPS5 and TkGGPS7 belonged to clade III, and two proteins (TkGGPS3 and TkGGPS6) belonged to clade VI (Figure 4A). Similarly, these six TkGGPS members are also in three clades (II, V, and VI) in the phylogenetic tree constructed with the ML method (Figure 4B). However, TkGGPS2 is in a different cluster from the other TKS members in both phylogenetic trees, so it was hypothesized that TkGGPS2 may play a different role. Interestingly, most of the A. thaliana members are in the same cluster, which was presumably more evolutionarily conserved and with similar functions.
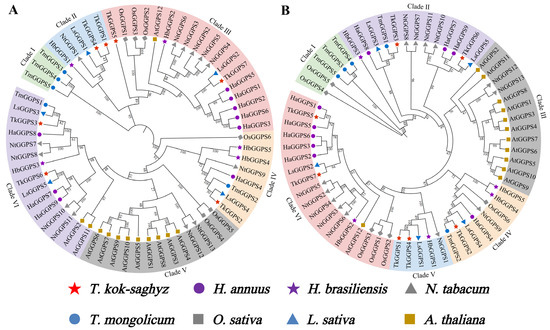
Figure 4.
Phylogenetic analysis of TkGGPS proteins in T. kok-saghyz. Phylogenetic analyses were conducted on GGPS proteins from T. kok-saghyz (TKS, red star), H. brasiliensis (Hb, purple star), L. sativa (Ls, blue triangle), T. mongolicum (TM, blue circle), N. tabacum (Nt, dimgray triangle), O. sativa (Os, dimgray square), A. thaliana (At, gold square) and H. annuus (Ha, purple circle). ClustalW was used for multiple sequence alignment. Phylogenetic trees were constructed using the Neighbor-Joining (NJ) method with 1000 bootstrap repeats (A) and the Maximum likelihood (ML) method with 5000 bootstrap repeats (B). Different colors represent different clades of GGPS. The number on the branch represents the Bootstrap method (BM) value.
2.6. Location and Collinearity Analysis of TkGGPS Genes on chromosomes
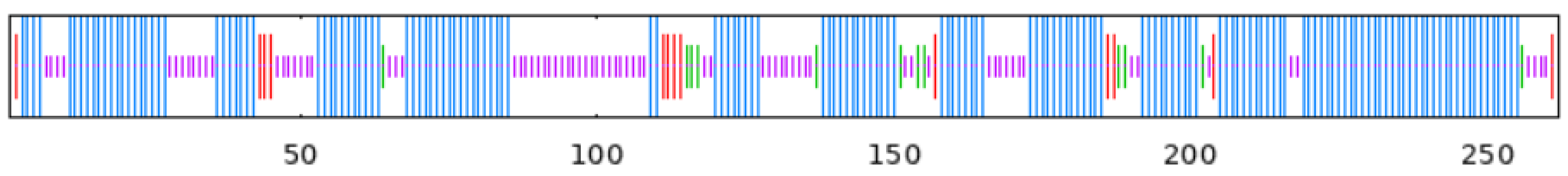
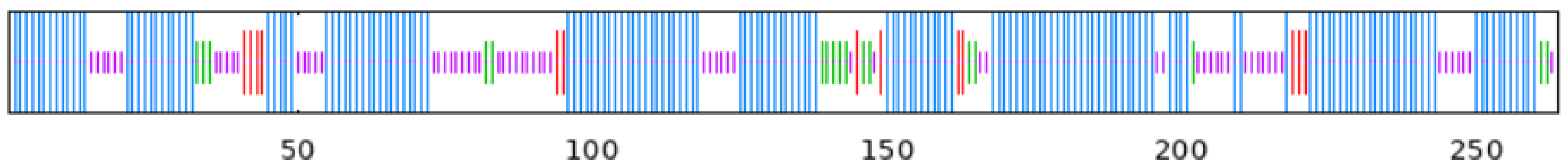
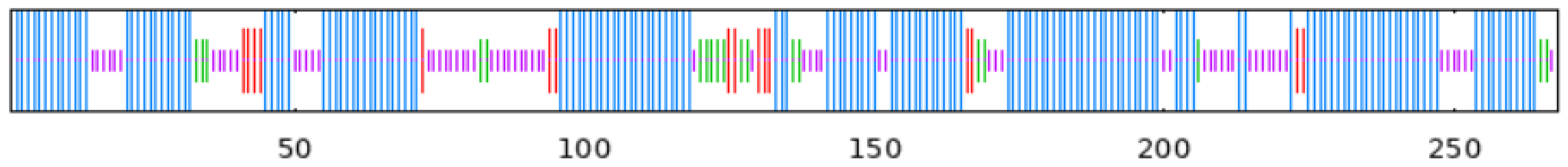
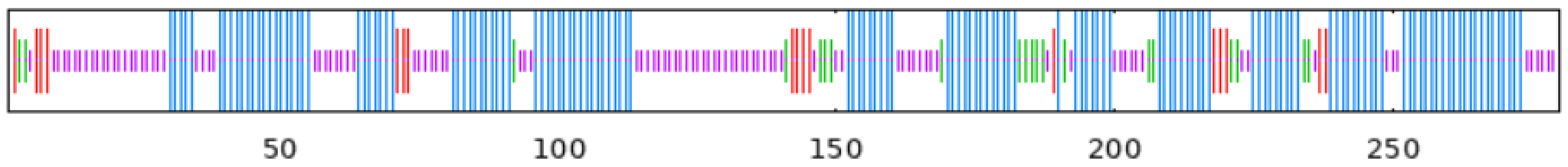
To observe whether the members of the TkGGPS gene family are distributed in clusters on chromosomes, the GGPS localizations of T. kok-saghyz and four similar species were investigated, and the results are shown in Figure 5. A total of eight chromosomes in TKS, of which seven members of TkGGPS were distributed on chromosomes 2, 3, 6, and 7. TkGGPS1 and TkGGPS4 were located on chromosome 2. TkGGPS5 and TkGGPS7 were located on chromosome 3, and TkGGPS2 and TkGGPS3 were located on chromosome 6. Only TkGGPS6 was located on chromosome 7 (Figure 5A). In addition, there were a total of eight chromosomes in T. mongolicum, and the TmGGPS members were distributed on chromosomes 1, 6, and 7 (Figure 5B). There were nine chromosomes in L. sativa, among which the LsGGPS family members were distributed on chromosomes 3, 5, 7, and 9, respectively (Figure 5C). There were five chromosomes in A. thaliana, and members of the AtGGPS family were distributed on chromosomes 1, 2, 3, and 4 (Figure 5D). In H. annuus, there were 17 chromosomes, and the HaGGPS members were distributed on chromosomes 5, 6, 8, 10, and 12 (Figure 5E).
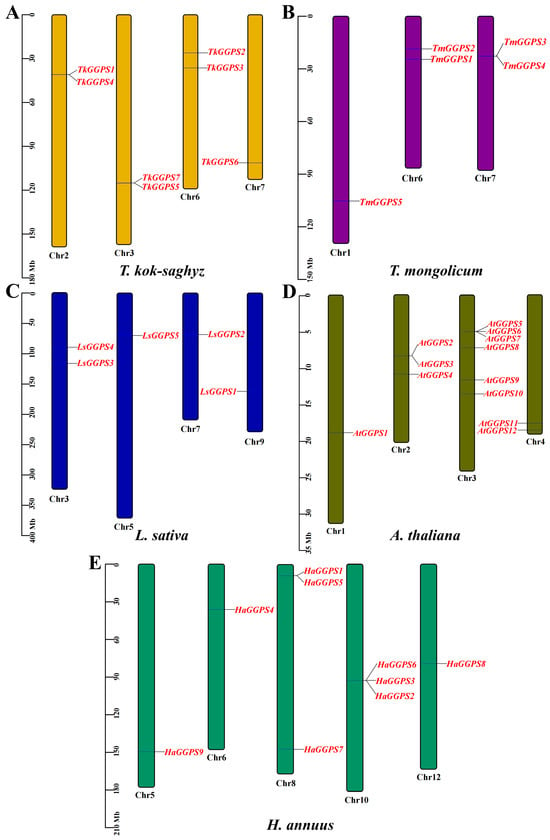
Figure 5.
