
Photosynthetic Responses of Racomitrium japonicum L. to Strontium Stress Evaluated through Chlorophyll a Fluorescence OJIP Transient Analysis
Abstract
:1. Introduction
2. Results
2.1. Effects of Sr2+ on Cell and Chlorophyll a Contents in R. japonicum
2.2. Effects of Sr2+ on Chl a Fluorescence Rise Kinetics in R. japonicum
2.2.1. Raw Fluorescence Rise Kinetic OJIP Curves and Relative Variable Fluorescence Vt
2.2.2. The L-Band
2.2.3. The K-Band
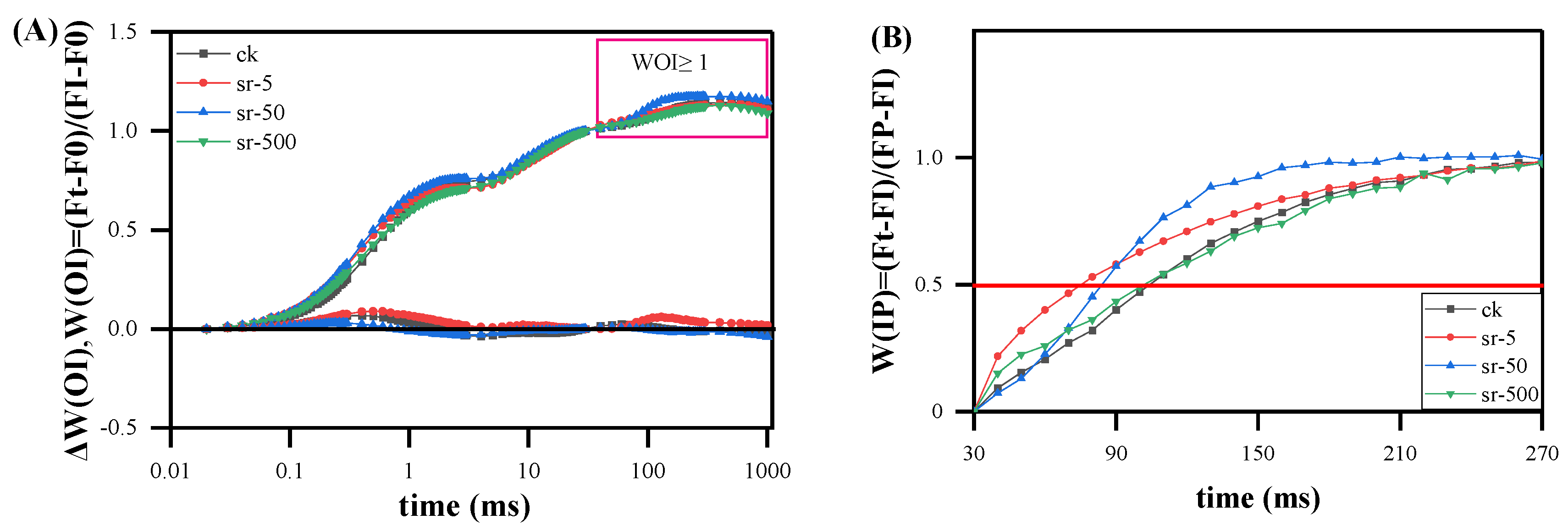
2.2.4. The IP Phase
2.3. The Energy Fluxes in R. japonicum
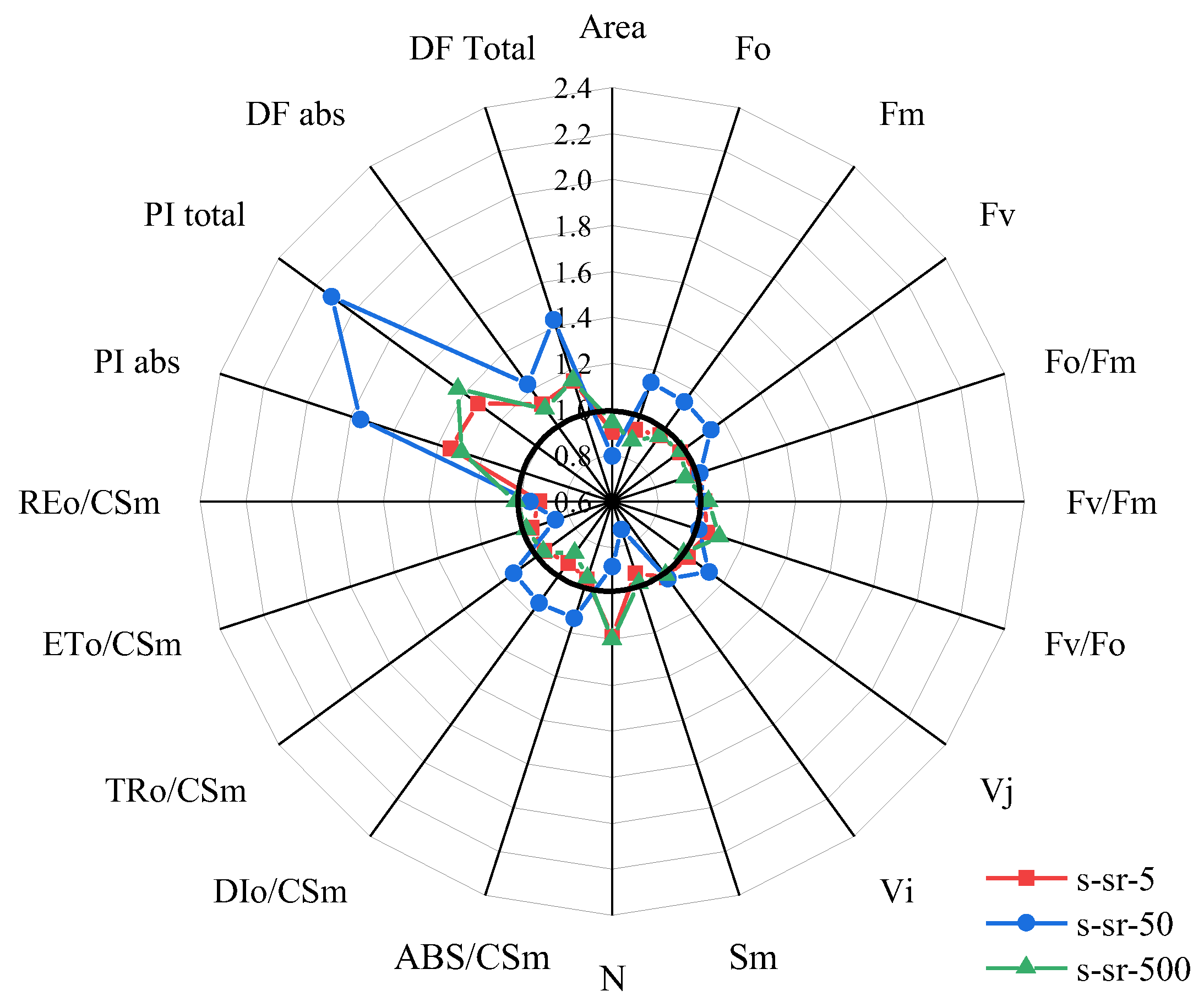
2.4. JIP-Test Analysis in R. japonicum
2.5. Photosynthetic Electron Transport in R. japonicum
3. Discussion
3.1. Chl a Fluorescence Rise Kinetic Response of Moss under Sr2+ Stress
3.2. Photosynthetic Organ Response of Mosses under Sr2+ Stress
3.3. Electron Transport Response of Moss under Sr2+ Stress
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Cell Structure Observation and Chlorophyll Content Determination
4.3. Chl a Fluorescence Kinetic Measurement and JIP Test
4.4. Data Analysis
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Baker, N.R.; Long, S.P. Photosynthesis in contrasting environments. Photosynth. Contrasting Environ. 1986, 7, 1. [Google Scholar]
- Baker, N.R.; Bowyer, J.R. Photoinhibition of photosynthesis: From molecular mechanisms to the field. BIOS Sci. 1994, 124, 156. [Google Scholar]
- Harborne, J.B. Physiological processes in plant ecology: Towards a synthesis of atriplex. Phytochemistry 1981, 20, 2449. [Google Scholar] [CrossRef]
- Yordanov, I.; Dilova, S.; Petkova, R.; Pangelova, T.; Zuss, K.H. Mechanisms of the temperature damage and acclimation of the photosynthetic apparatus. Photobiochem. Photobiophys. 1986, 12, 147–155. [Google Scholar]
- Xia, H.; Cheng, X.; Zheng, L.; Ren, H.; Li, W.; Lei, Y.; Plenković-Moraj, A.; Chen, K. Sex-Specific Physiological Responses of Populus cathayana to Uranium Stress. Forests 2022, 13, 1123. [Google Scholar] [CrossRef]
- Armond, P.A.; Björkman, O.; Staehelin, L.A. Dissociation of supramolecular complexes in chloroplast membranes A manifestation of heat damage to the photosynthetic apparatus. Biochim. Biophys. Acta (BBA)-Biomembr. 1980, 601, 433–442. [Google Scholar] [CrossRef]
- Gounaris, K.; Brain, A.; Quinn, P.; Williams, W. Structural reorganisation of chloroplast thylakoid membranes in response to heat-stress. Biochim. Biophys. Acta (BBA)-Bioenerg. 1984, 766, 198–208. [Google Scholar] [CrossRef]
- Jianrong, X.; Qiran, T. Early stage toxicity of excess copper to photosystem II of Chlorella pyrenoidosa–OJIP chlorophyll a fluorescence analysis. J. Environ. Sci. 2009, 21, 1569–1574. [Google Scholar]
