
The Laccase Family Gene CsLAC37 Participates in Resistance to Colletotrichum gloeosporioides Infection in Tea Plants

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
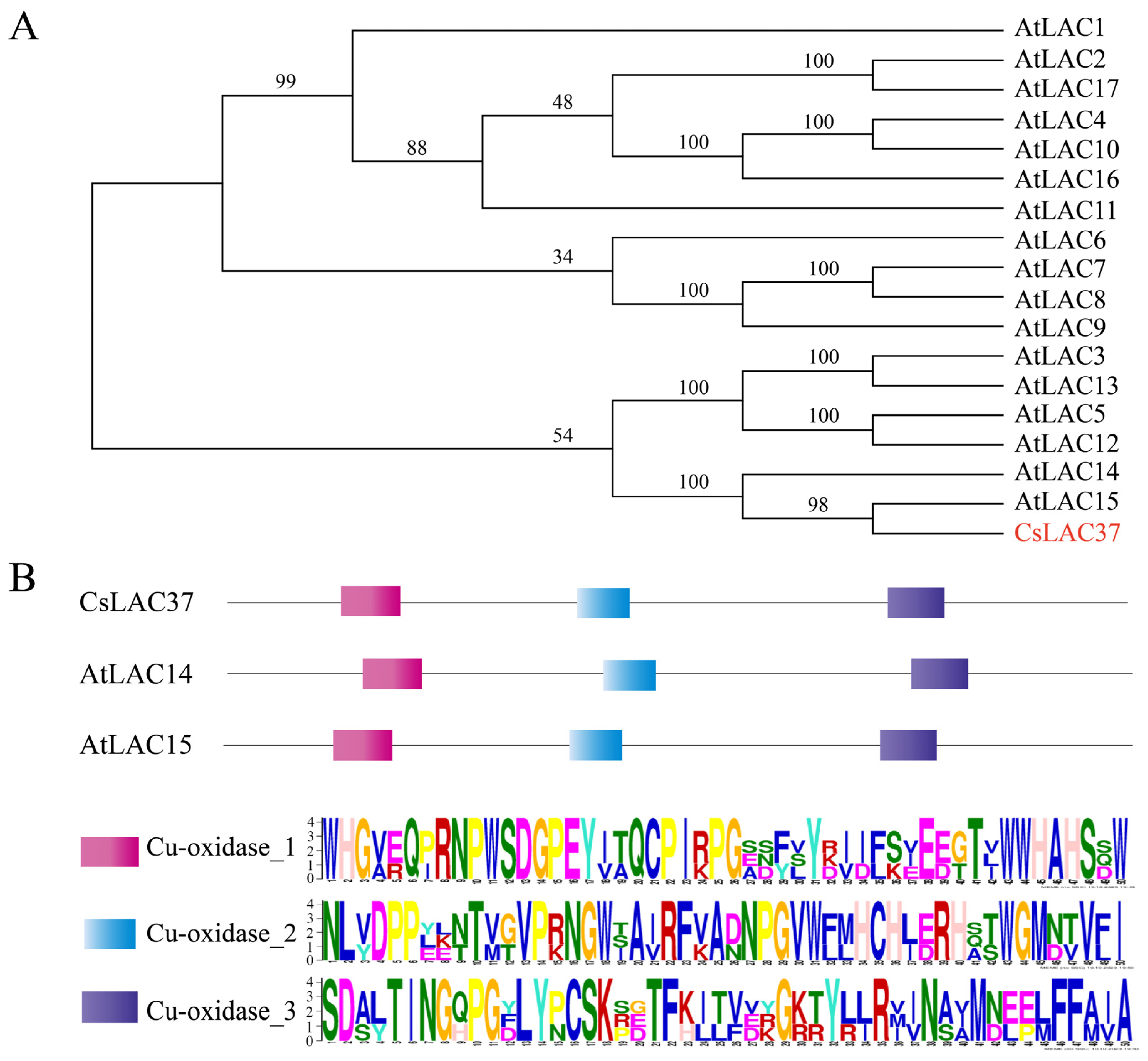
2.1. Identification of CsLAC37 and Protein Sequence Analysis
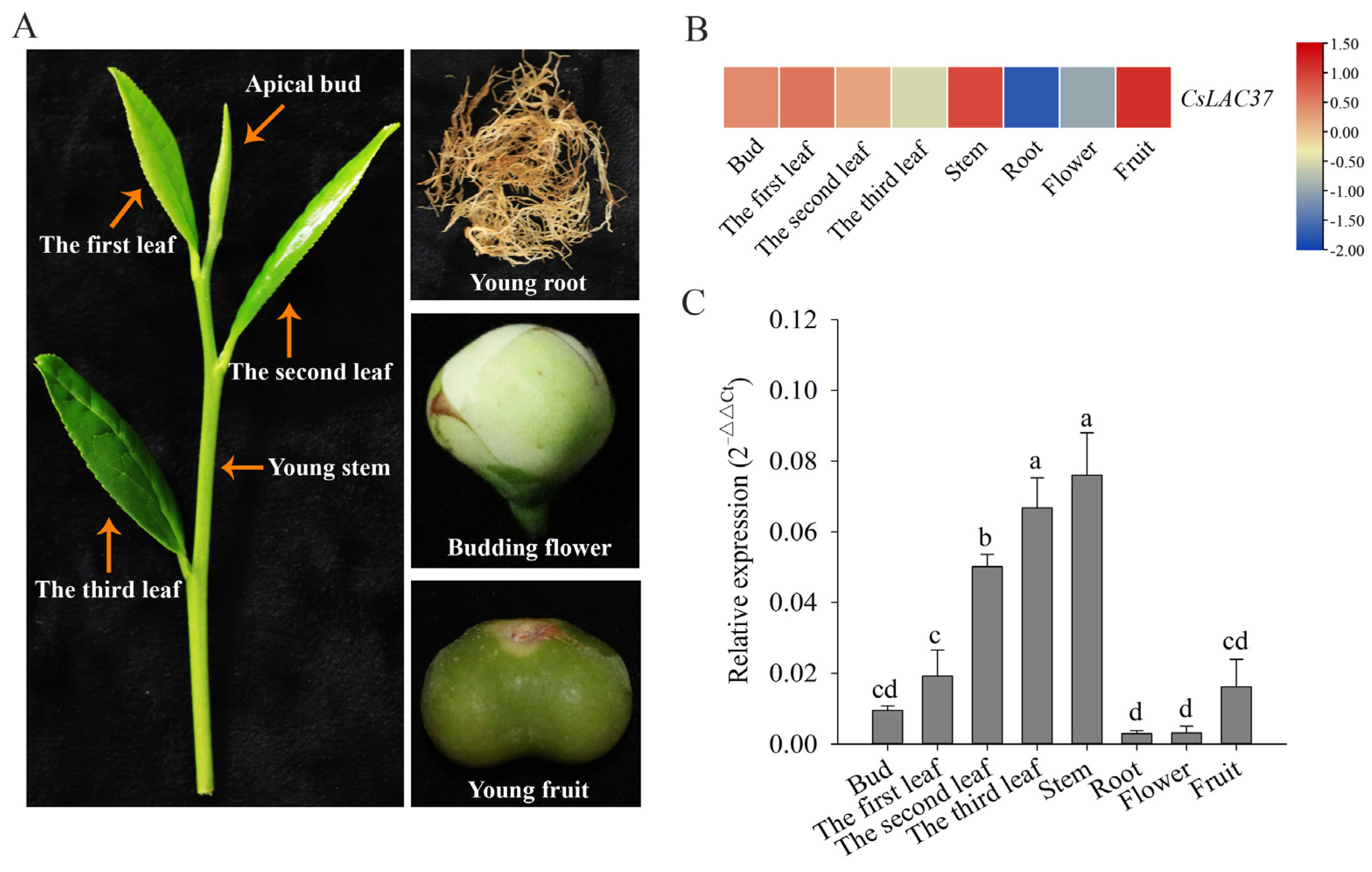
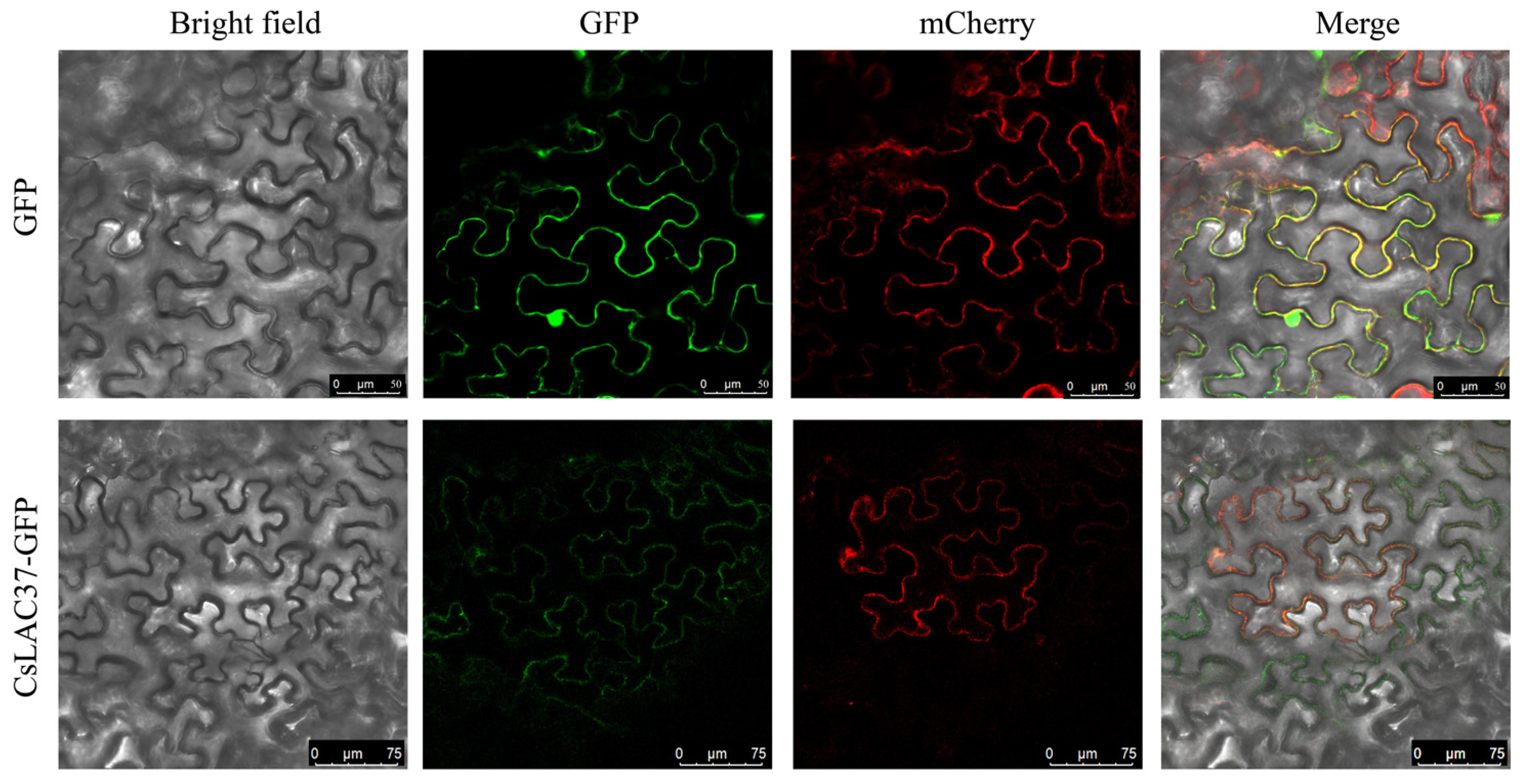
2.2. Tissue-Specific Expression Patterns and Subcellular Localization
2.3. Expression Analysis in Response to Fungal Infection
2.4. Expression Analysis in Response to Cold and Salt Stresses
2.5. Expression Analysis in Response to SA and ABA Treatments
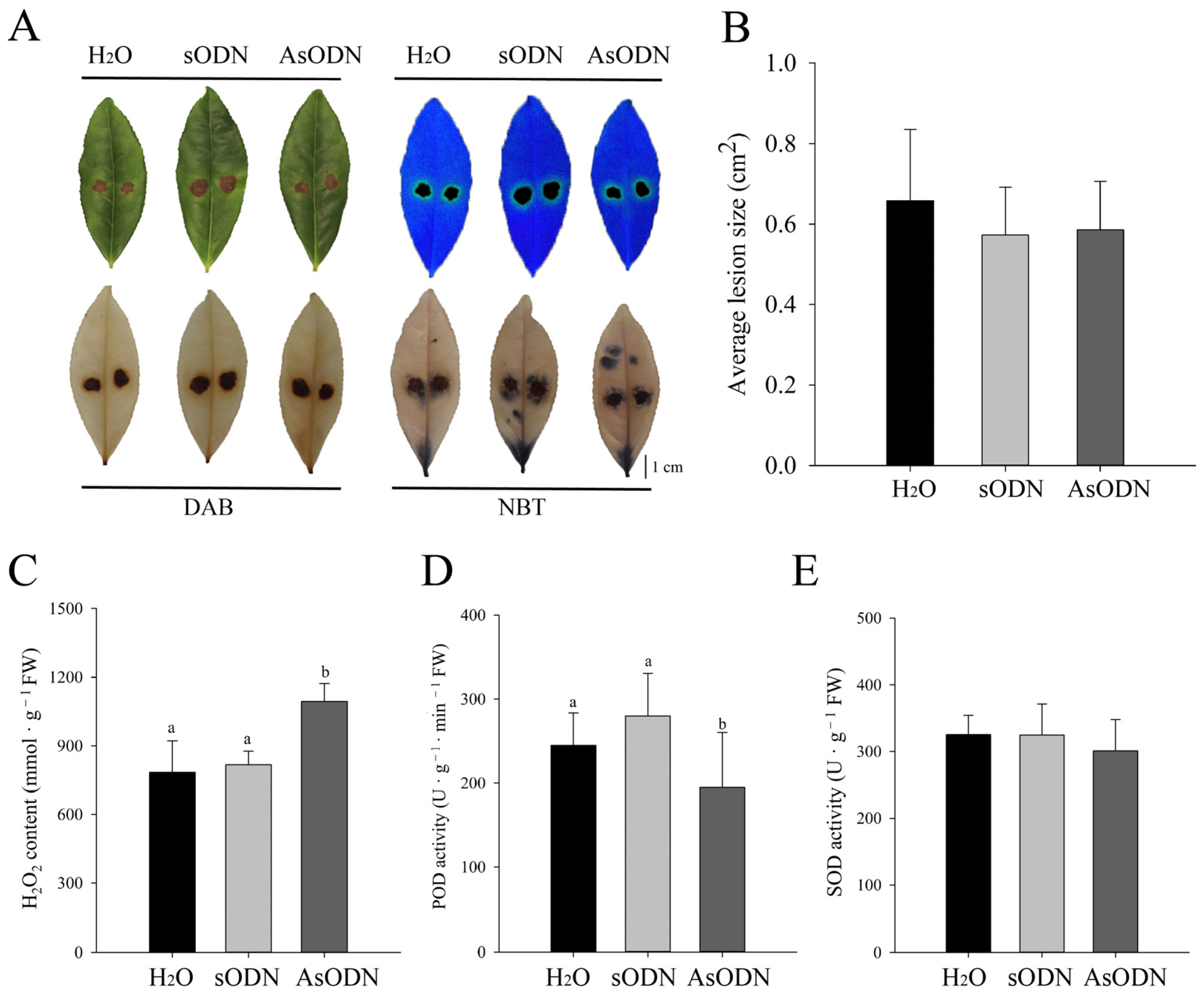
2.6. Functional Validation of CsLAC37 in Resistance to Fungal Infection
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Molecular Cloning and Bioinformatic Analysis
4.3. Subcellular Localization
4.4. RNA Extraction and qRT-PCR Analysis
4.5. Gene Suppression Using AsODNs
4.6. Pathogen Inoculation and Treatment
4.7. DAB and NBT Staining Assay
4.8. Enzyme Activity Detection of POD and SOD
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Q.; Zhu, J.; Ren, N.; Li, D.; Jin, Y.; Lu, W.; Lu, Q. Characteristics and Pathogenicity of Discula theae-sinensis Isolated from Tea Plant (Camellia sinensis) and Interaction with Colletotrichum spp. Plants 2023, 12, 3427. [Google Scholar] [CrossRef]
- Wang, W.; Xie, X.; Lv, Y.; Guan, H.; Liu, L.; Huang, Q.; Bao, Y.; Zhou, J.; Bao, L.; Gong, C.; et al. Identification and profile of phenolamides with anthracnose resistance potential in tea (Camellia sinensis). Hortic. Res. 2023, 10, uhad154. [Google Scholar] [CrossRef]