Chromosome Distribution of GGPS genes in the genomes of five species. (A) T. kok-saghyz, (B) T. mongolicum, (C) L. sativa, (D) A. thaliana, and (E) H. annuus. The scale on the left represents the chromosome size, the length of chromosomes is measured in Mb. The chromosome number is indicated below each chromosome. GGPS gene numbers are shown on the two sides of each chromosome, and they are marked in red.
To assess the collinearity relationships of the TkGGPS gene family within the various common model crops, we analyzed the molecular phylogeny of the GGPS gene family using the One Step MCScanX—Super Fast toolkit in TBtools software, as shown in Figure 6. The results showed that T. kok-saghyz exhibited three collinear gene pairs with T. mongolicum and four collinear gene pairs with A. thaliana, L. sativa, and H. annuus, respectively. Notably, TkGGPS3 and TkGGPS6 have homologous gene pairs with all four species (Supplementary Table S4), and TkGGPS7 and HaGGPS6 are homologous genes. In addition, TkGGPS2 is homologous with LsGGPS4 and TmGGPS2, respectively, suggesting that they may play similar roles in different species.
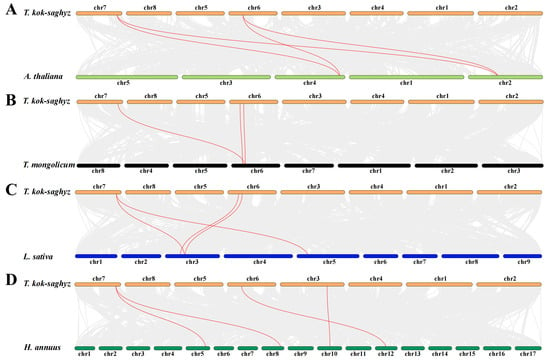
Figure 6.
Collinearity analysis of TkGGPS genes. Genome collinearity features between (A) T. kok-saghyz and A. thaliana, (B) T. kok-saghyz and T. mongolicum, (C) T. kok-saghyz and L. sativa, (D) T. kok-saghyz and H. annuus. All the collinear GGPS genes are shown in red lines. The grey lines indicate collinear blocks.
2.7. Transcriptomics and qRT-PCR Analysis of TkGGPS Genes
Since natural rubber is synthesized in latex from TKS roots, we performed transcriptome analysis to explore the expression patterns of the seven TkGGPS gene members in 12 different TKS tissues including latex (Figure 7A). We found that all seven TkGGPS members were expressed in latex, with TkGGPS1 having a relatively high expression, and TkGGPS3 and TkGGPS6 both having very high expressions, with a more than one-hundred-fold expression, whereas TkGGPS2, TkGGPS4, and TkGGPS7 had a low expression. We hypothesized that TkGGPS1, TkGGPS3, and TkGGPS6 may play an important role in NR synthesis. In addition, this result indicated that the TkGGPS2 expression was higher in young lateral roots compared to other tissues, while the TkGGPS1 gene was highly expressed in main roots, lateral roots, and stems. TkGGPS6 expression was generally high, and TkGGPS5 expression was more stable in all the tissues tested. Furthermore, TkGGPS3 and TkGGPS7 expression was high in the young leaves, mature leaves, flowers, peduncles, and seeds, whereas the expression pattern in roots showed a more stable trend. It indicated that they play a part in the plant growth and developmental processes.
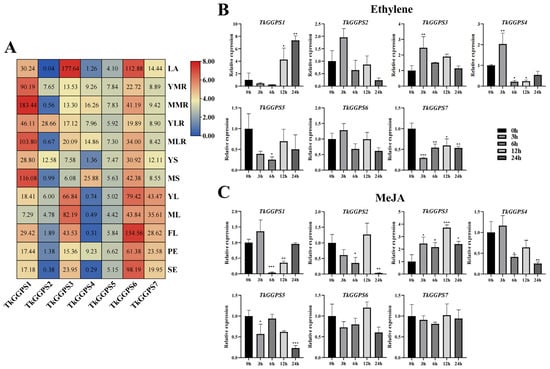
Figure 7.
Gene expression of 7 TkGGPS genes in TKS. (A) Heat map expression analysis of TkGGPS genes in TKS different tissues based on transcriptome data. LA, latex; YMR, young main root; MMR, mature main root; YLR, young lateral root; MLR, mature lateral root; YS, young stem; MS, mature stem; YL, young leaf; ML, mature leaf; FL, flower; PE, peduncle; SE, seed. Red indicates high expression levels, and blue indicates low expression levels. The numbers on the box represent the fragments per kilobase million (FPKM) value. The expression values were normalized by FPKM to create the heat map, using the average FPKM of three biological replicates. The expression level of 7 TkGGPS genes in TKS roots treated with ethylene (B) and MeJA (C) at 0 h (control), 3 h, 6 h, 12 h, and 24 h, respectively. Data were normalized to β-actin. Vertical bars for qRT-PCR indicate the standard deviation, while an asterisk indicates the summary p-value of the independent samples t-test of the corresponding gene compared to the control (* p < 0.05, ** p < 0.01, *** p < 0.001).
To understand the function of the TkGGPS gene members in plant development, we used different hormones (ethylene and MeJA) to treat a 6-month-old wild-type TKS and analyzed it by qRT-PCR to probe the expression levels of seven TkGGPSs in the roots and leaves. The expression of the seven TkGGPS genes was different in response to these two hormones, as shown in Figure 7. The expression of TkGGPS3 was up-regulated while two genes (TkGGPS5 and TkGGPS7) were down-regulated (compared to the control) by ethylene treatment in TKS roots (Figure 7B). After ethylene treatment for 3 h, the expressions of TkGGPS2, TkGGPS4, and TkGGPS6 were up-regulated and then down-regulated. At 12 h and 24 h, and the expression level of TkGGPS1 was up-regulated, and the accumulation was up to seven times at 24 h, thus we speculated that ethylene could regulate gene expression. However, the expression levels of two genes (TkGGPS2 and TkGGPS3) had little response to ethylene treatment in the TKS leaves; TkGGPS6 was up-regulated, while TkGGPS4 and TkGGPS5 were down-regulated (Supplementary Figure S5A). After treatment with MeJA, the expression levels of TkGGPS1 and TkGGPS4 in TKS roots were up-regulated at 3 h, whereas those of TkGGPS2 and TkGGPS6 were up-regulated at 12 h. The expression levels of the TkGGPS5 and TkGGPS7 genes were largely unaffected by MeJA (Figure 7C). Similarly, after MeJA treatment of leaves, the response of TkGGPS5 and TkGGPS7 to MeJA treatment remained small, and the expression of the TkGGPS3 gene was still up-regulated, demonstrating that TkGGPS3 plays an important role in the regulation of the growth and development of TKS as well as the metabolic pathways of ethylene and MeJA (Supplementary Figure S5B).
2.8. Subcellular Localization Analysis of TkGGPS Proteins
To determine the subcellular localization of the seven TkGGPS proteins, green fluorescent protein (eGFP)-expressing vector-fused TkGGPS proteins were constructed, and this plasmid was transiently expressed in tobacco leaf epidermal cells. As shown in Figure 8, the signals of 35S–TkGGPS–eGFP are found in the chloroplast. Additionally, subcellular localization prediction analysis revealed that most of the TkGGPS proteins are predominantly localized in the chloroplast. These localization patterns are consistent with the expected properties of the theoretical prediction. The above results suggest that TkGGPS members are not only present in the latex of the TKS roots involved in NR synthesis but also in the chloroplasts involved in plant photosynthesis.
