
Arabidopsis Leaf Chloroplasts Have a Specific Sphingolipidome

Abstract
:1. Introduction
2. Results
2.1. Purification of Chloroplast Fractions
2.2. Overall Assessment of Sphingolipid Profiles
2.3. Distribution of Sphingolipid Classes
2.4. LCB and FA Profiling of Sphingolipids
2.5. Predominant Sphingolipid Species
3. Discussion
4. Materials and Methods
4.1. Plants and Growth Conditions
4.2. Chloroplast Isolation
4.3. Protein Quantification and Immunoblotting Assays
4.4. Sphingolipid Analysis
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Heaver, S.L.; Johnson, E.L.; Ley, R.E. Sphingolipids in host-microbial interactions. Curr. Opin. Microbiol. 2018, 43, 92–99. [Google Scholar] [CrossRef]
- Sperling, P.; Heinz, E. Plant sphingolipids: Structural diversity, biosynthesis, first genes and functions. Biochim. Biophys. Acta 2003, 1632, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Grison, M.S.; Brocard, L.; Fouillen, L.; Nicolas, W.; Wewer, V.; Dörmann, P.; Nacir, H.; Benitez-Alfonso, Y.; Claverol, S.; Germain, V.; et al. Specific membrane lipid composition is important for plasmodesmata function in Arabidopsis. Plant Cell 2015, 27, 1228–1250. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; van Meer, G. Lipid sorting in epithelial cells. Biochemistry 1988, 27, 6197–6202. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.K.; Tanaka, T.; Duke, S.O.; Porter, J.K.; Wray, E.M.; Hodges, L.; Sessions, A.E.; Wang, E.; Merrill, A.H., Jr. Fumonisin-and AAL-toxin-induced disruption of sphingolipid metabolism with accumulation of free sphingolipid bases. Plant Physiol. 1994, 106, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.K.; Carr, K.; Mcainsh, M.R.; Powell, B.; Hetherington, A.M. Drought-induced guard cell signal transduction involves sphingosine-1-phosphate. Nature 2001, 410, 596–599. [Google Scholar] [CrossRef] [PubMed]
- Msanne, J.; Chen, M.; Luttgeharm, K.D.; Bradley, A.M.; Mays, E.S.; Paper, J.M.; Boyle, D.L.; Cahoon, R.E.; Schrick, K.; Cahoon, E.B. Glucosylceramides are critical for cell-type differentiation and organogenesis, but not for cell viability in Arabidopsis. Plant J. 2015, 84, 188–201. [Google Scholar] [CrossRef] [PubMed]
- Liao, P.; Huang, J.; Tong, P.; Nie, W.; Yan, X.; Feng, Y.; Peng, H.; Peng, X.; Li, S. Characterization and expression analysis of inositolphosphorylceramide synthase family genes in rice (Oryza sativa L.). Genes Genom. 2017, 39, 485–492. [Google Scholar] [CrossRef]
- Liu, N.J.; Zhang, T.; Liu, Z.H.; Chen, X.; Guo, H.S.; Ju, B.H.; Zhang, Y.Y.; Li, G.Z.; Zhou, Q.H.; Qin, Y.M.; et al. Phytosphinganine affects plasmodesmata permeability via facilitating PDLP5-stimulated callose accumulation in Arabidopsis. Mol. Plant 2020, 13, 128–143. [Google Scholar] [CrossRef] [PubMed]
- Moore, W.M.; Chan, C.; Ishikawa, T.; Rennie, E.A.; Wipf, H.M.; Benites, V.; Kawai-Yamada, M.; Mortimer, J.C.; Scheller, H.V. Reprogramming sphingolipid glycosylation is required for endosymbiont persistence in Medicago truncatula. Curr. Biol. 2021, 31, 2374–2385. [Google Scholar] [CrossRef] [PubMed]
- Markham, J.E.; Li, J.; Cahoon, E.B.; Jaworski, J.G. Separation and identification of major plant sphingolipid classes from leaves. J. Biol. Chem. 2006, 281, 22684–22694. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.J.; Hou, L.P.; Bao, J.J.; Wang, L.J.; Chen, X.Y. Sphingolipid metabolism, transport, and functions in plants: Recent progress and future perspectives. Plant Commun. 2021, 2, 100214. [Google Scholar] [CrossRef]
