
Characterization of OsPIN2 Mutants Reveal Novel Roles for Reactive Oxygen Species in Modulating Not Only Root Gravitropism but Also Hypoxia Tolerance in Rice Seedlings


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
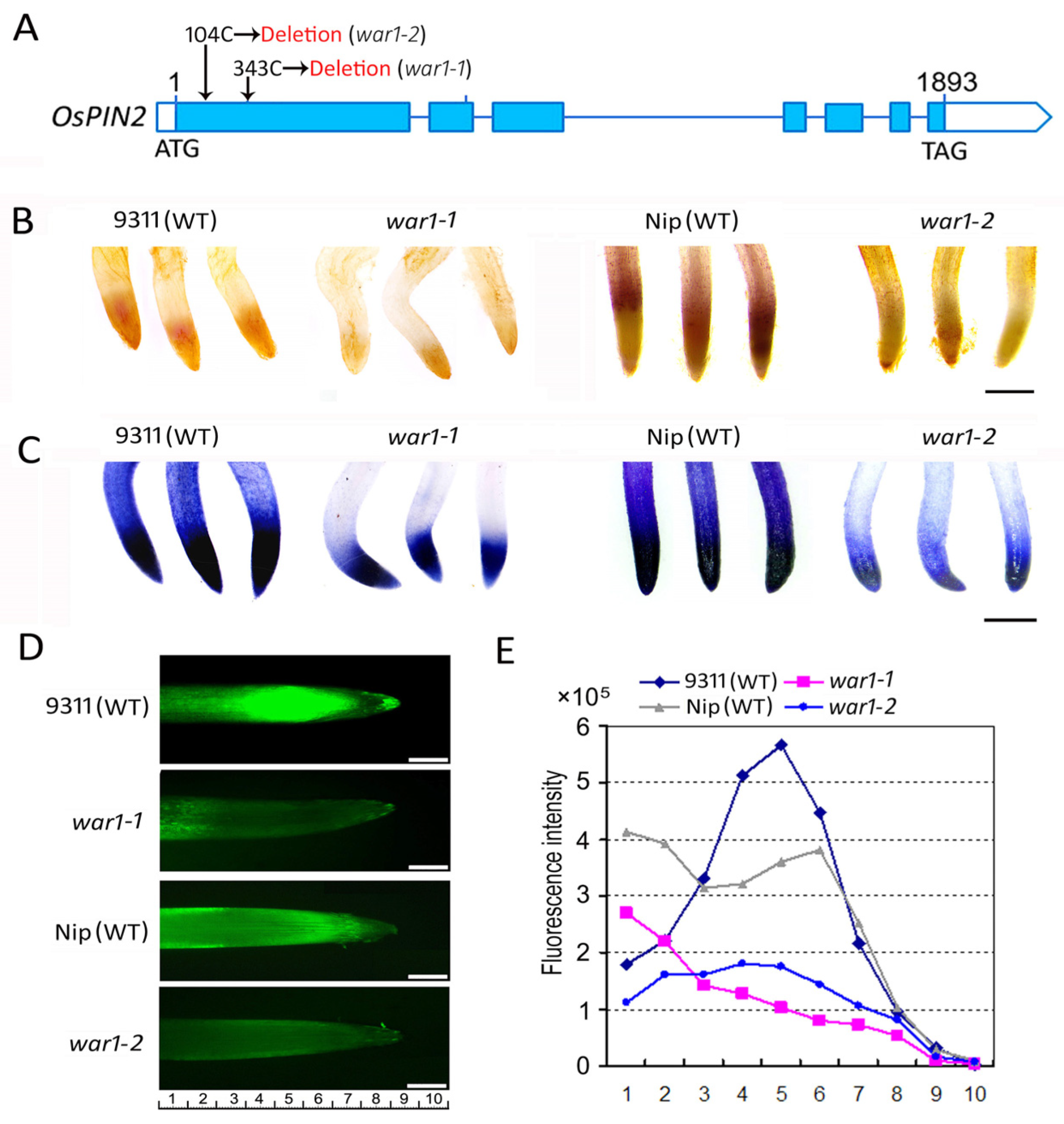
2.1. OsPIN2 Mutant Showed Lower ROS Accumulation in Root Tip
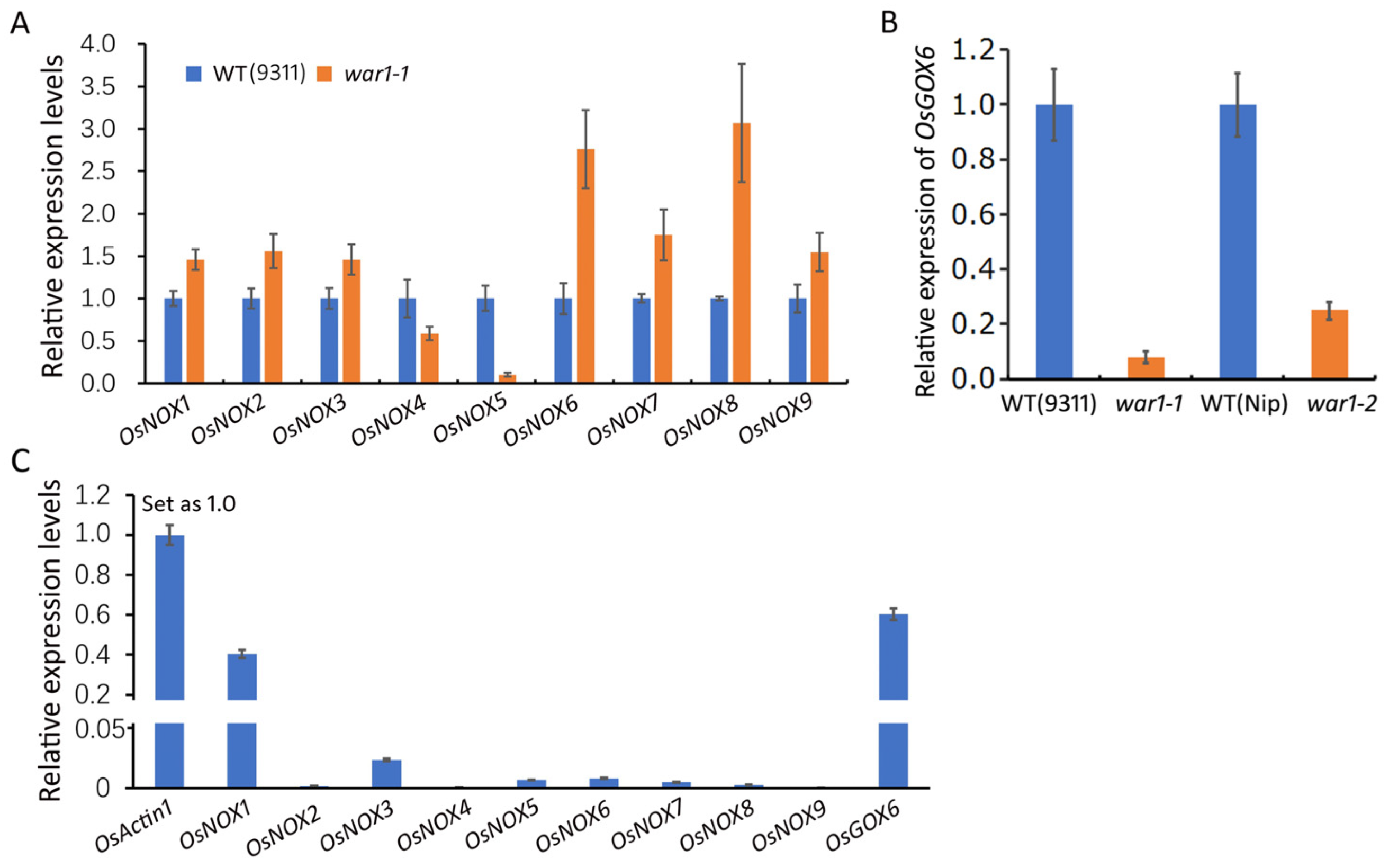
2.2. Expressions of ROS-Generating Genes in the Root of OsPIN2 Mutant
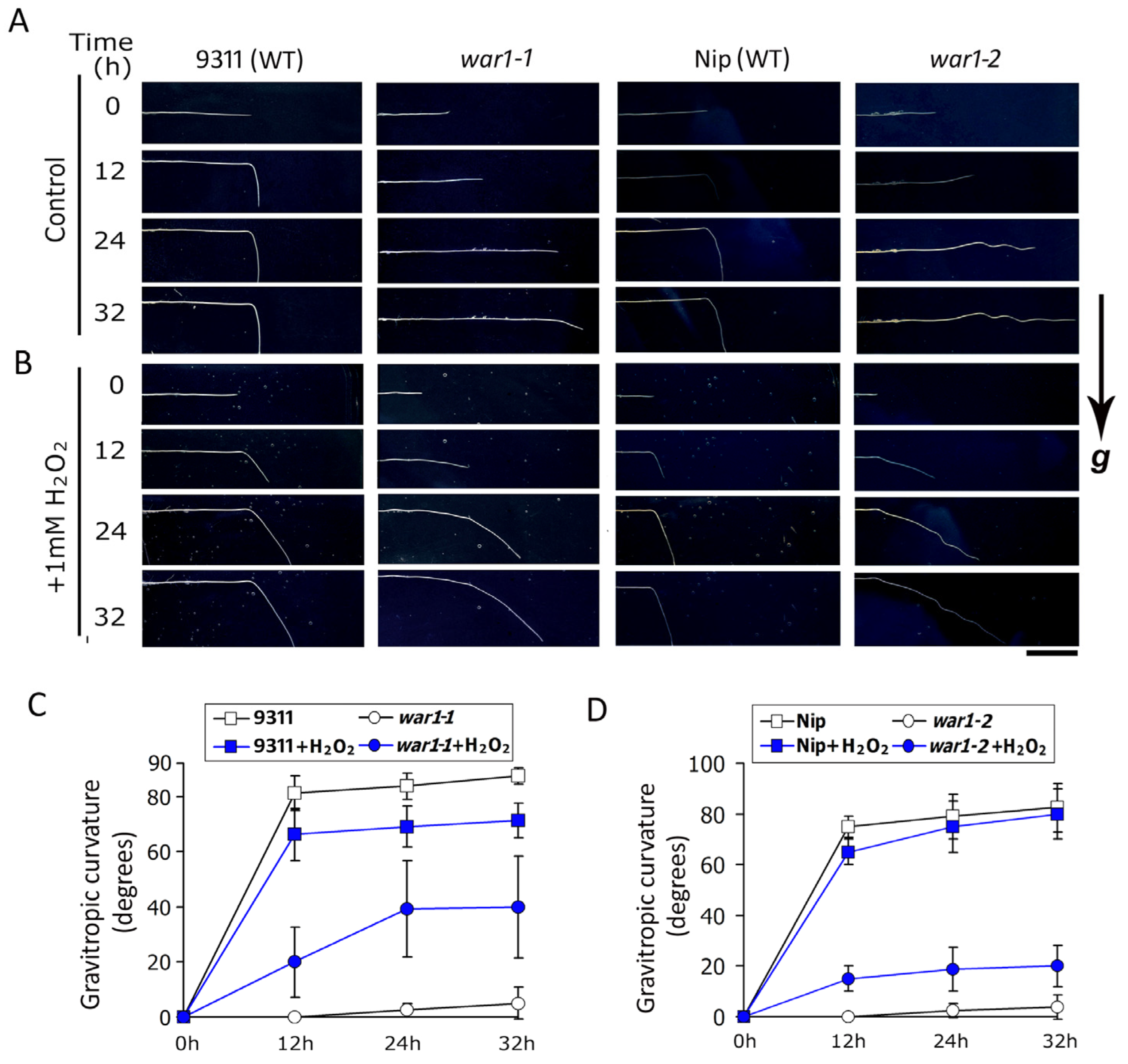
2.3. Application of Hydrogen Peroxide Induces Root Gravitropic Curvature in OsPIN2 Mutant
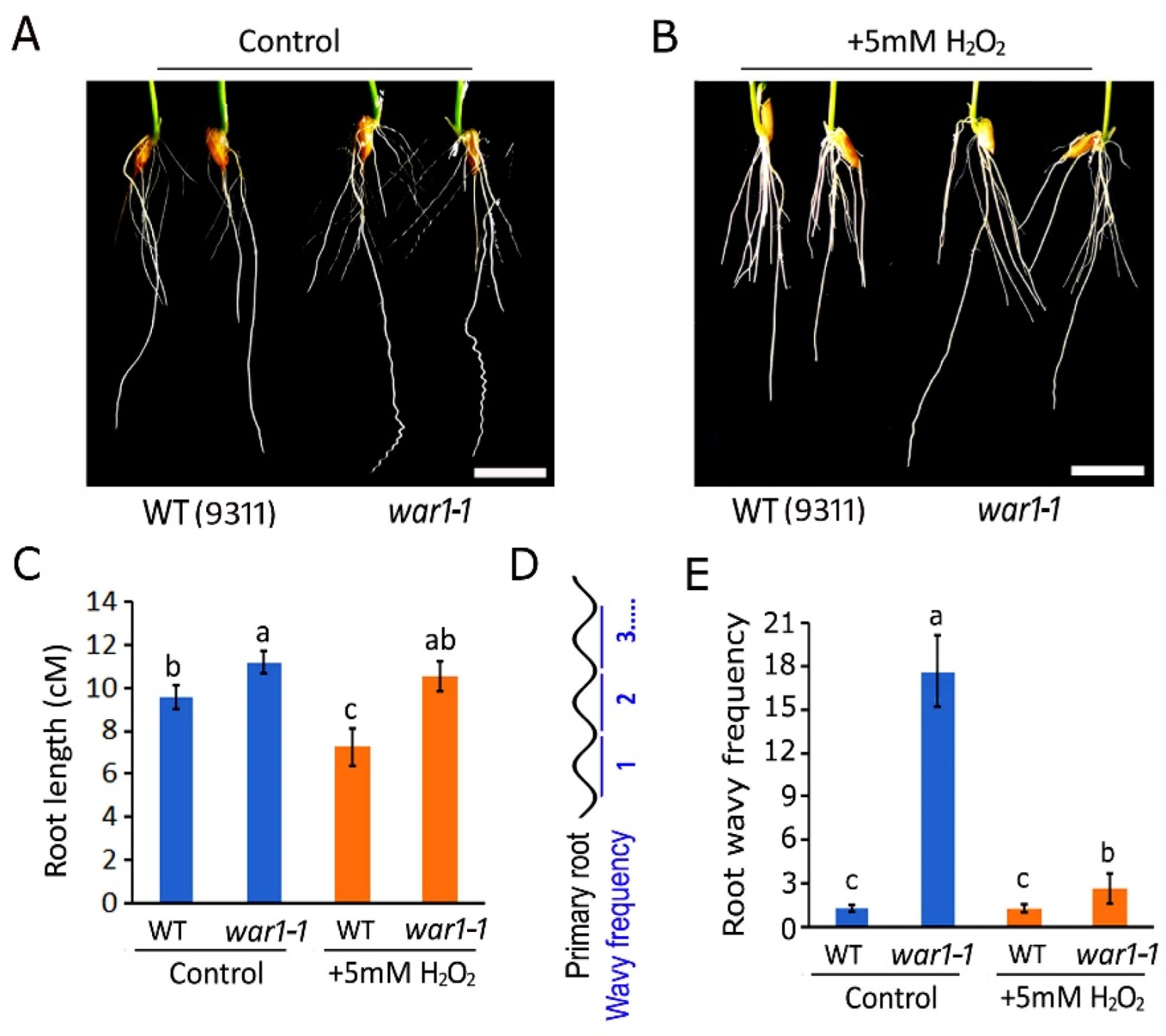
2.4. OsPIN2 Mutant Showed Increased Resistance to ROS Toxicity in Roots
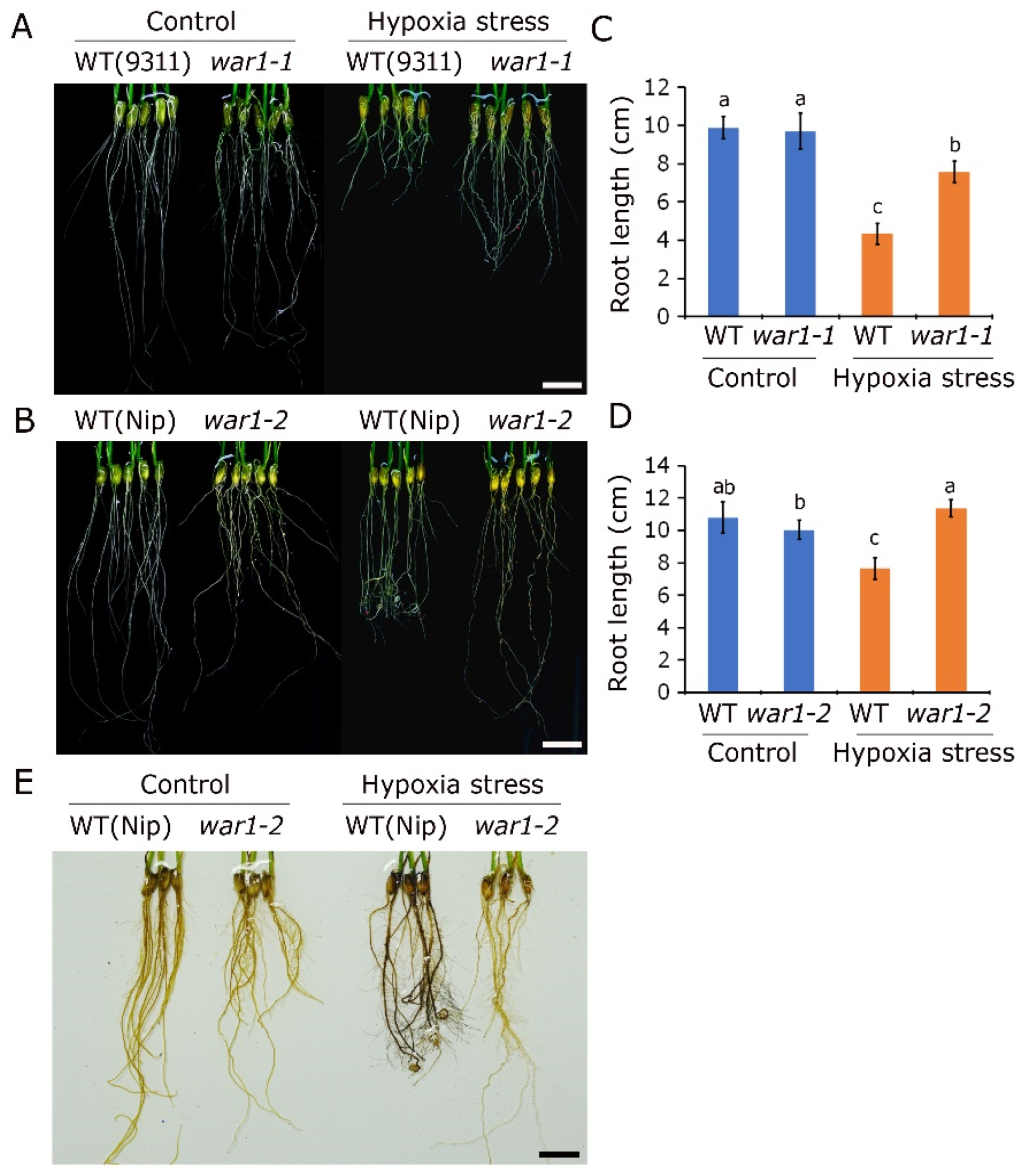
2.5. OsPIN2 Mutant Showed Lower ROS Accumulation and Stronger Tolerance under Hypoxic Stress
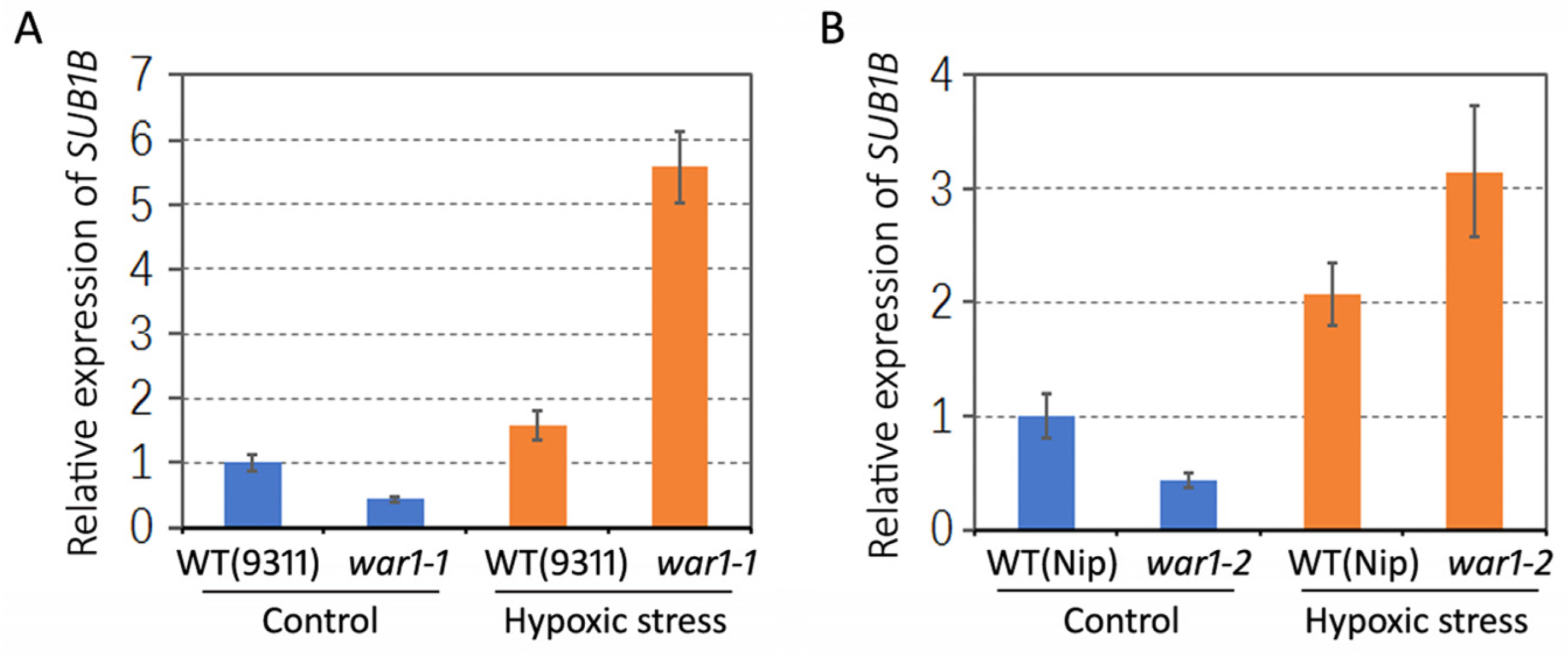
2.6. Differential Expression of Anoxic Resistance-Related Gene SUB1B in the Root of WT and OsPIN2 Mutant under Hypoxic Stress
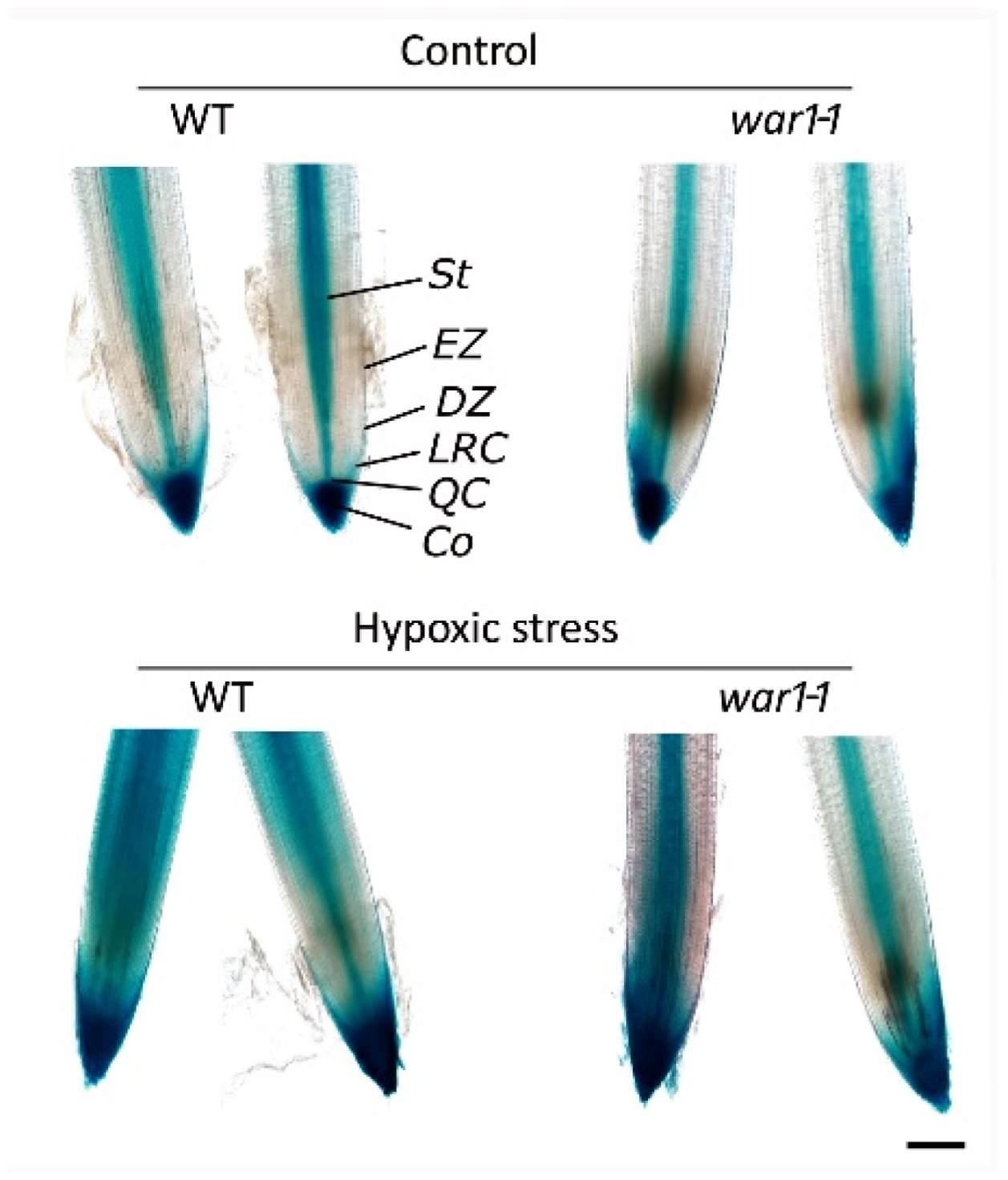
2.7. OsPIN2 Mutant Showed Reduced Auxin Distribution in the Root Tip under Hypoxia Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Hypoxic Stress Treatment
4.3. Detection of ROS Level in Roots
4.4. RNA Extraction and RT-qPCR Analysis
4.5. Detection of Auxin IAA Distribution by DR5 Promoter-GUS Experiment
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bakshi, A.; Gilroy, S. Analysis of plant flooding response. Methods Enzymol. 2023, 680, 461–491. [Google Scholar] [PubMed]
- Jethva, J.; Schmidt, R.R.; Sauter, M.; Selinski, J. Try or Die: Dynamics of Plant Respiration and How to Survive Low Oxygen Conditions. Plants 2022, 11, 205. [Google Scholar] [CrossRef]
- Yuan, L.B.; Chen, M.X.; Wang, L.N.; Sasidharan, R.; Voesenek, L.; Xiao, S. Multi-stress resilience in plants recovering from submergence. Plant Biotechnol. J. 2023, 21, 466–481. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.; Ali, S.; Park, S.; Bae, H. Exploring the Potential of Multiomics and Other Integrative Approaches for Improving Waterlogging Tolerance in Plants. Plants 2023, 12, 1544. