
Transcriptomic and Metabolomic Research on the Germination Process of Panax ginseng Overwintering Buds

 ,
,
Abstract
:1. Introduction
2. Results
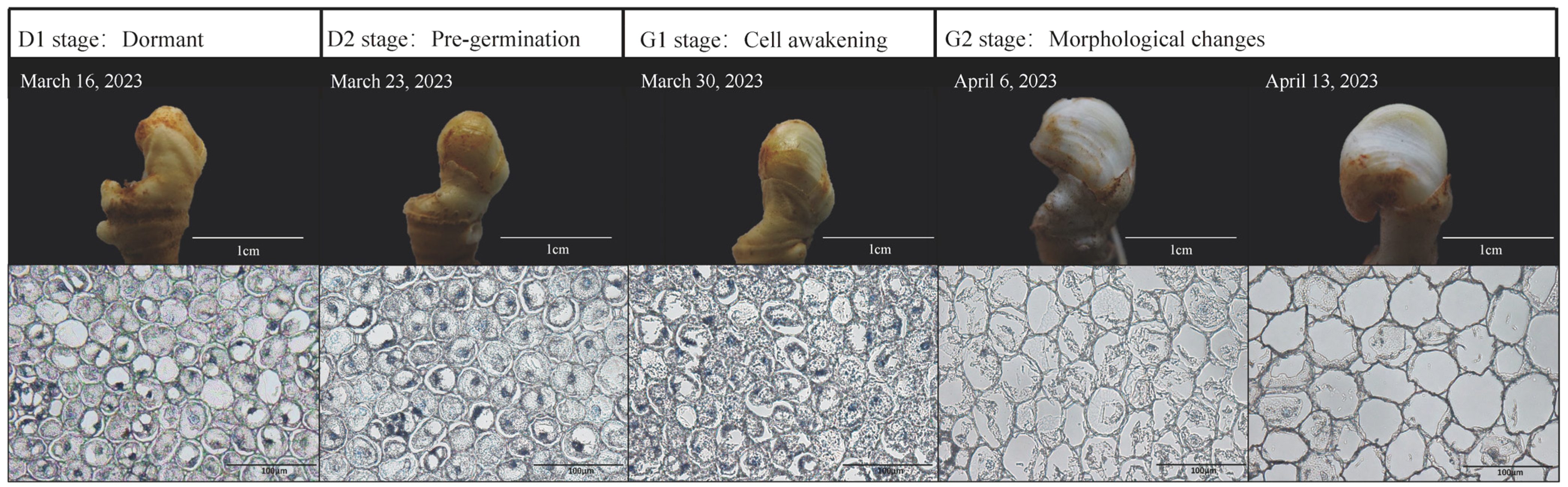
2.1. Observation on the Appearance of Overwintering Buds and Paraffin Sections of Young Stems