- Fang, W.P.; Yang, L.C.; Zhu, X.J.; Zeng, L.; Li, X. Seasonal and habitat dependent variations in culturable endophytes of Camellia sinensis. J. Plant Pathol. Microbiol. 2013, 4, 169. [Google Scholar] [CrossRef]
- Jeyaraj, A.; Wang, X.; Wang, S.; Liu, S.; Zhang, R.; Wu, A.; Wei, C. Identification of Regulatory Networks of MicroRNAs and Their Targets in Response to Colletotrichum gloeosporioides in Tea Plant (Camellia sinensis L.). Front. Plant Sci. 2019, 10, 1096. [Google Scholar] [CrossRef]
- Chen, X.; Chen, X.; Tan, Q.; Mo, X.; Liu, J.; Zhou, G. Recent progress on harm, pathogen classification, control and pathogenic molecular mechanism of anthracnose of oil-tea. Front. Microbiol. 2022, 13, 918339. [Google Scholar] [CrossRef]
- Wu, Y.; Tan, Y.; Peng, Q.; Xiao, Y.; Xie, J.; Li, Z.; Ding, H.; Pan, H.; Wei, L. Biocontrol potential of endophytic bacterium Bacillus altitudinis GS-16 against tea anthracnose caused by Colletotrichum gloeosporioides. PeerJ 2024, 12, e16761. [Google Scholar] [CrossRef]
- Holtappels, D.; Fortuna, K.; Lavigne, R.; Wagemans, J. The future of phage biocontrol in integrated plant protection for sustainable crop production. Curr. Opin. Biotech. 2021, 68, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhang, H.; Huang, K.; Guo, R.; Zhao, J.; Xie, H.; Zhu, J.; Gu, H.; Chen, H.; Li, G.; et al. Comprehensive analysis of the laccase gene family in tea plant highlights its roles in development and stress responses. BMC Plant Biol. 2023, 23, 129. [Google Scholar] [CrossRef]