- Luttgeharm, K.D.; Kimberlin, A.N.; Cahoon, E.B. Plant Sphingolipid Metabolism and Function. In Lipids in Plant and Algae Development; Nakamura, Y., Libeisson, Y., Eds.; Springer: Cham, Switzerland, 2016; Volume 86, pp. 249–286. [Google Scholar] [CrossRef]
- Markham, J.E.; Jaworski, J.G. Rapid measurement of sphingolipids from Arabidopsis thaliana by reversed-phase high-performance liquid chromatography coupled to electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 1304–1314. [Google Scholar] [CrossRef] [PubMed]
- Huby, E.; Napier, J.A.; Baillieul, F.; Michaelson, L.V.; Dhondt-Cordelier, S. Sphingolipids: Towards an integrated view of metabolism during the plant stress response. New Phytol. 2020, 225, 659–670. [Google Scholar] [CrossRef]
- Zeng, H.; Yao, N. Sphingolipids in plant immunity. Phytopathol. Res. 2022, 4, 20. [Google Scholar] [CrossRef]
- Sperling, P.; Franke, S.; Lüthje, S.; Heinz, E. Are glucocerebrosides the predominant sphingolipids in plant plasma membranes? Plant Physiol. Biochem. 2005, 43, 1031–1038. [Google Scholar] [CrossRef]
- Tarazona, P.; Feussner, K.; Feussner, I. An enhanced plant lipidomics method based on multiplexed liquid chromatography-mass spectrometry reveals additional insights into cold- and drought-induced membrane remodeling. Plant J. 2015, 84, 621–633. [Google Scholar] [CrossRef]
- Melser, S.; Batailler, B.; Peypelut, M.; Poujol, C.; Bellec, Y.; Wattelet-Boyer, V.; Maneta-Peyret, L.; Faure, J.D.; Moreau, P. Glucosylceramide biosynthesis is involved in Golgi morphology and protein secretion in plant cells. Traffic 2010, 11, 479–490. [Google Scholar] [CrossRef]
- Fouillen, L.; Maneta-Peyret, L.; Moreau, P. ER membrane lipid composition and metabolism: Lipidomic analysis. Methods Mol. Biol. 2018, 1691, 125–137. [Google Scholar] [CrossRef]
- Warnecke, D.; Heinz, E. Recently discovered functions of glucosylceramides in plants and fungi. Cell. Mol. Life Sci. 2003, 60, 919–941. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Li, J.; Gong, B.; Wang, R.H.; Chen, Y.L.; Yin, J.; Yang, C.; Lin, J.T.; Liu, H.Z.; Yang, Y.; et al. Orosomucoid proteins limit endoplasmic reticulum stress in plants. New Phyto. 2023, 240, 1134–1148. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Salazar, L.; Cahoon, R.E.; Gasca-Pineda, J.; Gonzalez-Solis, A.; Vera-Estrella, R.; Trevino, V.; Cahoon, E.B.; Gavilanes-Ruiz, M. Plasma and vacuolar membrane sphingolipidomes: Composition and insights on the role of main molecular species. Plant Physiol. 2021, 186, 624–639. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Senkler, J.; Herrfurth, C.; Braun, H.; Feussner, I. Defining the lipidome of Arabidopsis leaf mitochondria: Specific lipid complement and biosynthesis capacity. Plant Physiol. 2023, 191, 2185–2203. [Google Scholar] [CrossRef]
- Hölzl, G.; Dörmann, P. Chloroplast lipids and their biosynthesis. Annu. Rev. Plant Biol. 2019, 70, 51–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Benning, C. Chloroplast lipid synthesis and lipid trafficking through ER-plastid membrane contact sites. Biochem. Soc. Trans. 2012, 40, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Buré, C.; Cacas, J.L.; Wang, F.; Gaudin, K.; Domergue, F.; Mongrand, S.; Schmitter, J.M. Fast screening of highly glycosylated plant sphingolipids by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2011, 25, 3131–3145. [Google Scholar] [CrossRef]