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Seo, Y.S.; Walia, H.; Cao, P.; Fukao, T.; Canlas, P.E.; Amonpant, F.; Bailey-Serres, J.; Ronald, P.C. The submergence tolerance regulator Sub1A mediates stress-responsive expression of AP2/ERF transcription factors. Plant Physiol. 2010, 152, 1674–1692. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Waszczak, C.; Carmody, M.; Kangasjarvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [PubMed]
- Mhamdi, A.; Van Breusegem, F. Reactive oxygen species in plant development. Development 2018, 145, dev164376. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef]
- Ateeq, M.; Khan, A.H.; Zhang, D.; Alam, S.M.; Shen, W.; Wei, M.; Meng, J.; Shen, X.; Pan, J.; Zhu, K.; et al. Comprehensive physio-biochemical and transcriptomic characterization to decipher the network of key genes under waterlogging stress and its recuperation in Prunus persica. Tree Physiol. 2023, 43, 1265–1283. [Google Scholar] [CrossRef] [PubMed]
- Pucciariello, C.; Perata, P. The Oxidative Paradox in Low Oxygen Stress in Plants. Antioxidants 2021, 10, 332. [Google Scholar] [CrossRef]
- Tsukagoshi, H. Control of root growth and development by reactive oxygen species. Curr. Opin. Plant Biol. 2016, 29, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional regulation of ROS controls transition from proliferation to differentiation in the root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef]
- Kong, X.; Tian, H.; Yu, Q.; Zhang, F.; Wang, R.; Gao, S.; Xu, W.; Liu, J.; Shani, E.; Fu, C.; et al. PHB3 Maintains Root Stem Cell Niche Identity through ROS-Responsive AP2/ERF Transcription Factors in Arabidopsis. Cell Rep. 2018, 22, 1350–1363. [Google Scholar] [CrossRef] [PubMed]
- Mabuchi, K.; Maki, H.; Itaya, T.; Suzuki, T.; Nomoto, M.; Sakaoka, S.; Morikami, A.; Higashiyama, T.; Tada, Y.; Busch, W.; et al. MYB30 links ROS signaling, root cell elongation, and plant immune responses. Proc. Natl. Acad. Sci. USA 2018, 115, E4710–E4719. [Google Scholar] [CrossRef]
- Xia, X.J.; Zhou, Y.H.; Shi, K.; Zhou, J.; Foyer, C.H.; Yu, J.Q. Interplay between reactive oxygen species and hormones in the control of plant development and stress tolerance. J. Exp. Bot. 2015, 66, 2839–2856. [Google Scholar] [CrossRef]