- Jiang, Y.; Huang, R.; Jiang, L.; Chen, K.; Zhu, W. Alleviation of Cadmium Toxicity to Medicago Truncatula by AMF Involves the Changes of Cd Speciation in Rhizosphere Soil and Subcellular Distribution. Phyton 2021, 90, 403. [Google Scholar] [CrossRef]
- Dushenkov, S. Trends in phytoremediation of radionuclides. Plant Soil 2003, 249, 167–175. [Google Scholar] [CrossRef]
- Entry, J.A.; Vance, N.C.; Hamilton, M.A.; Zabowski, D.; Watrud, L.S.; Adriano, D.C. Phytoremediation of soil contaminated with low concentration of radionuclides. Water Air Soil Pollut. 1996, 664, 299–306. [Google Scholar] [CrossRef]
- Burger, A.; Lichtscheidl, I. Strontium in the environment: Review about reactions of plants towards stable and radioactive strontium isotopes. Sci. Total Environ. 2019, 653, 1458–1512. [Google Scholar] [CrossRef]
- Crompton, T.R. Determination of Metals in Natural Waters, Sediments and Soils; Metals in Natural Water Samples; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1–8. [Google Scholar]
- Sripathy, L.; Rao, P.; Kumar, N.M.A.; Yashwanth, S.; Sharada, K.R. Heavy metal contamination of soil due to vehicular traffic: A case study across nelamangala-dabaspet segment of national highway no. 4. Rasayan J. Chem. 2015, 8, 232–236. [Google Scholar]
- Bonari, G.; Monaci, F.; Nannoni, F.; Angiolini, C.; Protano, G. Trace Element Uptake and Accumulation in the Medicinal Herb Hypericum perforatum L. Across Different Geolithological Settings. Biol. Trace Elem. Res. 2019, 189, 267–276. [Google Scholar] [CrossRef]
- Zhang, W.; Kang, Z.; Wang, Q.; Qiu, N.; Zhou, F. The biological effects of strontium (88Sr) on Chinese cabbage. Plant Soil Environ. 2020, 66, 149–154. [Google Scholar] [CrossRef]
- Burger, A.; Lichtscheidl, I. Stable and radioactive cesium: A review about distribution in the environment, uptake and translocation in plants, plant reactions and plants’ potential for bioremediation. Sci. Total Environ. 2018, 618, 1459–1485. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, X.; Luo, X.; Tang, Y. Phytoextraction Ability of Amaranthus mangostanus L. from Contaminated Soils with Cs or Sr. J. Bioremediation Biodegrad. 2015, 6, 277. [Google Scholar]
- Yang, Y.; Cheng, Z.; Luo, Y.; Li, X.; Lei, Y.; Plenković-Moraj, A.; Chen, K. Preparation and characterization of Racomitrium japonicum moss biochar and its adsorption of Sr (II). BioResources 2022, 17, 3965. [Google Scholar] [CrossRef]