- Khan, W.; El-Shehawi, A.M.; Ali, F.; Ali, M.; Alqurashi, M.; Althaqafi, M.M.; Alharthi, S.B. A Genome-Wide Identification and Expression Pattern of LMCO Gene Family from Turnip (Brassica rapa L.) under Various Abiotic Stresses. Plants 2023, 12, 1904. [Google Scholar] [CrossRef]
- Yu, Y.; Xing, Y.; Liu, F.; Zhang, X.; Li, X.; Zhang, J.; Sun, X. The Laccase Gene Family Mediate Multi-Perspective Trade-Offs during Tea Plant (Camellia sinensis) Development and Defense Processes. Int. J. Mol. Sci. 2021, 22, 12554. [Google Scholar] [CrossRef]
- Bai, Y.; Ali, S.; Liu, S.; Zhou, J.; Tang, Y. Characterization of plant laccase genes and their functions. Gene 2023, 852, 147060. [Google Scholar] [CrossRef]
- Berthet, S.; Demont-Caulet, N.; Pollet, B.; Bidzinski, P.; Cezard, L.; Le Bris, P.; Borrega, N.; Herve, J.; Blondet, E.; Balzergue, S.; et al. Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification of Arabidopsis thaliana stems. Plant Cell 2011, 23, 1124–1137. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Nakashima, J.; Chen, F.; Yin, Y.; Fu, C.; Yun, J.; Shao, H.; Wang, X.; Wang, Z.Y.; Dixon, R.A. Laccase is necessary and nonredundant with peroxidase for lignin polymerization during vascular development in Arabidopsis. Plant Cell 2013, 25, 3976–3987. [Google Scholar] [CrossRef]
- Wang, Y.; Bouchabke-Coussa, O.; Lebris, P.; Antelme, S.; Soulhat, C.; Gineau, E.; Dalmais, M.; Bendahmane, A.; Morin, H.; Mouille, G.; et al. LACCASE5 is required for lignification of the Brachypodium distachyon Culm. Plant Physiol. 