- Qi, J.S.; Wang, J.L.; Gong, Z.Z.; Zhou, J.M. Apoplastic ROS signaling in plant immunity. Curr. Opin. Plant Biol. 2017, 38, 92–100. [Google Scholar] [CrossRef]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- Zafari, S.; Vanlerberghe, G.C.; Igamberdiev, A.U. The Role of Alternative Oxidase in the Interplay between Nitric Oxide, Reactive Oxygen Species, and Ethylene in Tobacco (Nicotiana tabacum L.) Plants Incubated under Normoxic and Hypoxic Conditions. Int. J. Mol. Sci. 2022, 23, 7153. [Google Scholar] [CrossRef]
- Liu, Z.; Hartman, S.; van Veen, H.; Zhang, H.; Leeggangers, H.; Martopawiro, S.; Bosman, F.; de Deugd, F.; Su, P.; Hummel, M.; et al. Ethylene augments root hypoxia tolerance via growth cessation and reactive oxygen species amelioration. Plant Physiol. 2022, 190, 1365–1383. [Google Scholar] [CrossRef]
- Gorb, S.N.; Sauter, M.; Kovalev, A.; Steffens, B. Emerging roots alter epidermal cell fate through mechanical and reactive oxygen species signaling. Plant Cell 2012, 24, 3296–3306. [Google Scholar]
- Ma, F.; Wang, L.; Li, J.; Samma, M.K.; Xie, Y.; Wang, R.; Wang, J.; Zhang, J.; Shen, W. Interaction between HY1 and H2O2 in auxin-induced lateral root formation in Arabidopsis. Plant Mol. Biol. 2014, 85, 49–61. [Google Scholar] [CrossRef]
- Wang, P.; Du, Y.; Li, Y.; Ren, D.; Song, C.P. Hydrogen peroxide-mediated activation of MAP kinase 6 modulates nitric oxide biosynthesis and signal transduction in Arabidopsis. Plant Cell 2010, 22, 2981–2998. [Google Scholar] [CrossRef]
- Peer, W.A.; Cheng, Y.; Murphy, A.S. Evidence of oxidative attenuation of auxin signalling. J. Exp. Bot. 2013, 64, 2629–2639. [Google Scholar] [CrossRef]
- Su, S.H.; Gibbs, N.M.; Jancewicz, A.L.; Masson, P.H. Molecular Mechanisms of Root Gravitropism. Curr. Biol. 2017, 27, R964–R972. [Google Scholar] [CrossRef]
- Sato, E.M.; Hijazi, H.; Bennett, M.J.; Vissenberg, K.; Swarup, R. New insights into root gravitropic signalling. J. Exp. Bot. 2015, 66, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Krieger, G.; Shkolnik, D.; Miller, G.; Fromm, H. Reactive Oxygen Species Tune Root Tropic Responses. Plant Physiol. 