- Straaer, B. Measuring fast fluorescence transients to address environmental questions: The JIP-test. Photosynth. Light Biosph. 1995, 94, 977–980. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. Probing Photosynth. Mech. Regul. Adapt. 2000, 25, 445–483. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence; Springer: Berlin/Heidelberg, Germany, 2004; pp. 321–362. [Google Scholar]
- Strauss, A.; Krüger, G.; Strasser, R.; Van Heerden, P. Ranking of dark chilling tolerance in soybean genotypes probed by the chlorophyll a fluorescence transient OJIP. Environ. Exp. Bot. 2006, 56, 147–157. [Google Scholar] [CrossRef]
- Strasser, R.J. The Fo and the OJIP fluorescence rise in higher plants and algae. In Regulation of Chloroplast Biogenesis; Springer: Berlin/Heidelberg, Germany, 1992; pp. 423–426. [Google Scholar]
- Stirbet, A. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B Biol. 2011, 104, 236–257. [Google Scholar] [CrossRef]
- Zushi, K.; Kajiwara, S.; Matsuzoe, N. Chlorophyll a fluorescence OJIP transient as a tool to characterize and evaluate response to heat and chilling stress in tomato leaf and fruit. Sci. Hortic. 2012, 148, 39–46. [Google Scholar] [CrossRef]
- Tsimilli-Michael, M.; Pêcheux, M.; Strasser, R.J. Light and heat stress adaptation of the symbionts of temperate and coral reef foraminifers probed in hospite by the chlorophyll a fluorescence kinetics. Z. Naturforschung C 1999, 54, 671–680. [Google Scholar] [CrossRef]
- Stirbet, A. Chlorophyll a fluorescence induction: A personal perspective of the thermal phase, the J–I–P rise. Photosynth. Res. 2012, 113, 15–61. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-S.; Li, P.; Cheng, L. Comparison of thermotolerance of sun-exposed peel and shaded peel of ‘Fuji’ apple. Environ. Exp. Bot. 2009, 66, 110–116. [Google Scholar] [CrossRef]
- Stefanov, D.; Petkova, V.; Denev, I.D. Screening for heat tolerance in common bean (Phaseolus vulgaris L.) lines and cultivars using JIP-test. Sci. Hortic. 2011, 128, 1–6. [Google Scholar] [CrossRef]
- Lazár, D. Parameters of photosynthetic energy partitioning. J. Plant Physiol. 2015, 175, 131–147. [Google Scholar] [CrossRef]
- Chen, L.-S.; Li, P.; Cheng, L. Effects of high temperature coupled with high light on the balance between photooxidation and photoprotection in the sun-exposed peel of apple. Planta 2008, 228, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-S.; Cheng, L. Photosystem 2 is more tolerant to high temperature in apple (Malus domestica Borkh.) leaves than in fruit peel. Photosynthetica 2009, 47, 112–120. [Google Scholar] [CrossRef]