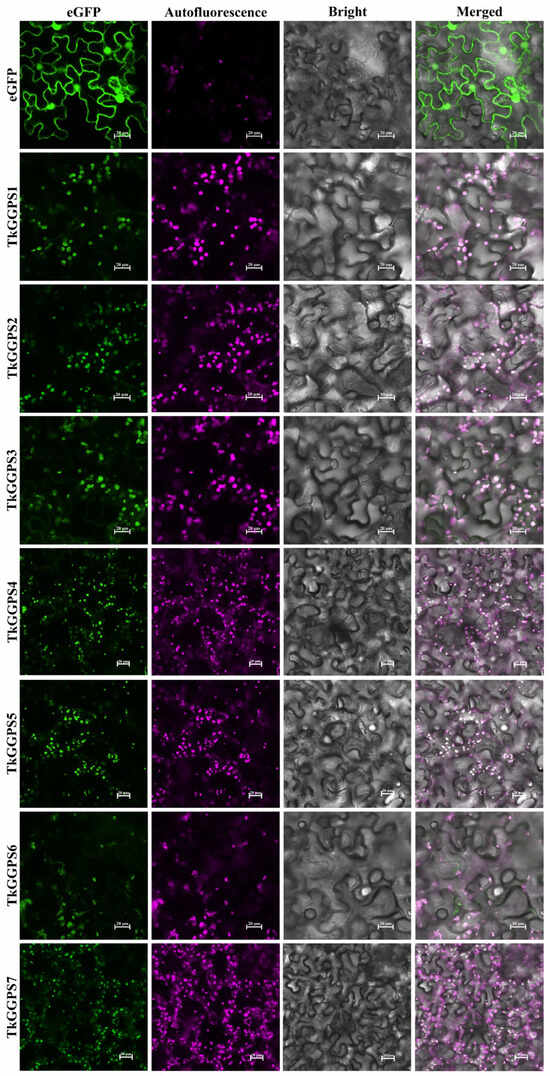
Figure 8.
Subcellular localization of TkGGPS proteins in Nicotiana tabacum leaves. pCAMBIA1300-35S–eGFP and pCAMBIA1300-35–TkGGPS–eGFP fusion proteins were transiently expressed in N. tabacum leaves. The fields included green fluorescence filed (488 nm), chloroplast autofluorescence field (640 nm), bright field, and merged filed. Empty vector control showing the expression of 35S–eGFP in epidermal cells of N. tabacum, and co-localization of 35S–eGFP with TkGGPS proteins observed by chloroplast autofluorescence. Bars = 20 µm.
3. Discussion
Geranylgeranyl pyrophosphate synthase (GGPS) is a key enzyme in the synthesis of GGPP and is widely found in plants, animals, bacteria, and fungi. It catalyzes the condensation reaction of farnesyl pyrophosphate (FPP) and isopentenyl pyrophosphate (IPP) to produce GGPP, which is an important precursor for the synthesis of natural rubber [38]. NR is synthesized in the roots of TKS and stored in rubber particles, which has important application value in human life [39]. At present, the GGPS gene has only been reported in a few plants, such as sweet potato [40], Taxus media [41], A. thaliana [42], N. tabacum [43], and so on. However, the identification and analysis of the GGPS gene family is rarely studied in T. kok-saghyz. Through previous transcriptomics and proteomic studies, GGPS was found to be important for NR biosynthesis in small rubber particles in TKS roots [44]. Here, we identified and verified a total of seven members of the TkGGPS gene family based on the newly published genome by Lin et al. [45]. In this publication, Lin et al. researchers found that the contents of GPP and GGPP in TKS latex were high. Therefore, it is necessary to explore the molecular characteristics and regulation mechanism of the TkGGPS gene family. In this study, the physicochemical property, conserved motif, domain, gene structure, cis-acting element, phylogenetic relationship, chromosome localization, and collinearity of the TkGGPS gene family members were analyzed, which is of great significance for further understanding the molecular characteristics of the GGPS genes in TKS.
The amino acid sequence of a protein determines how its higher structure is formed as well as its ultimate function [46], so it is imperative to analyze the physicochemical properties of a protein. The number of amino acids, the molecular weight, and the isoelectric point of the TkGGPS members were described in this study (Table 1). Most members of TkGGPS are hydrophilic, have no transmembrane domains, no signal peptides, and are unstable, non-secreted proteins (Supplementary Figures S1–S3). α-helix and random curling are the main components of the secondary and tertiary structure of TkGGPS proteins (Table 2, Figure 1). Conserved motifs and domains are usually associated with the function of proteins [47]. By analyzing the motif and domain structures, we found that although several motifs are missing in TkGGPS, some motifs are present in all members of TkGGPS, such as motif 1 and motif 2 (Figure 2A). It was noteworthy that the polyprenyl_synt superfamily domain was present in all the TkGGPS members (Figure 2B), suggesting that they are highly conserved in TkGGPS and may play similar roles in plant growth and development. Introns are genomic sequences removed from the corresponding RNA transcripts of genes and play an important role in species’ genetic and evolutionary processes. In earlier reports, introns of some genes were lost over time during gene replication [48]. Among the seven TkGGPS members in this study, introns were missing in some genes, such as TkGGPS1 and TkGGPS4, which may be related to their evolution (Figure 2C). Multiple sequence alignment results (Supplementary Figure S4) showed that there are two aspartate-rich motifs in the TkGGPS members, FARM and SARM. The FARM motif is sequenced as DDxxD or DDxxxxD, while the SARM motif is always DDxxD; both motifs bind three Mg2+ ions, which is conducive to the binding of IPP and DMAPP to GGPS substrates and determines the catalytic activity of GGPS [49,50].
Phylogenetic relationships are used to represent the relationship between species or amino acid sequences, and accurate phylogenetic trees can increase our understanding of evolutionary relationships [51]. In the present study, all GGPSs were grouped into a broad category, the phylogenetic tree divides the GGPSs of eight species into six clades (Figure 4). It was concluded that the TkGGPS members in the same clade have similar gene structure, conserved domain, and motif composition (Figure 2). Collinear relationships between species are common in nature, and homologous genes perform the same function in the different species [52]. Here, TkGGPS genes showed good collinearity with T. mongolicum, L. sativa, and H. annuus, which belong to dicotyledonous plants in the Asteraceae family. In particular, the TkGGPS3 and TkGGPS6 genes on chromosomes 6 and 7 of T. kok-saghyz shared homologous genes with four other species (Figure 5 and Figure 6), which were predicted to have potentially evolved from a common ancestor, with a high degree of conservation and a slower evolutionary rate.
Transcriptomics provides comprehensive access to transcript information for species-specific tissues or organs [53]. Here, almost all the gene members of TkGGPS were expressed in TKS latex (Figure 7A), where NR is synthesized. In the previous study, a total of 102 rubber biosynthesis-related genes were identified by genome assembly, including 19 genes for initiator synthesis [54]. It was shown that the NR initiators—the GPS, FPS, and GGPS enzymes—can catalyze IPP and DAMPP to generate isopentenyl diphosphate intermediates such as GPP, FPP, and GGPP, thereby initiating rubber biosynthesis in vitro [55]. In this study, the expression of TkGGPS1 was higher in the TKS roots and latex than in the other tissues examined, such as the leaves and flowers, while the expression of the TkGGPS3 and TkGGPS6 genes was significantly higher in latex than in the other gene members, which were in agreement with the conclusion drawn by Lin et al. [45]. We speculated that these three genes could be candidate genes and may play a key role in NR synthesis. In addition, all the TkGGPS gene members were differentially expressed in various tissues in Taraxacum kok-saghyz, which may be related to their role and specific function in each tissue.