2016, 172, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Joo, J.H.; Bae, Y.S.; Lee, J.S. Role of auxin-induced reactive oxygen species in root gravitropism. Plant Physiol. 2001, 126, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guo, M.; Li, Y.; Ruan, W.; Mo, X.; Wu, Z.; Sturrock, C.J.; Yu, H.; Lu, C.; Peng, J.; et al. LARGE ROOT ANGLE1, encoding OsPIN2, is involved in root system architecture in rice. J. Exp. Bot. 2018, 69, 385–397. [Google Scholar] [CrossRef]
- Inahashi, H.; Shelley, I.J.; Yamauchi, T.; Nishiuchi, S.; Takahashi-Nosaka, M.; Matsunami, M.; Ogawa, A.; Noda, Y.; Inukai, Y. OsPIN2, which encodes a member of the auxin efflux carrier proteins, is involved in root elongation growth and lateral root formation patterns via the regulation of auxin distribution in rice. Physiol. Plant. 2018, 164, 216–225. [Google Scholar] [CrossRef]
- Li, W.; Zhang, M.; Qiao, L.; Chen, Y.; Zhang, D.; Jing, X.; Gan, P.; Huang, Y.; Gao, J.; Liu, W.; et al. Characterization of wavy root 1, an agravitropism allele, reveals the functions of OsPIN2 in fine regulation of auxin transport and distribution and in ABA biosynthesis and response in rice (Oryza sativa L.). Crop J. 2022, 10, 980–992. [Google Scholar] [CrossRef]
- Wang, G.-F.; Li, W.-Q.; Li, W.-Y.; Wu, G.-L.; Zhou, C.-Y.; Chen, K.-M. Characterization of rice NADPH oxidase genes and their expression under various environmental conditions. Int. J. Mol. Sci. 2013, 14, 9440–9458. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Challa, G.S.; Gupta, A.; Gu, L.; Wu, Y.; Li, W. Physiological and Transcriptomic Characterization of Sea-Wheatgrass-Derived Waterlogging Tolerance in Wheat. Plants 2022, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Muller, A.; Guan, C.; Galweiler, L.; Tanzler, P.; Huijser, P.; Marchant, A.; Parry, G.; Bennett, M.; Wisman, E.; Palme, K. AtPIN2 defines a locus of Arabidopsis for root gravitropism control. EMBO J. 1998, 17, 6903–6911. [Google Scholar] [CrossRef] [PubMed]
- Sukumar, P.; Edwards, K.S.; Rahman, A.; DeLong, A.; Muday, G.K. PINOID Kinase Regulates Root Gravitropism through Modulation of PIN2-Dependent Basipetal Auxin Transport in Arabidopsis. Plant Physiol. 2009, 150, 722–735. [Google Scholar] [CrossRef]