- Chen, M.; Markham, J.E.; Cahoon, E.B. Sphingolipid Δ8 unsaturation is important for glucosylceramide biosynthesis and low-temperature performance in Arabidopsis. Plant J. 2012, 69, 769–781. [Google Scholar] [CrossRef]
- Lenarčič, T.; Albert, I.; Böhm, H.; Hodnik, V.; Pirc, K.; Zavec, A.B.; Podobnik, M.; Pahovnik, D.; Žagar, E.; Pruitt, R.; et al. Eudicot plant-specific sphingolipids determine host selectivity of microbial NLP cytolysins. Science 2017, 358, 1431–1434. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhou, X.; Tao, M.; Yuan, F.; Liu, L.; Wu, F.; Wu, X.; Xiang, Y.; Niu, Y.; Liu, F.; et al. Plant cell-surface GIPC sphingolipids sense salt to trigger Ca2+ influx. Nature 2019, 572, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Nagano, M.; Ishikawa, T.; Ogawa, Y.; Iwabuchi, M.; Nakasone, A.; Shimamoto, K.; Uchimiya, H.; Kawai-Yamada, M. Arabidopsis Bax inhibitor-1 promotes sphingolipid synthesis during cold stress by interacting with ceramide-modifying enzymes. Planta 2014, 240, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Ohlrogge, J.; Pollard, M.; Bao, X.; Focke, M.; Girke, T.; Ruuska, S.; Mekhedov, S.; Benning, C. Fatty acid synthesis: From CO2 to functional genomics. Biochem. Soc. Trans. 2000, 28, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Rawsthorne, S. Carbon flux and fatty acid synthesis in plants. Prog. Lipid Res. 2002, 41, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Berkey, R.; Bendigeri, D.; Xiao, S. Sphingolipids and plant defense/disease: The “death” connection and beyond. Front. Plant Sci. 2012, 3, 68. [Google Scholar] [CrossRef] [PubMed]
- Haslam, T.M.; Feussner, I. Diversity in sphingolipid metabolism across land plants. J. Exp. Bot. 2022, 73, 2785–2798. [Google Scholar] [CrossRef] [PubMed]
- Cha, H.J.; He, C.; Zhao, H.; Dong, Y.; An, I.S.; An, S. Intercellular and intracellular functions of ceramides and their metabolites in skin (Review). Int. J. Mol. Med. 2016, 38, 16–22. [Google Scholar] [CrossRef]
- Melser, S.; Molino, D.; Batailler, B.; Peypelut, M.; Laloi, M.; Wattelet-Boyer, V.; Bellec, Y.; Faure, J.D.; Moreau, P. Links between lipid homeostasis, organelle morphodynamics and protein trafficking in eukaryotic and plant secretory pathways. Plant Cell Rep. 2011, 30, 177–193. [Google Scholar] [CrossRef] [PubMed]
- Gronnier, J.; Germain, V.; Gouguet, P.; Cacas, J.L.; Mongrand, S. GIPC: Glycosyl Inositol Phospho Ceramides, the major sphingolipids on earth. Plant Signal. Behav. 2016, 11, e1152438. [Google Scholar] [CrossRef]
- Wattelet-Boyer, V.; Brocard, L.; Jonsson, K.; Esnay, N.; Joubès, J.; Domergue, F.; Mongrand, S.; Raikhel, N.; Bhalerao, R.P.; Moreau, P.; et al. Enrichment of hydroxylated C24- and C26-acyl-chain sphingolipids mediates PIN2 apical sorting at trans-Golgi network subdomains. Nat. Commun. 2016, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Wei, K.; Hu, K.; Tian, T.; Zhang, F.; Yu, Z.; Zhang, D.; Su, Y.; Sang, Y.; Zhang, X.; et al. MPK14-mediated auxin signaling controls lateral root development via ERF13-regulated very-long-chain fatty acid biosynthesis. Mol. Plant 2021, 14, 285–297. [Google Scholar] [CrossRef]
- Akolová, B.; Hudská, K.; Pullmannová, P.; Kováčik, A.; Palát, K.; Roh, J.; Fleddermann, J.; Estrela-Lopis, I.; Vávrová, K. Different phase behavior and packing of ceramides with long (C16) and very long (C24) acyls in model membranes: Infrared spectroscopy using deuterated lipids. J. Phys. Chem. B 2014, 118, 10460–10470. [Google Scholar] [CrossRef] [PubMed]