Ethylene and MeJA are widely involved in regulating plant growth and development and secondary metabolite synthesis [56]. Cis-acting elements are involved in the regulation of gene expression and are necessary for plant growth, adaptation, and response to the environment [57]. In this study, we predicted the cis-acting elements of the TkGGPS gene members, including hormone-responsive elements, stress-responsive elements, growth- and development-responsive elements, and light-responsive elements (Figure 3). It has been reported that exogenous MeJA can activate JA biosynthesis and signal transduction and regulate the downstream MEP, MVA, and natural rubber biosynthesis pathways by altering gene expression [58]. Exogenous ethylene can promote seed germination and rooting [59], and exogenous methyl jasmonate can induce CgGGPS expression in Corylus avellana L. Gasaway [60]. Therefore, we treated different tissues of TKS with ethylene and MeJA and performed qRT-PCR to detect the expression level of the TkGGPS genes (Figure 7); we found that these two hormones induced the expression of GGPS genes in TKS roots, and the expression levels of TkGGPS3 were always up-regulated after treatment. Moreover, these two hormones also affect the expression of GGPS genes in the TKS leaves (Supplementary Figure S5). Therefore, the expression of the TkGGPS gene members was regulated by the ethylene and MeJA pathways, and these members had different expression levels after hormone induction.
The subcellular localization of GGPS is significantly differentiated in plants to accommodate its different physiological functions [61]. In plants, the GGPS gene was first successfully cloned in Capsicum annuum, and the in situ immunolocalization experiment proved that the CaGGPS protein was only located in plastids. Furthermore, it was heavily induced in the process of chloroplast to chromoplast transformation in mature fruit, and the corresponding enzyme activity was also enhanced [62]. In A. thaliana, a total of twelve members of the AtGGPS family were identified in earlier studies, among which AtGGPS1 was located in the mitochondria, AtGGPS3 and AtGGPS4 in the endoplasmic reticulum, and AtGGPS2, AtGGPS6, AtGGPS7, and AtGGPS11 in the chloroplasts or plastids [63]. In rice, OsGGPS1 is mainly located in the plastoglobules and stroma of chloroplasts [64]. In Liriodendron tulipifera, the LtGGPS2 protein is localized to the chloroplasts, suggesting that this gene may be involved in carotenoid and chlorophyll synthesis [65]. In H. brasiliensis, HbGGPSs were detected in all the tissues examined, including the latex, leaves, flowers, and pedicels, which may participate in the synthesis of a variety of compounds [32]. In this study, we predicted similar results for all seven TkGGPS protein members. Most genes such as TkGGPS1, TkGGPS3, TkGGPS6, and TkGGPS7 were highly expressed in stems, leaves, and flowers (Figure 7A), and the expression of certain genes was up-regulated after the treatment of TKS leaves with ethylene and MeJA (Supplementary Figure S5), suggesting that the TkGGPS genes were also more active in the chloroplasts. Plants generally do not contain chloroplasts in mature latex [66]; therefore, in terms of subcellular localization, it was not surprising to find that almost all the TkGGPS members were located on chloroplasts (Figure 8). Based on all the above analysis, we surmised that the TkGGPS gene may be involved in the biosynthesis of NR in latex and important isoprenoids such as carotenoids, chlorophylls and gibberellins in chloroplasts, which will provide us with some assistance in the subsequent validation of its function.
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
TKS was collected from the river area of Shihezi City, Xinjiang, China, and successfully transplanted and cultivated in our laboratory, Shihezi University. Then, tissue culture seedlings of TKS were cultivated in the phytotron (temperature 23 °C, air humidity 68%, light 16 h) consisting of a 3:1:1 mixture of nutrient soil, vermiculite, and perlite for 5–6 months until flowering.
4.2. Identification and Characterization Analysis of the TkGGPS Gene Family
To identify GGPSs from the TKS genome, the 15 GGPS protein sequences in A. thaliana were downloaded from NCBI (https://www.ncbi.nlm.nih.gov/, accessed on 5 October 2023) and invoked as queries to perform local BLAST and HMM algorithm-based searches [67,68] against T. kok-saghyz and T. mongolicum genome databases from the National Genomics Data Center (NGDC) Genome Warehouse (GWH, https://ngdc.cncb.ac.cn/gwh/, accessed on 5 October 2023, (BioProject accession: PRJCA005187) [69]. In addition, other species’ genome databases in this study were downloaded from NCBI (https://www.ncbi.nlm.nih.gov/, accessed on 6 October 2023), such as A. thaliana (assembly TAIR10.1) (Supplementary Tables S7 and S8) [31], N. tabacum (assembly Ntab-TN90), L. sativa (assembly Lsat_Salinas_v11), H. annuus (assembly HanXRQr2.0-SUNRISE), H. brasiliensis (assembly ASM165405v1), and O. sativa (assembly GCF_001433935.1). We also selected the above eight species to further characterize the TKGGPS gene family by orthology-based method (https://github.com/davidemms/OrthoFinder, accessed on 6 October 2023) [70,71]. The functional domain prediction (Accession cl46106) was performed using the conserved domain tool of NCBI CDD (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi, accessed on 7 October 2023) [72]. To validate the accuracy of the conserved domains, these potential sequences (Pfam number (PF00348)) were further searched against the Pfam database (http://pfam-legacy.xfam.org/, accessed on 7 October 2023) [73].
In addition, the basic physicochemical properties, such as amino acid sequence length, protein molecular weight (Mw), theoretical isoelectric point (pI), and hydrophilicity of TkGGPSs, used the ExPASy website (https://web.expasy.org/compute_pi/, accessed on 10 October 2023) [74] for analysis. Prediction of the signal peptides and transmembrane domains of TkGGPS proteins used the Signal 6.0 (https://services.healthtech.dtu.dk/services/SignalP-6.0/, accessed on 13 October 2023) [75] and TMHMM 2.0 websites (https://services.healthtech.dtu.dk/services/TMHMM-2.0/, accessed on 13 October 2023) [76]. The subcellular localization of TkGGPS proteins was analyzed using the Cell-PLoc 2.0 tool (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/, accessed on 14 October 2023) [77]. The secondary and three-dimensional structures were predicted by the online websites SOPMA (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html, accessed on 14 October 2023) [78] and SWISS-MODEL (https://swissmodel.expasy.org/interactive, accessed on 17 October 2023) [79].
4.3. Phylogeny, Gene Structure and Conserved Motif Analysis of TkGGPSs
First, the multiple sequence alignment of full-length GGPS amino acid sequences was performed using the ClustalW program in MEGA 11, and they were further analyzed in GeneDoc v2.7.000 software [80]. Then, the phylogenetic tree was constructed using the Neighbor-Joining (NJ) method with 1000 bootstrap replications and the Maximum likelihood (ML) method with 5000 bootstrap repeats in MEGA 11 [81]. The phylogenetic tree of GGPS proteins was beautified using EvolView v. 2 (http://evolgenius.info/evolview/#/login, accessed on 20 October 2023) [82]. Exon/intron sites and length information were extracted from the respective genome annotation GFF files of the T. kok-saghyz from the NGDC Genome Warehouse (BioProject accession: PRJCA005187), and the structure drawings were scaled and displayed using the Gene Structure View (Advanced) in TBtools v1.099 (Toolbox for Biologists) [83]. The conserved protein motif of the TkGGPS family was identified using MEME Suite v.5.5.5 (https://meme-suite.org/meme/tools/meme, accessed on 23 October 2023) [84], with a maximum number of 10 motifs.