- Sinclair, W.; Trewavas, A.J. Calcium in gravitropism. A re-examination. Planta 1997, 203, S85–S90. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.T.; Feldman, L.J. Light-regulated root gravitropism: A role for, and characterization of, a calcium/calmodulin-dependent protein kinase homolog. Planta 1997, 203, S91–S97. [Google Scholar] [CrossRef] [PubMed]
- Perera, I.Y.; Hung, C.Y.; Brady, S.; Muday, G.K.; Boss, W.F. A universal role for inositol 1,4,5-trisphosphate-mediated signaling in plant gravitropism. Plant Physiol. 2006, 140, 746–760. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Neill, S.J.; Tang, Z.; Cai, W. Nitric oxide mediates gravitropic bending in soybean roots. Plant Physiol. 2005, 137, 663–670. [Google Scholar] [CrossRef]
- Joo, J.H.; Yoo, H.J.; Hwang, I.; Lee, J.S.; Nam, K.H.; Bae, Y.S. Auxin-induced reactive oxygen species production requires the activation of phosphatidylinositol 3-kinase. FEBS Lett. 2005, 579, 1243–1248. [Google Scholar] [CrossRef]
- Eljebbawi, A.; Rondon, Y.; Dunand, C.; Estevez, J. Highlighting Reactive Oxygen Species (ROS) as multitaskers in root development. iScience 2020, 24, 101978. [Google Scholar] [CrossRef] [PubMed]
- Orman-Ligeza, B.; Parizot, B.; de Rycke, R.; Fernandez, A.; Himschoot, E.; Van Breusegem, F.; Bennett, M.J.; Perilleux, C.; Beeckman, T.; Draye, X. RBOH-mediated ROS production facilitates lateral root emergence in Arabidopsis. Development 2016, 143, 3328–3339. [Google Scholar] [CrossRef] [PubMed]
- Ivanchenko, M.G.; den Os, D.; Monshausen, G.B.; Dubrovsky, J.G.; Bednarova, A.; Krishnan, N. Auxin increases the hydrogen peroxide (H2O2) concentration in tomato (Solanum lycopersicum) root tips while inhibiting root growth. Ann. Bot. 2013, 112, 1107–1116. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, Y.; Xie, Z.; Li, X.; He, Z.H.; Peng, X.X. Association-Dissociation of Glycolate Oxidase with Catalase in Rice: A Potential Switch to Modulate Intracellular H2O2 Levels. Mol. Plant 2016, 9, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Rojas, C.; Mysore, K.S. Glycolate oxidase is an alternative source for H2O2 production during plant defense responses and functions independently from NADPH oxidase. Plant Signal. Behav. 