- Appenroth, K.-J.; Stöckel, J.; Srivastava, A.; Strasser, R. Multiple effects of chromate on the photosynthetic apparatus of Spirodela polyrhiza as probed by OJIP chlorophyll a fluorescence measurements. Environ. Pollut. 2001, 115, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yang, J.; Zhang, M.; Strasser, R.J.; Qiang, S. Classification and characteristics of heat tolerance in Ageratina adenophora populations using fast chlorophyll a fluorescence rise OJIP. Environ. Exp. Bot. 2016, 122, 126–140. [Google Scholar] [CrossRef]
- Guo, Y.; Lu, Y.; Goltsev, V.; Strasser, R.J.; Kalaji, H.M.; Wang, H.; Wang, X.; Chen, S.; Qiang, S. Comparative effect of tenuazonic acid, diuron, bentazone, dibromothymoquinone and methyl viologen on the kinetics of Chl a fluorescence rise OJIP and the MR820 signal. Plant Physiol. Biochem. 2020, 156, 39–48. [Google Scholar] [CrossRef]
- Oukarroum, A.; Schansker, G.; Strasser, R.J. Drought stress effects on photosystem I content and photosystem II thermotolerance analyzed using Chl a fluorescence kinetics in barley varieties differing in their drought tolerance. Physiol. Plant. 2009, 137, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, M.A.; Kumar, D.; Rajwanshi, R.; Strasser, R.J.; Tsimilli-Michael, M.; Sarin, N.B. Overexpression of γ-tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurements. Biochim. Biophys. Acta (BBA)-Bioenerg. 2010, 1797, 1428–1438. [Google Scholar] [CrossRef]
- Strasser, R.J. Energy pipeline model of the photosynthetic apparatus. In Progress in Photosynthesis Research; Springer: Dordrecht, The Netherlands, 1987; pp. 717–720. [Google Scholar]
- Strasser, R.; Eggenberg, P.; Strasser, B. How to work without stress but with fluorescence. Bull. Soc. R. Sci. Liege 1996, 65, 330–349. [Google Scholar]
- Krüger, G.H.; Tsimilli-Michael, M.; Strasser, R.J. Light stress provokes plastic and elastic modifications in structure and function of photosystem II in camellia leaves. Physiol. Plant. 1997, 101, 265–277. [Google Scholar] [CrossRef]
- Tsimilli-Michael, M.; Eggenberg, P.; Biro, B.; Köves-Pechy, K.; Vörös, I.; Strasser, R. Synergistic and antagonistic effects of arbuscular mycorrhizal fungi and Azospirillum and Rhizobium nitrogen-fixers on the photosynthetic activity of alfalfa, probed by the polyphasic chlorophyll a fluorescence transient OJIP. Appl. Soil Ecol. 2000, 15, 169–182. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, Y.; Lu, Y.; Shi, J.; Chen, S.; Strasser, R.; Qiang, S.; Hu, Z. Special issue in honour of Prof. Reto J. Strasser-Effect of AtLFNR1 deficiency on chlorophyll a fluorescence rise kinetics OJIP of Arabidopsis. Photosynthetica 2020, 58, 206–213. [Google Scholar] [CrossRef]