4.4. Chromosome Distribution and Collinearity Analysis of TkGGPS Genes
Interspecific collinearity analysis and chromosomal localization were performed with TBtools. The chromosomal localization was operated by Gene Location Visualize from GTF/GFF, and collinearity analysis was conducted using the One Step MCScanX—Super Fast toolkit [85].
4.5. Identification of Cis-Regulatory Elements in the Promoters of TKGGPS Genes
Cis-acting elements were obtained from the 2000 bp upstream regions of the TkGGPS genes. The predictions were made using the PlantCARE website (http://bioinformatics.psb.ugent.be/webtools/plantcare/html, accessed on 28 October 2023) [86]. The detected elements were divided into different response types based on their annotated functions.
4.6. Quantitative Real-Time PCR (qRT-PCR) Analysis of TkGGPS Genes
To investigate the expression of TkGGPS genes in different tissues, the 6-month-old TKS seedlings with similar size and growth were selected for hormonal treatment. The seedling was hydroponic in Hogren culture solution with 100 µmol/L ethylene and 1 mmol/L MeJA hormones. Samples were collected at different treatment periods of 0 h, 3 h, 6 h, 12 h, and 24 h and packed in pre-cooled 10mL tubes, then quickly frozen in liquid nitrogen and stored in a refrigerator at −80 °C. We performed Quantitative Real-time PCR (qRT-PCR) analysis using the SYBR Green qPCR Master Mix on the Roche LightCycler 480 instrument. The reaction volume was 20 μL. The RNA was extracted according to the difficult-to-extract Plant Total RNA Kit (purchased from Magen Biotech, Guangzhou, China), and the Reverse-Transcribed cDNA Kit was used HiScript ⅡQ RT SuperMix for qPCR (+gDNA wiper) (derived from Vazyme Biotech Co., Ltd., Nanjing, China). All qRT-PCR primer sequences were designed with Primer 5.0 software (PREMIER Biosoft, San Francisco, CA, USA) and are shown in Supplementary Table S5.
To ensure the accurate normalization of the gene expression data, we used a stable and reliable reference gene, the Tkactin gene (GenBank accession: DY824357), as an internal control [87,88,89]. In addition, gene expression levels were quantified as relative fold changes, with the 0 h time point serving as the control group and assigned a value of one. Each reaction was conducted with three biological replicates. The obtained qRT-PCR data were analyzed using the 2−ΔΔCT method, a widely accepted approach for relative quantification of gene expression [90]. The bar graph was drawn by GraphPad Prism (version 9.0, GraphPad Software, San Diego, CA, USA).
4.7. Transcriptomic Data Analysis of T. kok-saghyz
The transcriptomic raw data of Taraxacum kok-saghyz in different tissues were obtained from the Genome Sequence Archive (GSA) database (https://ngdc.cncb.ac.cn/gsa/, accessed on 15 December 2023, Accession Number: PRJCA000437). The raw data were converted into files in fastq format for quality assessment and filtered low-quality sequences [91]. Then, the obtained reads were compared to the reference genomes, and gene expression analysis was calculated according to the FPKM value [92,93]. The TBtools software was used to generate the heatmap of the TkGGPS genes.
4.8. Subcellular Localization Assay of TkGGPS Proteins
The coding sequences (CDS) of the TkGGPSs was amplified using primers that removed terminators but contained arms homologous to the expression vector, and the target fragment was recovered. Then, the sequences were cloned into the KpnI and SpeI digested pCAMBIA1300–eGFP vector by homologous recombination. Seven recombinant vectors included 35S–TkGGPS1–eGFP, 35S–TkGGPS2–eGFP, 35S–TkGGPS3–eGFP, 35S–Tk–GGPS4–eGFP, 35S–TkGGPS5–eGFP, 35S–TkGGPS6-eGFP, and 35S–TkGGPS7–eGFP, along with the empty vector 35S–1300–eGFP. The gene cloning primer sequence is shown in Supplementary Table S6.
The plasmids were introduced into Agrobacterium tumefaciens (strain GV3101) and injected into 4-week-old N. benthamiana leaves. Photographic observations of the tobacco leaf epidermal cells were carried out in dark culture for 48 h after transfection on a Nikon Eclipse Ti2 ultra-high-resolution confocal fluorescence microscope (Nikon, Japan). For GFP fluorescence and chloroplast autofluorescence analysis, 488/640 nm excitation laser lines were used.
5. Conclusions
TKS is a promising substitute crop for natural rubber, which is widely used in production and life. In this study, seven members of the TkGGPS gene family were identified by bioinformatics methods, and their evolutionary mechanism and functional characteristics were studied. All TkGGPS members are not only presented in latex but also located in chloroplasts. qRT-PCR analysis showed that the TkGGPS gene family members are also induced by ethylene and MeJA. Moreover, the transcriptome data analysis also indicated that TkGGPS1, TkGGPS3, and TkGGPS6 would be the key candidate genes for NR synthesis, which will lay the foundation for our future research. These results provide valuable perceptions into the involvement of TkGGPS in natural rubber synthesis and breeding in TKS, as well as the utilization of TKS resources.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants13192788/s1, Figure S1. Hydrophilic/hydrophobic analysis of all TkGGPS members; Figure S2. Transmembrane domain analysis of all TkGGPS members; Figure S3. Signal peptide analysis of all TkGGPS members; Figure S4. Multiple sequence alignment of the GGPS amino acid sequence in T. kok-saghyz; Figure S5. Gene expression of 7 TkGGPS genes at 0 h (control), 3 h, 6 h, 12 h, and 24 h after Ethylene and at 0 h (control), 6 h, 12 h, and 24 h after MeJA treatments in TKS leaves; Table S1. OrthoFinder result for eight species; Table S2. OrthoFinder result; Table S3. The amino acid sequence of eight species; Table S4. Collinearity relationships between TkGGPS and four species’ GGPS; Table S5. qRT-PCR primer sequence; Table S6. Gene cloning primer; Table S7. The whole proteome sequence of Arabidopsis thaliana (NCBI); Table S8. The whole proteome sequence of Arabidopsis thaliana (TAIR).
Author Contributions
L.W (Lili Wang)., H.H., J.W., F.W., H.L. and Q.X. conceived and designed the research experiments; L.W (Lili Wang)., H.H. and J.W. wrote the original manuscript. L.W (Lili Wang)., H.H. and J.W. contributed equally to this work. Z.M., L.W (Lei Wang)., J.Z., X.J., P.D. and L.Z. performed the experiments; H.H., J.W., Z.M. and X.J. analyzed the data. L.Z. and Z.M. contributed materials and analytical tools; F.W., H.L. and Q.X. wrote and corrected this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant number 32060072, Xinjiang Production and Construction Corps Guiding Technology Program Project, grant number 2023ZD050, and Special Project of New Varieties Cultivation in Shihezi University, grant number YZZX202403.
Data Availability Statement
All data generated or analyzed during this study are included in this article.