2012, 7, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, Z.; Zhang, K.; Zhang, X.; Zhang, Y.; Wang, X.; Han, C.; Yu, J.; Xu, K.; Li, D. Barley Stripe Mosaic Virus gammab Interacts with Glycolate Oxidase and Inhibits Peroxisomal ROS Production to Facilitate Virus Infection. Mol. Plant 2018, 11, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.S.; Lu, Y.S.; Zhai, L.G.; Deng, R.S.; Jiang, J.; Li, Y.; He, Z.H.; Peng, X.X. Glycolate Oxidase Isozymes Are Coordinately Controlled by GLO1 and GLO4 in Rice. PLoS ONE 2012, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Esser, C.; Kuhn, A.; Groth, G.; Lercher, M.J.; Maurino, V.G. Plant and animal glycolate oxidases have a common eukaryotic ancestor and convergently duplicated to evolve long-chain 2-hydroxy acid oxidases. Mol. Biol. Evol. 2014, 31, 1089–1101. [Google Scholar] [CrossRef]
- Yamada, M.; Higashiyama, T.; Kishino, S.; Kataoka, M.; Ogawa, J.; Shimizu, S.; Isobe, K. Novel alcohol oxidase with glycolate oxidase activity from Ochrobactrum sp. AIU 033. J. Mol. Catal. B-Enzym. 2014, 105, 41–48. [Google Scholar] [CrossRef]
- Williams, E.; Cregeen, D.; Rumsby, G. Identification and expression of a cDNA for human glycolate oxidase. Biochim. Biophys. Acta. 2000, 1493, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Pennati, A.; Gadda, G. Involvement of ionizable groups in catalysis of human liver glycolate oxidase. J. Biol. Chem. 2009, 284, 31214–31222. [Google Scholar] [CrossRef]
- Vignaud, C.; Pietrancosta, N.; Williams, E.L.; Rumsby, G.; Lederer, F. Purification and characterization of recombinant human liver glycolate oxidase. Arch. Biochem. Biophys. 2007, 465, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, J.; Hudig, M.; Meier, D.; Linka, N.; Maurino, V.G. The genome of Ricinus communis encodes a single glycolate oxidase with different functions in photosynthetic and heterotrophic organs. Planta 2020, 252, 100. [Google Scholar] [CrossRef] [PubMed]