- Lei, Y.-B.; Xia, H.-X.; Chen, K.; Plenković-Moraj, A.; Huang, W.; Sun, G. Photosynthetic regulation in response to fluctuating light conditions under temperature stress in three mosses with different light requirements. Plant Sci. 2021, 311, 111020. [Google Scholar] [CrossRef]
- Sung, D.-Y.; Kaplan, F.; Lee, K.-J.; Guy, C.L. Acquired tolerance to temperature extremes. Trends Plant Sci. 2003, 8, 179–187. [Google Scholar] [CrossRef]
- Liu, X.; Huang, B. Photosynthetic acclimation to high temperatures associated with heat tolerance in creeping bentgrass. J. Plant Physiol. 2008, 165, 1947–1953. [Google Scholar] [CrossRef]
- Schröder, W.P.; Arellano, J.B.; Bittner, T.; Baron, M.; Eckert, H.-J.; Renger, G. Flash-induced absorption spectroscopy studies of copper interaction with photosystem II in higher plants. J. Biol. Chem. 1994, 269, 32865–32870. [Google Scholar] [CrossRef]
- Yruela, I.; Gatzen, G.; Picorel, R.; Holzwarth, A.R. Cu (II)-inhibitory effect on photosystem II from higher plants. A picosecond time-resolved fluorescence study. Biochemistry 1996, 35, 9469–9474. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Guissé, B.; Greppin, H.; Strasser, R.J. Regulation of antenna structure and electron transport in photosystem II of Pisum sativum under elevated temperature probed by the fast polyphasic chlorophyll a fluorescence transient: OKJIP. Biochim. Biophys. Acta (BBA)-Bioenerg. 1997, 1320, 95–106. [Google Scholar] [CrossRef]
- Strasser, B.J. Donor side capacity of photosystem II probed by chlorophyll a fluorescence transients. Photosynth. Res. 1997, 52, 147–155. [Google Scholar] [CrossRef]
- Küpper, H.; Benedikty, Z.; Morina, F.; Andresen, E.; Mishra, A.; Trtílek, M. Analysis of OJIP chlorophyll fluorescence kinetics and QA reoxidation kinetics by direct fast imaging. Plant Physiol. 2019, 179, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Tóth, S.Z.; Schansker, G.; Garab, G.; Strasser, R.J. Photosynthetic electron transport activity in heat-treated barley leaves: The role of internal alternative electron donors to photosystem II. Biochim. Biophys. Acta (BBA)-Bioenerg. 2007, 1767, 295–305. [Google Scholar] [CrossRef]
- Briantais, J.-M.; Dacosta, J.; Goulas, Y.; Ducruet, J.-M.; Moya, I. Heat stress induces in leaves an increase of the minimum level of chlorophyll fluorescence, Fo: A time-resolved analysis. Photosynth. Res. 1996, 48, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Havaux, M.; Greppin, H.; Strasser, R.J. Functioning of photosystems I and II in pea leaves exposed to heat stress in the presence or absence of light. Planta 1991, 186, 88–98. [Google Scholar] [CrossRef]