2015, 168, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhuo, C.; Xiao, X.; Wang, X.; Docampo-Palacios, M.; Chen, F.; Dixon, R.A. Substrate Specificity of LACCASE8 Facilitates Polymerization of Caffeyl Alcohol for C-Lignin Biosynthesis in the Seed Coat of Cleome hassleriana. Plant Cell 2020, 32, 3825–3845. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, L.; Wang, X.; Chen, B.; Zhao, J.; Cui, J.; Li, Z.; Yang, J.; Wu, L.; Wu, J.; et al. The cotton laccase gene GhLAC15 enhances Verticillium wilt resistance via an increase in defence-induced lignification and lignin components in the cell walls of plants. Mol. Plant Pathol. 2019, 20, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Long, Y.; Yin, X.; Wang, W.; Li, W.; Jiang, L.; Chen, X.; Wang, B.; Ma, J. Genome-wide identification and expression patterns of the laccase gene family in response to kiwifruit bacterial canker infection. BMC Plant Biol. 2023, 23, 591. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, L.; Mi, X.; Zhao, S.; An, Y.; Xia, X.; Guo, R.; Wei, C. Multi-omics analysis to visualize the dynamic roles of defense genes in the response of tea plants to gray blight. Plant J. 2021, 106, 862–875. [Google Scholar] [CrossRef]
- Tai, Y.; Liu, C.; Yu, S.; Yang, H.; Sun, J.; Guo, C.; Huang, B.; Liu, Z.; Yuan, Y.; Xia, E.; et al. Gene co-expression network analysis reveals coordinated regulation of three characteristic secondary biosynthetic pathways in tea plant (Camellia sinensis). BMC Plant Biol. 2018, 19, 616. [Google Scholar] [CrossRef]