- Engqvist, M.K.; Schmitz, J.; Gertzmann, A.; Florian, A.; Jaspert, N.; Arif, M.; Balazadeh, S.; Mueller-Roeber, B.; Fernie, A.R.; Maurino, V.G. GLYCOLATE OXIDASE3, a Glycolate Oxidase Homolog of Yeast l-Lactate Cytochrome c Oxidoreductase, Supports l-Lactate Oxidation in Roots of Arabidopsis. Plant Physiol. 2015, 169, 1042–1061. [Google Scholar] [CrossRef]
- Yu, X.L.; Wu, D.M.; Fu, Y.Q.; Yang, X.J.; Baluska, F.; Shen, H. OsGLO4 is involved in the formation of iron plaques on surface of rice roots grown under alternative wetting and drying condition. Plant Soil 2018, 423, 111–123. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Steffens, B.; Geske, T.; Sauter, M. Aerenchyma formation in the rice stem and its promotion by H2O2. New Phytol. 2011, 190, 369–378. [Google Scholar] [CrossRef]
- Steffens, B.; Sauter, M. Epidermal cell death in rice is confined to cells with a distinct molecular identity and is mediated by ethylene and H2O2 through an autoamplified signal pathway. Plant Cell 2009, 21, 184–196. [Google Scholar] [CrossRef]
- Gechev, T.S.; Hille, J. Hydrogen peroxide as a signal controlling plant programmed cell death. J. Cell Biol. 2005, 168, 17–20. [Google Scholar] [CrossRef]
- Houot, V.; Etienne, P.; Petitot, A.S.; Barbier, S.; Blein, J.P.; Suty, L. Hydrogen peroxide induces programmed cell death features in cultured tobacco BY-2 cells, in a dose-dependent manner. J. Exp. Bot. 2001, 52, 1721–1730. [Google Scholar] [PubMed]
- Niroula, R.K.; Pucciariello, C.; Ho, V.T.; Novi, G.; Fukao, T.; Perata, P. SUB1A-dependent and -independent mechanisms are involved in the flooding tolerance of wild rice species. Plant J. 2012, 72, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.K.; Callis, J.; Wemmer, D.; Walbot, V.; Jardetzky, O. Mechanisms of cytoplasmic pH regulation in hypoxic maize root tips and its role in survival under hypoxia. Proc. Natl. Acad. Sci. USA 1984, 81, 3379–3383. [Google Scholar] [CrossRef]
- Mira, M.M.; Hill, R.D.; Stasolla, C. Phytoglobins Improve Hypoxic Root Growth by Alleviating Apical Meristem Cell Death. Plant Physiol. 2016, 172, 2044–2056. [Google Scholar] [CrossRef]
- Mira, M.M.; El-Khateeb, E.A.; Gaafar, R.M.; Igamberdiev, A.U.; Hill, R.D.; Stasolla, C. Stem cell fate in hypoxic root apical meristems is influenced by phytoglobin expression. J. Exp. Bot. 2020, 71, 1350–1362. [Google Scholar] [CrossRef]