Acknowledgments
We are very grateful to Xuchu Wang from the Guizhou University for providing the Taraxacum kok-saghyz seeds. We are also very grateful to Paul P. Dijkwel from the School of Natural Sciences, Massey University, for his helpful suggestions.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Böttner, L.; Malacrinò, A.; Schulze Gronover, C.; van Deenen, N.; Müller, B.; Xu, S.; Gershenzon, J.; Prüfer, D.; Huber, M. Natural rubber reduces herbivory and alters the microbiome below ground. New Phytol. 2023, 239, 1475–1489. [Google Scholar] [CrossRef] [PubMed]
- van Beilen, J.B.; Poirier, Y. Establishment of new crops for the production of natural rubber. Trends Biotechnol. 2007, 25, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Cherian, S.; Ryu, S.B.; Cornish, K. Natural rubber biosynthesis in plants, the rubber transferase complex, and metabolic engineering progress and prospects. Plant Biotechnol. J. 2019, 17, 2041–2061. [Google Scholar] [CrossRef] [PubMed]
- van Beilen, J.B.; Poirier, Y. Guayule and Russian dandelion as alternative sources of natural rubber. Crit. Rev. Biotechnol. 2007, 27, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Guerra, N.B.; Sant’Ana Pegorin, G.; Boratto, M.H.; de Barros, N.R.; de Oliveira Graeff, C.F.; Herculano, R.D. Biomedical applications of natural rubber latex from the rubber tree Hevea brasiliensis. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 126, 112126. [Google Scholar] [CrossRef]
- Rojruthai, P.; Sakdapipanich, J.; Wiriyanantawong, J.; Ho, C.C.; Chaiear, N. Effect of Latex Purification and Accelerator Types on Rubber Allergens Prevalent in Sulphur Prevulcanized Natural Rubber Latex: Potential Application for Allergy-Free Natural Rubber Gloves. Polymers 2022, 14, 4679. [Google Scholar] [CrossRef]
- Yip, E.; Cacioli, P. The manufacture of gloves from natural rubber latex. J. Allergy Clin. Immunol. 2002, 110, S3–S14. [Google Scholar] [CrossRef]
- Pichayakorn, W.; Chaiya, P.; Chinpaisal, C.; Phaechamud, T. Natural rubber blends for floating theophylline beads. Int. J. Biol. Macromol. 2023, 224, 725–738. [Google Scholar] [CrossRef]
- Yamashita, S.; Takahashi, S. Molecular Mechanisms of Natural Rubber Biosynthesis. Annu. Rev. Biochem. 2020, 89, 821–851. [Google Scholar] [CrossRef]
- Klongklaew, P.; Khamjapo, P.; Sae-Oui, P.; Jittham, P.; Loykulnant, S.; Intiya, W. Characterization and Application in Natural Rubber of Leucaena Leaf and Its Extracted Products. Polymers 2023, 15, 3698. [Google Scholar] [CrossRef]
- Sethulekshmi, A.S.; Saritha, A.; Joseph, K. A comprehensive review on the recent advancements in natural rubber nanocomposites. Int. J. Biol. Macromol. 2022, 194, 819–842. [Google Scholar] [CrossRef] [PubMed]
- Supriya, R.; Priyadarshan, P.M. Genomic technologies for Hevea breeding. Adv. Genet. 2019, 104, 1–73. [Google Scholar] [CrossRef] [PubMed]
- Stonebloom, S.H.; Scheller, H.V. Transcriptome analysis of rubber biosynthesis in guayule (Parthenium argentatum gray). BMC Plant Biol. 2019, 19, 71. [Google Scholar] [CrossRef] [PubMed]
- Bushman, B.S.; Scholte, A.A.; Cornish, K.; Scott, D.J.; Brichta, J.L.; Vederas, J.C.; Ochoa, O.; Michelmore, R.W.; Shintani, D.K.; Knapp, S.J. Identification and comparison of natural rubber from two Lactuca species. Phytochemistry 2006, 67, 2590–2596. [Google Scholar] [CrossRef]
- Buranov, A.U.; Elmuradov, B.J. Extraction and characterization of latex and natural rubber from rubber-bearing plants. J. Agric. Food Chem. 2010, 58, 734–743. [Google Scholar] [CrossRef]
- Mooibroek, H.; Cornish, K. Alternative sources of natural rubber. Appl. Microbiol. Biotechnol. 2000, 53, 355–365. [Google Scholar] [CrossRef]
- Ramirez-Cadavid, D.A.; Cornish, K.; Michel, F.C. Taraxacum kok-saghyz (TK): Compositional analysis of a feedstock for natural rubber and other bioproducts. Ind. Crop. Prod. 2017, 107, 624–640. [Google Scholar] [CrossRef]
- Arias, M.; Hernandez, M.; Remondegui, N.; Huvenaars, K.; van Dijk, P.; Ritter, E. First genetic linkage map of Taraxacum koksaghyz Rodin based on AFLP, SSR, COS and EST-SSR markers. Sci. Rep. 2016, 6, 31031. [Google Scholar] [CrossRef]
- Salehi, M.; Cornish, K.; Bahmankar, M.; Naghavi, M.R. Natural rubber-producing sources, systems, and perspectives for breeding and biotechnology studies of Taraxacum kok-saghyz. Ind. Crop. Prod. 2021, 170, 113667. [Google Scholar] [CrossRef]
- Wollenweber, T.E.; van Deenen, N.; Roelfs, K.U.; Prüfer, D.; Gronover, C.S. Microscopic and Transcriptomic Analysis of Pollination Processes in Self-Incompatible Taraxacum koksaghyz. Plants 2021, 10, 555. [Google Scholar] [CrossRef]
- Cheng, Y.; Luo, J.; Li, H.; Wei, F.; Zhang, Y.; Jiang, H.; Peng, X. Identification of the WRKY Gene Family and Characterization of Stress-Responsive Genes in Taraxacum kok-saghyz Rodin. Int. J. Mol. Sci. 2022, 23, 10270. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, I.; Choi, S.C.; Bae, S.W.; Park, J.C.; Ryu, S.B. Heterologous activation of the Hevea PEP16 promoter in the rubber-producing laticiferous tissues of Taraxacum kok-saghyz. Sci. Rep. 2020, 10, 10844. [Google Scholar] [CrossRef] [PubMed]
- Lightfoot, D.A.; Wieghaus, A.; Prüfer, D.; Schulze Gronover, C. Loss of function mutation of the Rapid Alkalinization Factor (RALF1)-like peptide in the dandelion Taraxacum koksaghyz entails a high-biomass taproot phenotype. PLoS ONE 2019, 14, e0217454. [Google Scholar] [CrossRef]
- Wieghaus, A.; Roelfs, K.U.; Twyman, R.M.; Prüfer, D.; Schulze Gronover, C. Comparative Transcriptome Analysis in Taraxacum koksaghyz to Identify Genes that Determine Root Volume and Root Length. Front. Genet. 2021, 12, 784883. [Google Scholar] [CrossRef]
- Zhang, Y.; Iaffaldano, B.J.; Zhuang, X.; Cardina, J.; Cornish, K. Chloroplast genome resources and molecular markers differentiate rubber dandelion species from weedy relatives. BMC Plant Biol. 2017, 17, 34. [Google Scholar] [CrossRef]
- Niephaus, E.; Müller, B.; van Deenen, N.; Lassowskat, I.; Bonin, M.; Finkemeier, I.; Prüfer, D.; Schulze Gronover, C. Uncovering mechanisms of rubber biosynthesis in Taraxacum koksaghyz—Role of cis-prenyltransferase-like 1 protein. Plant J. Cell Mol. Biol. 2019, 100, 591–609. [Google Scholar] [CrossRef]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Pu, X.; Dong, X.; Li, Q.; Chen, Z.; Liu, L. An update on the function and regulation of methylerythritol phosphate and mevalonate pathways and their evolutionary dynamics. J. Integr. Plant Biol. 2021, 63, 1211–1226. [Google Scholar] [CrossRef]