- Wan, S.; Wang, W.; Zhou, T.; Zhang, Y.; Chen, J.; Xiao, B.; Yang, Y.; Yu, Y. Transcriptomic analysis reveals the molecular mechanisms of Camellia sinensis in response to salt stress. Plant Growth Regul. 2018, 84, 481–492. [Google Scholar] [CrossRef]
- McCaig, B.; Meagher, R.; Dean, J. Gene structure and molecular analysis of the laccase-like multicopper oxidase (LMCO) gene family in Arabidopsis thaliana. Planta 2005, 221, 619–636. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Li, G.; Ma, C.; Abdullah, M.; Zhang, J.; Zhao, H.; Jin, Q.; Cai, Y.; Lin, Y. Comprehensive genome-wide analysis of thepear (Pyrus bretschneideri) laccase gene (PbLAC) family and functional identification of PbLAC1 involved in lignin biosynthesis. PLoS ONE 2019, 14, e0210892. [Google Scholar]
- Li, L.; Yang, K.; Wang, S.; Lou, Y.; Zhu, C.; Gao, Z. Genome-wide analysis of laccase genes in moso bamboo highlights PeLAC10 involved in lignin biosynthesis and in response to abiotic stresses. Plant Cell Rep. 2020, 39, 751–763. [Google Scholar] [CrossRef]
- Zhuang, Y.; Zuo, D.; Tao, Y.; Cai, H.; Li, L. Laccase3-based extracellular domain provides possible positional information for directing Casparian strip formation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2020, 117, 15400–15402. [Google Scholar] [CrossRef] [PubMed]
- Yi Chou, E.; Schuetz, M.; Hoffmann, N.; Watanabe, Y.; Sibout, R.; Samuels, A. Distribution, Mobility and Anchoring of Lignin-Related Oxidative Enzymes in Arabidopsis Secondary Cell Walls. J. Exp. Bot. 2018, 69, 1849–1859. [Google Scholar] [CrossRef]