- Xu, Q.T.; Yang, L.; Zhou, Z.Q.; Mei, F.Z.; Qu, L.H.; Zhou, G.S. Process of aerenchyma formation and reactive oxygen species induced by waterlogging in wheat seminal roots. Planta 2013, 238, 969–982. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.M.; Wang, Y.J.; Gao, Y.T.; Li, R.; Wang, G.F.; Li, W.Q.; Liu, W.T.; Chen, K.M. The plasma membrane NADPH oxidase OsRbohA plays a crucial role in developmental regulation and drought-stress response in rice. Physiol. Plant. 2016, 156, 421–443. [Google Scholar] [CrossRef]
- Zhou, J.; Yu, F.; Wang, X.; Yang, Y.; Yu, C.; Liu, H.; Cheng, Y.; Yan, C.; Chen, J. Specific expression of DR5 promoter in rice roots using a tCUP derived promoter-reporter system. PLoS ONE 2014, 9, e87008. [Google Scholar] [CrossRef][Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, B.; Zhang, R.; Zhang, C.; Wen, N.; Xia, Y.; Zhao, Y.; Li, Q.; Qiao, L.; Li, W. Characterization of OsPIN2 Mutants Reveal Novel Roles for Reactive Oxygen Species in Modulating Not Only Root Gravitropism but Also Hypoxia Tolerance in Rice Seedlings. Plants 2024, 13, 476. https://doi.org/10.3390/plants13040476
Hao B, Zhang R, Zhang C, Wen N, Xia Y, Zhao Y, Li Q, Qiao L, Li W. Characterization of OsPIN2 Mutants Reveal Novel Roles for Reactive Oxygen Species in Modulating Not Only Root Gravitropism but Also Hypoxia Tolerance in Rice Seedlings. Plants. 2024; 13(4):476. https://doi.org/10.3390/plants13040476
Chicago/Turabian StyleHao, Bowen, Ruihan Zhang, Chengwei Zhang, Na Wen, Yu Xia, Yang Zhao, Qinying Li, Lei Qiao, and Wenqiang Li. 2024. "Characterization of OsPIN2 Mutants Reveal Novel Roles for Reactive Oxygen Species in Modulating Not Only Root Gravitropism but Also Hypoxia Tolerance in Rice Seedlings" Plants 13, no. 4: 476. https://doi.org/10.3390/plants13040476
APA StyleHao, B., Zhang, R., Zhang, C., Wen, N., Xia, Y., Zhao, Y., Li, Q., Qiao, L., & Li, W. (2024). Characterization of OsPIN2 Mutants Reveal Novel Roles for Reactive Oxygen Species in Modulating Not Only Root Gravitropism but Also Hypoxia Tolerance in Rice Seedlings. Plants, 13(4), 476. https://doi.org/10.3390/plants13040476