- Schansker, G.; Srivastava, A.; Strasser, R.J. Characterization of the 820-nm transmission signal paralleling the chlorophyll a fluorescence rise (OJIP) in pea leaves. Funct. Plant Biol. 2003, 30, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Schansker, G.; Tóth, S.Z.; Strasser, R.J. Methylviologen and dibromothymoquinone treatments of pea leaves reveal the role of photosystem I in the Chl a fluorescence rise OJIP. Biochim. Biophys. Acta (BBA)-Bioenerg. 2005, 1706, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Chen, K.; Liu, L.; Plenkovic-Moraj, A.; Sun, G.; Lei, Y. Photosynthetic regulation in fluctuating light under combined stresses of high temperature and dehydration in three contrasting mosses. Plant Sci. 2022, 323, 111379. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Huang, R.; Li, Y.; Li, W.; Zheng, L.; Lei, Y.; Chen, K. Photosynthetic regulation in response to strontium stress in moss Racomitrium japonicum L. Environ. Sci. Pollut. Res. 2022, 30, 20923–20933. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef]
- Dimitrova, S.; Paunov, M.; Pavlova, B.; Dankov, K.; Kouzmanova, M.; Velikova, V.; Tsonev, T.; Kalaji, H.; Goltsev, V. Photosynthetic efficiency of two Platanus orientalis L. ecotypes exposed to moderately high temperature–JIP-test analysis. Photosynthetica 2020, 58, 657–670. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment (Sr2+ mg/L) | Chl (mg/g) | Chl a (mg/g) | Chl b (mg/g) |
|---|---|---|---|
| 0 | 101.00 ± 1.06 a | 69.68 ± 0.96 a | 31.33 ± 0.10 a |
| 5 | 106.51 ± 0.35 a | 74.50 ± 0.23 a | 32.01 ± 0.13 a |
| 50 | 67.50 ± 2.47 b | 46.35 ± 2.12 b | 21.15 ± 0.35 b |
| 500 | 66.10 ± 3.57 b | 47.07 ± 2.97 b | 19.03 ± 0.62 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, H.; Lu, Y.; Tang, Y.; Ren, P.; Tang, H.; Chen, Q.; Kuang, P.; Huang, R.; Zhu, W.; Chen, K. Photosynthetic Responses of Racomitrium japonicum L. to Strontium Stress Evaluated through Chlorophyll a Fluorescence OJIP Transient Analysis. Plants 2024, 13, 591. https://doi.org/10.3390/plants13050591
Ren H, Lu Y, Tang Y, Ren P, Tang H, Chen Q, Kuang P, Huang R, Zhu W, Chen K. Photosynthetic Responses of Racomitrium japonicum L. to Strontium Stress Evaluated through Chlorophyll a Fluorescence OJIP Transient Analysis. Plants. 2024; 13(5):591. https://doi.org/10.3390/plants13050591
Chicago/Turabian StyleRen, Hui, Yunmei Lu, Yunlai Tang, Peng Ren, Hao Tang, Qunlong Chen, Peigang Kuang, Renhua Huang, Wenkun Zhu, and Ke Chen. 2024. "Photosynthetic Responses of Racomitrium japonicum L. to Strontium Stress Evaluated through Chlorophyll a Fluorescence OJIP Transient Analysis" Plants 13, no. 5: 591. https://doi.org/10.3390/plants13050591