- Liang, M.H.; Liang, Y.J.; Jin, H.H.; Jiang, J.G. Characterization and Functional Identification of a Gene Encoding Geranylgeranyl Diphosphate Synthase from Dunaliella bardawil. J. Agric. Food Chem. 2015, 63, 7805–7812. [Google Scholar] [CrossRef]
- Thabet, I.; Guirimand, G.; Guihur, A.; Lanoue, A.; Courdavault, V.; Papon, N.; Bouzid, S.; Giglioli-Guivarc’h, N.; Simkin, A.J.; Clastre, M. Characterization and subcellular localization of geranylgeranyl diphosphate synthase from Catharanthus roseus. Mol. Biol. Rep. 2012, 39, 3235–3243. [Google Scholar] [CrossRef]
- Wang, C.; Chen, Q.; Fan, D.; Li, J.; Wang, G.; Zhang, P. Structural Analyses of Short-Chain Prenyltransferases Identify an Evolutionarily Conserved GFPPS Clade in Brassicaceae Plants. Mol. Plant 2016, 9, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Takaya, A.; Zhang, Y.W.; Asawatreratanakul, K.; Wititsuwannakul, D.; Wititsuwannakul, R.; Takahashi, S.; Koyama, T. Cloning, expression and characterization of a functional cDNA clone encoding geranylgeranyl diphosphate synthase of Hevea brasiliensis. Biochim. Biophys. Acta (BBA)-Gene Struct. Expr. 2003, 1625, 214–220. [Google Scholar] [CrossRef]
- Tata, S.K.; Jung, J.; Kim, Y.H.; Choi, J.Y.; Jung, J.Y.; Lee, I.J.; Shin, J.S.; Ryu, S.B. Heterologous expression of chloroplast-localized geranylgeranyl pyrophosphate synthase confers fast plant growth, early flowering and increased seed yield. Plant Biotechnol. J. 2016, 14, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.; Qanmber, G.; Wei, Z.; Yu, D.; Li, Y.H.; Gan, L.; Li, F.; Wang, Z. Genome-wide characterization and expression analysis of geranylgeranyl diphosphate synthase genes in cotton (Gossypium spp.) in plant development and abiotic stresses. BMC Genom. 2020, 21, 561. [Google Scholar] [CrossRef]
- Feng, W.; Mehari, T.G.; Fang, H.; Ji, M.; Qu, Z.; Jia, M.; Wang, D.; Ditta, A.; Khan, M.K.R.; Cao, Y.; et al. Genome-wide identification of the geranylgeranyl pyrophosphate synthase (GGPS) gene family involved in chlorophyll synthesis in cotton. BMC Genom. 2023, 24, 176. [Google Scholar] [CrossRef]
- Zhang, L.; Dai, Z.; Cui, G.; Cheng, Y.; Qi, X.; Gao, Z. Cloning and characterization of geranylgeranyl diphosphate synthase gene of Salvia miltiorrhia. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Med. 2009, 34, 2704–2708. [Google Scholar]
- Beck, G.; Coman, D.; Herren, E.; Ruiz-Sola, M.A.; Rodríguez-Concepción, M.; Gruissem, W.; Vranová, E. Characterization of the GGPP synthase gene family in Arabidopsis thaliana. Plant Mol. Biol. 2013, 82, 393–416. [Google Scholar] [CrossRef]
- Engprasert, S.; Taura, F.; Kawamukai, M.; Shoyama, Y. Molecular cloning and functional expression of geranylgeranyl pyrophosphate synthase from Coleus forskohlii Briq. BMC Plant Biol. 2004, 4, 18. [Google Scholar] [CrossRef]
- Yang, Y.; Qin, B.; Chen, Q.; Zhang, J.; Zhang, L.; Nie, Q.; Liu, S. Comparative full-length transcriptome analysis provides novel insights into the regulatory mechanism of natural rubber biosynthesis in Taraxacum kok-saghyz Rodin roots. Ind. Crop. Prod. 2022, 175, 114278. [Google Scholar] [CrossRef]
- Chen, W.; He, S.; Liu, D.; Patil, G.B.; Zhai, H.; Wang, F.; Stephenson, T.J.; Wang, Y.; Wang, B.; Valliyodan, B.; et al. A Sweetpotato Geranylgeranyl Pyrophosphate Synthase Gene, IbGGPS, Increases Carotenoid Content and Enhances Osmotic Stress Tolerance in Arabidopsis thaliana. PLoS ONE 2015, 10, e0137623. [Google Scholar] [CrossRef]
- Liao, Z.; Gong, Y.; Kai, G.; Zuo, K.; Chen, M.; Tan, Q.; Wei, Y.; Guo, L.; Tan, F.; Sun, X.; et al. An intron-free methyl jasmonate inducible geranylgeranyl diphosphate synthase gene from Taxus media and its functional identification in yeast. Mol. Biol. 2005, 39, 14–20. [Google Scholar] [CrossRef]
- Ruppel, N.J.; Kropp, K.N.; Davis, P.A.; Martin, A.E.; Luesse, D.R.; Hangarter, R.P. Mutations in GERANYLGERANYL DIPHOSPHATE SYNTHASE 1 affect chloroplast development in Arabidopsis thaliana (Brassicaceae). Am. J. Bot. 2013, 100, 2074–2084. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Zhang, M.; Song, S.; Wei, F.; Qin, L.; Fan, P.; Shi, Y.; Wang, X.; Wang, R. A Small Subunit of Geranylgeranyl Diphosphate Synthase Functions as an Active Regulator of Carotenoid Synthesis in Nicotiana tabacum. Int. J. Mol. Sci. 2023, 24, 992. [Google Scholar] [CrossRef]
- Xie, Q.; Ma, J.; Ding, G.; Yuan, B.; Wang, Y.; He, L.; Han, Y.; Cao, A.; Li, R.; Zhang, W.; et al. Transcriptomics and proteomics profiles of Taraxacum kok-saghyz roots revealed different gene and protein members play different roles for natural rubber biosynthesis. Ind. Crop. Prod. 2022, 181, 114776. [Google Scholar] [CrossRef]
- Lin, T.; Xu, X.; Du, H.; Fan, X.; Chen, Q.; Hai, C.; Zhou, Z.; Su, X.; Kou, L.; Gao, Q.; et al. Extensive sequence divergence between the reference genomes of Taraxacum kok-saghyz and Taraxacum mongolicum. Sci. China Life Sci. 2022, 65, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Liu, H.; Xu, Y.; Nie, Y. Entering an Era of Protein Structuromics. Biochemistry 2023, 62, 3167–3169. [Google Scholar] [CrossRef]
- Bailey, T.L. Discovering sequence motifs. Methods Mol. Biol. 2008, 452, 231–251. [Google Scholar] [CrossRef]
- Roy, S.W.; Gilbert, W. The evolution of spliceosomal introns: Patterns, puzzles and progress. Nat. Rev. Genet. 2006, 7, 211–221. [Google Scholar] [CrossRef]
- Nagel, R.; Bernholz, C.; Vranová, E.; Košuth, J.; Bergau, N.; Ludwig, S.; Wessjohann, L.; Gershenzon, J.; Tissier, A.; Schmidt, A. Arabidopsis thaliana isoprenyl diphosphate synthases produce the C25 intermediate geranylfarnesyl diphosphate. Plant J. Cell Mol. Biol. 2015, 84, 847–859. [Google Scholar] [CrossRef]
- Liang, P.H. Reaction kinetics, catalytic mechanisms, conformational changes, and inhibitor design for prenyltransferases. Biochemistry 2009, 48, 6562–6570. [Google Scholar] [CrossRef]
- Kapli, P.; Yang, Z.; Telford, M.J. Phylogenetic tree building in the genomic age. Nat. Rev. Genet. 2020, 21, 428–444. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of Gene Duplication in Plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Madrigal, P.; Tarazona, S.; Gomez-Cabrero, D.; Cervera, A.; McPherson, A.; Szcześniak, M.W.; Gaffney, D.J.; Elo, L.L.; Zhang, X.; et al. A survey of best practices for RNA-seq data analysis. Genome Biol. 2016, 17, 13. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Xu, X.; Ruan, J.; Liu, S.; Wu, S.; Shao, X.; Wang, X.; Gan, L.; Qin, B.; Yang, Y.; et al. Genome analysis of Taraxacum kok-saghyz Rodin provides new insights into rubber biosynthesis. J. Natl. Sci. Rev. 2018, 5, 78–87. [Google Scholar] [CrossRef]