- Ternes, P.; Franke, S.; Zähringer, U.; Sperling, P.; Heinz, E. Identification and characterization of a sphingolipid delta 4-desaturase family. J. Biol. Chem. 2002, 277, 25512–25518. [Google Scholar] [CrossRef] [PubMed]
- Flores-Pérez, Ú.; Jarvis, P. Isolation and suborganellar fractionation of Arabidopsis chloroplasts. Methods Mol. Biol. 2017, 1511, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.Y.; Liu, Y.; Chen, D.K.; Bao, H.N.; Huang, L.Q.; Yin, J.; Chen, Y.L.; Xiao, S.; Yao, N. The immune components ENHANCED DISEASE SUSCEPTIBILITY 1 and PHYTOALEXIN DEFICIENT 4 are required for cell death caused by overaccumulation of ceramides in Arabidopsis. Plant J. 2021, 107, 1447–1465. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, H.Z.; Chen, D.K.; Zeng, H.Y.; Chen, Y.L.; Yao, N. PlantMetSuite: A user-friendly web-based tool for metabolomics analysis and visualisation. Plants-Basel 2023, 12, 2880. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cer | hCer | GlcCer | GIPC | Tissue | Ref. | |
|---|---|---|---|---|---|---|
| TL |  |  |  |  | Leaf | This study |
| MIC | 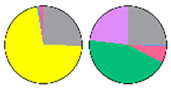 | 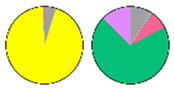 |  |  | Leaf | [23] |
| DRM |  |  |  |  | ||
| PM |  |  |  |  | ||
| VM |  |  |  |  | ||
| Mito |  |  |  |  | Leaf | [24] |
| Chloroplast |  |  |  |  | Leaf | This study |
| ER |  |  |  | / | Seedling | [22] |
| Nuclei |  |  |  | / | ||
| LCB ■t18:1 ■t18:0 ■d18:0 ■d18:1 ■d18:2 FA ■16C ■18C ■20C ■22C ■24C ■26C ■28C | ||||||
| Cer | hCer | GlcCer | GIPC | Tissue | Ref. | |
|---|---|---|---|---|---|---|
| TL | t18:1 c24:0 t18:1 c26:0 | t18:1 h24:0 t18:1 h24:1 | t18:1 h24:1 | t18:1 h24:0 t18:1 h24:1 | Leaf | This study |
| MIC | t18:1 c24:0 t18:1 c26:0 t18:1 c16:0 t18:1 c24:1 | t18:1 h24:1 t18:1 h24:0 | t18:1 h24:1 | t18:1 h24:0 t18:1 h24:1 | Leaf | [23] |
| DRM | t18:1 c24:0 t18:1 c26:0 | t18:1 h24:0 t18:1 h26:0 t18:1 h24:1 | d18:1 h16:0 | t18:1 h24:0 t18:0 h24:0 | ||
| PM | t18:1 c24:0 t18:1 c26:0 t18:0 c16:0 | t18:1 h24:1 t18:1 h24:0 | t18:1 h24:1 d18:1 h16:0 t18:1 h16:0 | t18:1 h24:1 t18:1 h24:0 | ||
| VM | t18:1 c24:0 t18:1 c26:0 t18:0 c24:0 | t18:1 h24:1 t18:1 h24:0 | t18:1 h24:1 d18:1 h16:0 t18:1 h16:0 | t18:1 h24:1 t18:1 h24:0 | ||
| Mito | d18:1 c24:0 t18:1 c26:0 | t18:0 h24:0 d18:1 h24:0 | d18:1 h24:1 | t18:1 h24:1 t18:1 h24:0 | Leaf | [24] |
| Chloroplast | t18:1 c24:0 t18:1 c26:0 | t18:1 h24:1 t18:1 h24:0 t18:1 h16:0 | d18:1 h16:0 t18:1 h16:0 | t18:1 h24:0 t18:1 h24:1 | Leaf | This study |
| ER | t18:0 c24:0 t18:1 c24:0 t18:1 c26:0 | t18:0 h24:0 t18:1 h24:0 t18:0 h26:1 | t18:1 h24:0 t18:1 h24:1 | / | Seedling | [22] |
| Nuclei | t18:0 c24:0 d18:1 c20:0 | t18:0 h26:1 t18:0 h24:0 | t18:1 h24:1 | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Wang, L.-Y.; Li, Y.-K.; Lin, J.-T.; Chen, D.-K.; Yao, N. Arabidopsis Leaf Chloroplasts Have a Specific Sphingolipidome. Plants 2024, 13, 299. https://doi.org/10.3390/plants13020299
Yang C, Wang L-Y, Li Y-K, Lin J-T, Chen D-K, Yao N. Arabidopsis Leaf Chloroplasts Have a Specific Sphingolipidome. Plants. 2024; 13(2):299. https://doi.org/10.3390/plants13020299
Chicago/Turabian StyleYang, Chang, Ling-Yan Wang, Yong-Kang Li, Jia-Ting Lin, Ding-Kang Chen, and Nan Yao. 2024. "Arabidopsis Leaf Chloroplasts Have a Specific Sphingolipidome" Plants 13, no. 2: 299. https://doi.org/10.3390/plants13020299