- He, F.; Machemer-Noonan, k.; Golfier, P.; Unda, F.; Dechert, J.; Zhang, W.; Hoffmann, N.; Samuels, L.; Mansfield, S.; Rausch, T.; et al. The in vivo impact of MsLAC1, a Miscanthus laccase isoform, on lignification and lignin composition contrasts with its in vitro substrate preference. BMC Plant Biol. 2019, 19, 552. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.M.; Kerhoas, L.; Caboche, M.; Lepiniec, L.; Debeaujon, I. TRANSPARENT TESTA10 encodes a laccase-like enzyme involved in oxidative polymerization of flavonoids in Arabidopsis seed coat. Plant Cell 2005, 17, 2966–2980. [Google Scholar] [CrossRef]
- Liang, M.; Davis, E.; Gardner, D.; Cai, X.; Wu, Y. Involvement of AtLAC15 in lignin synthesis in seeds and in root elongation of Arabidopsis. Planta 2006, 224, 1185–1196. [Google Scholar] [CrossRef]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic Acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef]
- He, J.; Liu, Y.; Yuan, D.; Duan, M.; Liu, Y.; Shen, Z.; Yang, C.; Qiu, Z.; Liu, D.; Wen, P.; et al. An R2R3 MYB transcription factor confers brown planthopper resistance by regulating the phenylalanine ammonia-lyase pathway in rice. Proc. Natl. Acad. Sci. USA 2020, 117, 271–277. [Google Scholar] [CrossRef]
- Gallego-Giraldoa, L.; Escamilla-Trevinob, L.; Jacksona, L.; Dixon, R. Salicylic acid mediates the reduced growth of lignin down-regulated plants. Proc. Natl. Acad. Sci. USA 2011, 108, 20814–20819. [Google Scholar] [CrossRef]
- Zhang, Z.; Long, Y.; Yin, X.; Yang, S. Sulfur-Induced Resistance against Pseudomonas syringae pv. actinidiae via Triggering Salicylic Acid Signaling Pathway in Kiwifruit. Int. J. Mol. Sci. 2021, 22, 12710. [Google Scholar] [PubMed]