- Stubbe, J.; Tian, J.; He, A.; Sinskey, A.J.; Lawrence, A.G.; Liu, P. Nontemplate-dependent polymerization processes: Polyhydroxyalkanoate synthases as a paradigm. Annu. Rev. Biochem. 2005, 74, 433–480. [Google Scholar] [CrossRef]
- Wu, X.; Chen, Y.; Shi, X.; Qi, K.; Cao, P.; Liu, X.; Yin, H.; Zhang, S. Effects of palmitic acid (16:0), hexacosanoic acid (26:0), ethephon and methyl jasmonate on the cuticular wax composition, structure and expression of key gene in the fruits of three pear cultivars. Funct. Plant Biol. FPB 2020, 47, 156–169. [Google Scholar] [CrossRef]
- Marand, A.P.; Eveland, A.L.; Kaufmann, K.; Springer, N.M. cis-Regulatory Elements in Plant Development, Adaptation, and Evolution. Annu. Rev. Plant Biol. 2023, 74, 111–137. [Google Scholar] [CrossRef]
- Dong, G.; Wang, H.; Qi, J.; Leng, Y.; Huang, J.; Zhang, H.; Yan, J. Transcriptome analysis of Taraxacum kok-saghyz reveals the role of exogenous methyl jasmonate in regulating rubber biosynthesis and drought tolerance. Gene 2023, 867, 147346. [Google Scholar] [CrossRef]
- Zhang, Z.; Shen, G.; Yang, Y.; Li, C.; Chen, X.; Yang, X.; Guo, X.; Miao, J.; Li, L.; Lei, M. Metabolic and Transcriptomic Analyses Reveal the Effects of Ethephon on Taraxacum kok-saghyz Rodin. Molecules 2022, 27, 3548. [Google Scholar] [CrossRef]
- Wang, Y.; Miao, Z.; Tang, K. Molecular cloning and functional expression analysis of a new gene encoding geranylgeranyl diphosphate synthase from hazel (Corylus avellana L. Gasaway). Mol. Biol. Rep. 2010, 37, 3439–3444. [Google Scholar] [CrossRef]
- Barja, M.V.; Rodriguez-Concepcion, M. Plant geranylgeranyl diphosphate synthases: Every (gene) family has a story. aBIOTECH 2021, 2, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, M.; Römer, S.; Suire, C.; Hugueney, P.; Weil, J.H.; Schantz, R.; Camara, B. Identification of a cDNA for the plastid-located geranylgeranyl pyrophosphate synthase from Capsicum annuum: Correlative increase in enzyme activity and transcript level during fruit ripening. Plant J. Cell Mol. Biol. 1992, 2, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Saito, T.; Nakagawa, T.; Kawamukai, M.; Kamiya, Y. Five geranylgeranyl diphosphate synthases expressed in different organs are localized into three subcellular compartments in Arabidopsis. Plant Physiol. 2000, 122, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- You, M.K.; Lee, Y.J.; Yu, J.S.; Ha, S.H. The Predicted Functional Compartmentation of Rice Terpenoid Metabolism by Trans-Prenyltransferase Structural Analysis, Expression and Localization. Int. J. Mol. Sci. 2020, 21, 8927. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, H.; Zong, Y.; Tu, Z.; Li, H. Isolation, expression, and functional analysis of the geranylgeranyl pyrophosphate synthase (GGPPS) gene from Liriodendron tulipifera. Plant Physiol. Biochem. 2021, 166, 700–711. [Google Scholar] [CrossRef]
- Gomez, J. Comparative ultracytology of young and mature latex vessels in Hevea brasiliensis; Rubber Research Institute of Malaysia: Kuala Lumput, Malaysia, 1975; Volume 2, pp. 143–164. [Google Scholar]
- Wang, C.; Wang, X.; Zhou, P.; Li, C. Genome-Wide Identification and Characterization of RdHSP Genes Related to High Temperature in Rhododendron delavayi. Plants 2024, 13, 1878. [Google Scholar] [CrossRef]
- Xu, Y.; Cheng, J.; Hu, H.; Yan, L.; Jia, J.; Wu, B. Genome-Wide Identification of NAC Family Genes in Oat and Functional Characterization of AsNAC109 in Abiotic Stress Tolerance. Plants 2024, 13, 1017. [Google Scholar] [CrossRef]
- Chen, M.; Ma, Y.; Wu, S.; Zheng, X.; Kang, H.; Sang, J.; Xu, X.; Hao, L.; Li, Z.; Gong, Z.; et al. Genome Warehouse: A Public Repository Housing Genome-scale Data. Genom. Proteom. Bioinform. 2021, 19, 584–589. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16, 157. [Google Scholar] [CrossRef]
- Yang, M.; Derbyshire, M.K.; Yamashita, R.A.; Marchler-Bauer, A. NCBI’s Conserved Domain Database and Tools for Protein Domain Analysis. Curr. Protoc. Bioinform. 2020, 69, e90. [Google Scholar] [CrossRef] [PubMed]
- Sonnhammer, E.L.; Eddy, S.R.; Durbin, R. Pfam: A comprehensive database of protein domain families based on seed alignments. Proteins 1997, 28, 405–420. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef]
- El-Rami, F.E.; Sikora, A.E. Bioinformatics Workflow for Gonococcal Proteomics. Methods Mol. Biol. (Clifton N.J.) 2019, 1997, 185–205. [Google Scholar] [CrossRef]
- Chou, K.C.; Shen, H.B. Cell-PLoc: A package of Web servers for predicting subcellular localization of proteins in various organisms. Nat. Protoc. 2008, 3, 153–162. [Google Scholar] [CrossRef]
- Geourjon, C.; Deléage, G. SOPM: A self-optimized method for protein secondary structure prediction. Protein Eng. 1994, 7, 157–164. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, 2, 2.3.1–2.3.22. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- He, Z.; Zhang, H.; Gao, S.; Lercher, M.J.; Chen, W.H.; Hu, S. Evolview v2: An online visualization and management tool for customized and annotated phylogenetic trees. Nucleic Acids Res. 2016, 44, W236–W241. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Collins-Silva, J.; Nural, A.T.; Skaggs, A.; Scott, D.; Hathwaik, U.; Woolsey, R.; Schegg, K.; McMahan, C.; Whalen, M.; Cornish, K.; et al. Altered levels of the Taraxacum kok-saghyz (Russian dandelion) small rubber particle protein, TkSRPP3, result in qualitative and quantitative changes in rubber metabolism. Phytochemistry 2012, 79, 46–56. [Google Scholar] [CrossRef]
- Chen, J.; Yang, Y.; Li, C.; Chen, Q.; Liu, S.; Qin, B. Genome-Wide Identification of MADS-Box Genes in Taraxacum kok-saghyz and Taraxacum mongolicum: Evolutionary Mechanisms, Conserved Functions and New Functions Related to Natural Rubber Yield Formation. Int. J. Mol. Sci. 2023, 24, 10997. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, Research0034. [Google Scholar] [CrossRef]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2(-△△CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Burks, D.J.; Azad, R.K. RNA-Seq Data Analysis Pipeline for Plants: Transcriptome Assembly, Alignment, and Differential Expression Analysis. Methods Mol. Biol. 2022, 2396, 47–60. [Google Scholar] [CrossRef]
- Franco, M.; Vivo, J.M. Cluster Analysis of Microarray Data. Methods Mol. Biol. 2019, 1986, 153–183. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Liu, X.; Su, H.; Li, M.; Li, M.; Wei, J. Regulatory Networks of Flowering Genes in Angelica sinensis during Vernalization. Plants 2022, 11, 1355. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).