- Mittler, R.; Zandalinas, S.; Fichman, Y.; Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Dong, S.; Li, M.; Gu, F.; Yang, G.; Guo, T.; Chen, Z.; Wang, J. The Class III peroxidase gene OsPrx30, transcriptionally modulated by the AT-hook protein OsATH1, mediates rice bacterial blight-induced ROS accumulation. J. Integr. Plant Biol. 2021, 63, 393–408. [Google Scholar] [CrossRef]
- Li, Y.; Ma, X.; Xiao, L.D.; Yu, Y.N.; Yan, H.L.; Gong, Z.H. CaWRKY50 Acts as a Negative Regulator in Response to Colletotrichum scovillei Infection in Pepper. Plants 2023, 12, 1962. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Guo, L.; Zhou, Q.; Jiang, Z.; Jin, L.; Zhu, J.; Xie, H.; Wei, C. Identification and Functional Analysis of Two Alcohol Dehydrogenase Genes Involved in Catalyzing the Reduction of (Z)-3-Hexenal into (Z)-3-Hexenol in Tea Plants (Camellia sinensis). J. Agric. Food Chem. 2022, 70, 1830–1839. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wang, X.; Guo, L.; Xu, Q.; Zhao, S.; Li, F.; Yan, X.; Liu, S.; Wei, C. Characterization and Alternative Splicing Profiles of the Lipoxygenase Gene Family in Tea Plant (Camellia sinensis). Plant Cell Physiol. 2018, 59, 1765–1781. [Google Scholar] [CrossRef]
- Liu, S.; Rao, J.; Zhu, J.; Li, G.; Li, F.; Zhang, H.; Tao, L.; Zhou, Q.; Tao, Y.; Zhang, Y.; et al. Integrated physiological, metabolite and proteomic analysis reveal the glyphosate stress response mechanism in tea plant (Camellia sinensis). J. Hazard. Mater. 2023, 454, 131419. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using realtime quantitative PCR and the 2−ΔΔCt method. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef]
- Dinc, E.; Toth, S.Z.; Schansker, G.; Ayaydin, F.; Kovacs, L.; Dudits, D.; Garab, G.; Bottka, S. Synthetic antisense oligodeoxynucleotides to transiently suppress different nucleus- and chloroplast-encoded proteins of higher plant chloroplasts. Plant Physiol. 2011, 157, 1628–1641. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Zhang, H.; Zhou, Q.; Tao, Y.; Wang, S.; Wang, P.; Wang, A.; Wei, C.; Liu, S. The Laccase Family Gene CsLAC37 Participates in Resistance to Colletotrichum gloeosporioides Infection in Tea Plants. Plants 2024, 13, 884. https://doi.org/10.3390/plants13060884
Li D, Zhang H, Zhou Q, Tao Y, Wang S, Wang P, Wang A, Wei C, Liu S. The Laccase Family Gene CsLAC37 Participates in Resistance to Colletotrichum gloeosporioides Infection in Tea Plants. Plants. 2024; 13(6):884. https://doi.org/10.3390/plants13060884
Chicago/Turabian StyleLi, Dangqiang, Hongxiu Zhang, Qianqian Zhou, Yongning Tao, Shuangshuang Wang, Pengke Wang, Aoni Wang, Chaoling Wei, and Shengrui Liu. 2024. "The Laccase Family Gene CsLAC37 Participates in Resistance to Colletotrichum gloeosporioides Infection in Tea Plants" Plants 13, no. 6: 884. https://doi.org/10.3390/plants13060884
APA StyleLi, D., Zhang, H., Zhou, Q., Tao, Y., Wang, S., Wang, P., Wang, A., Wei, C., & Liu, S. (2024). The Laccase Family Gene CsLAC37 Participates in Resistance to Colletotrichum gloeosporioides Infection in Tea Plants. Plants, 13(6), 884. https://doi.org/10.3390/plants13060884