2.2. Results of Metabolomic Analysis
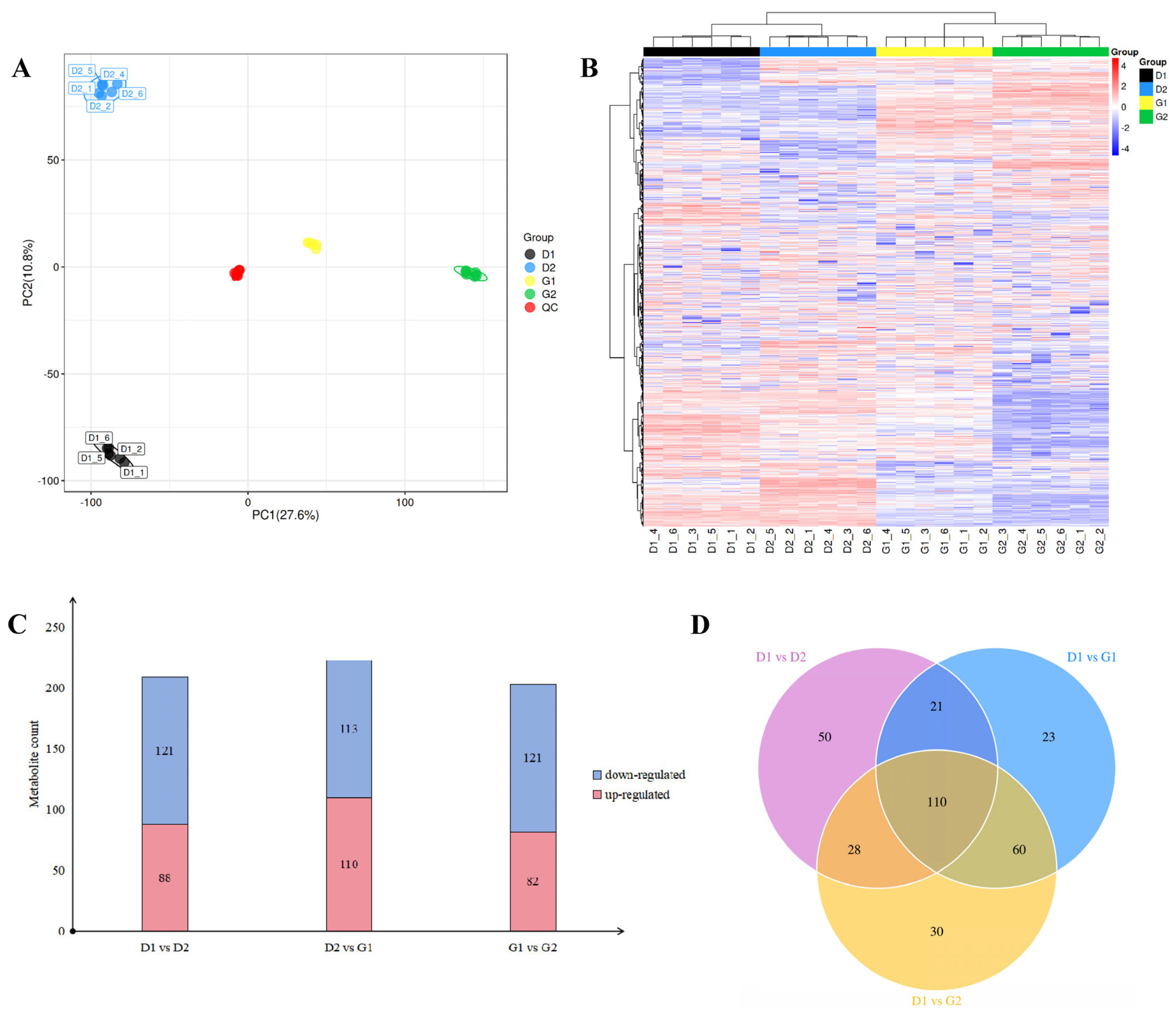
2.2.1. Quality Evaluation of Metabolome Data
2.2.2. Screening of Differentially Accumulated Metabolites (DAMs) at Different Stages during the Dormancy Release Process of Ginseng Overwintering Buds
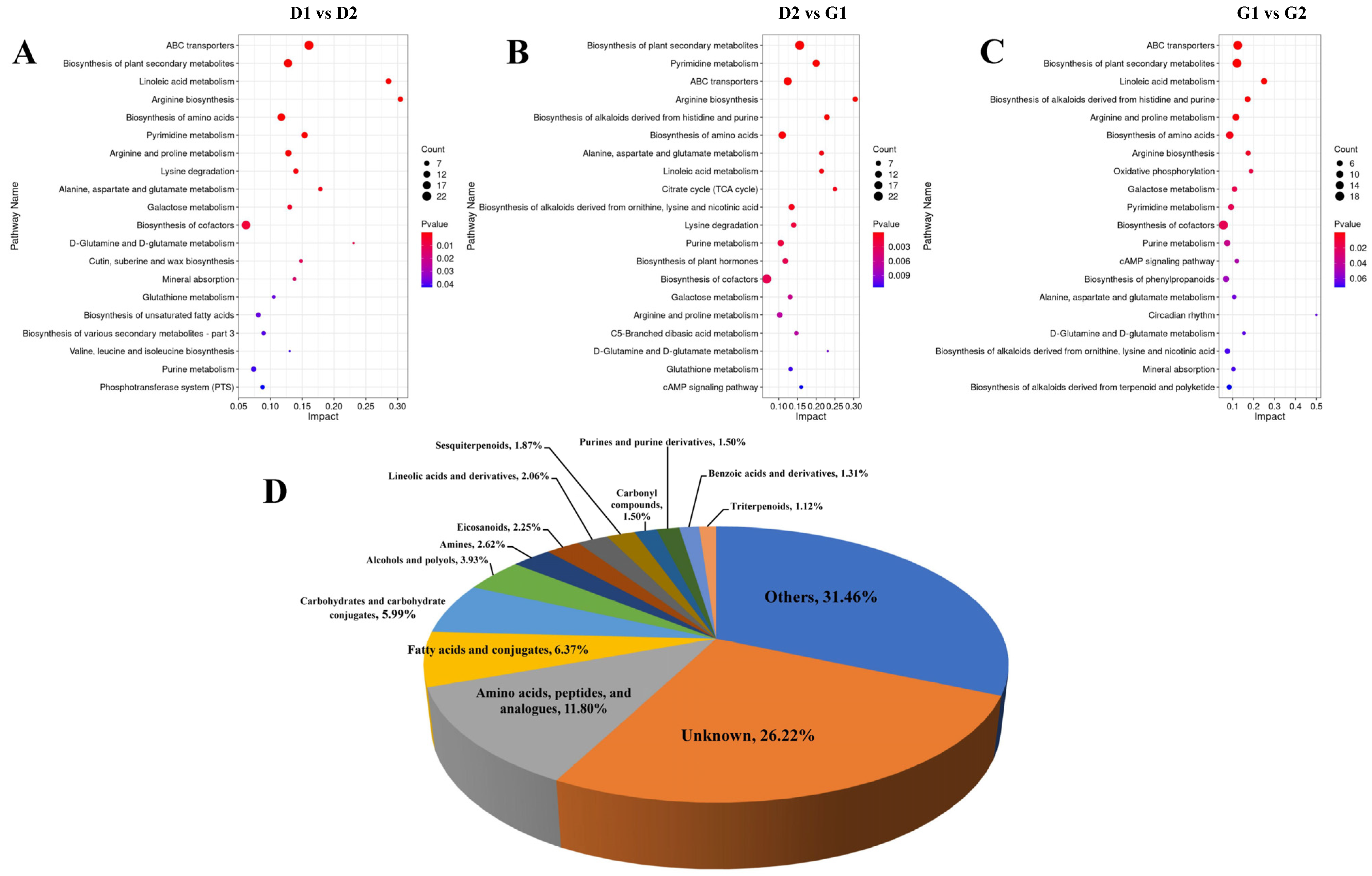
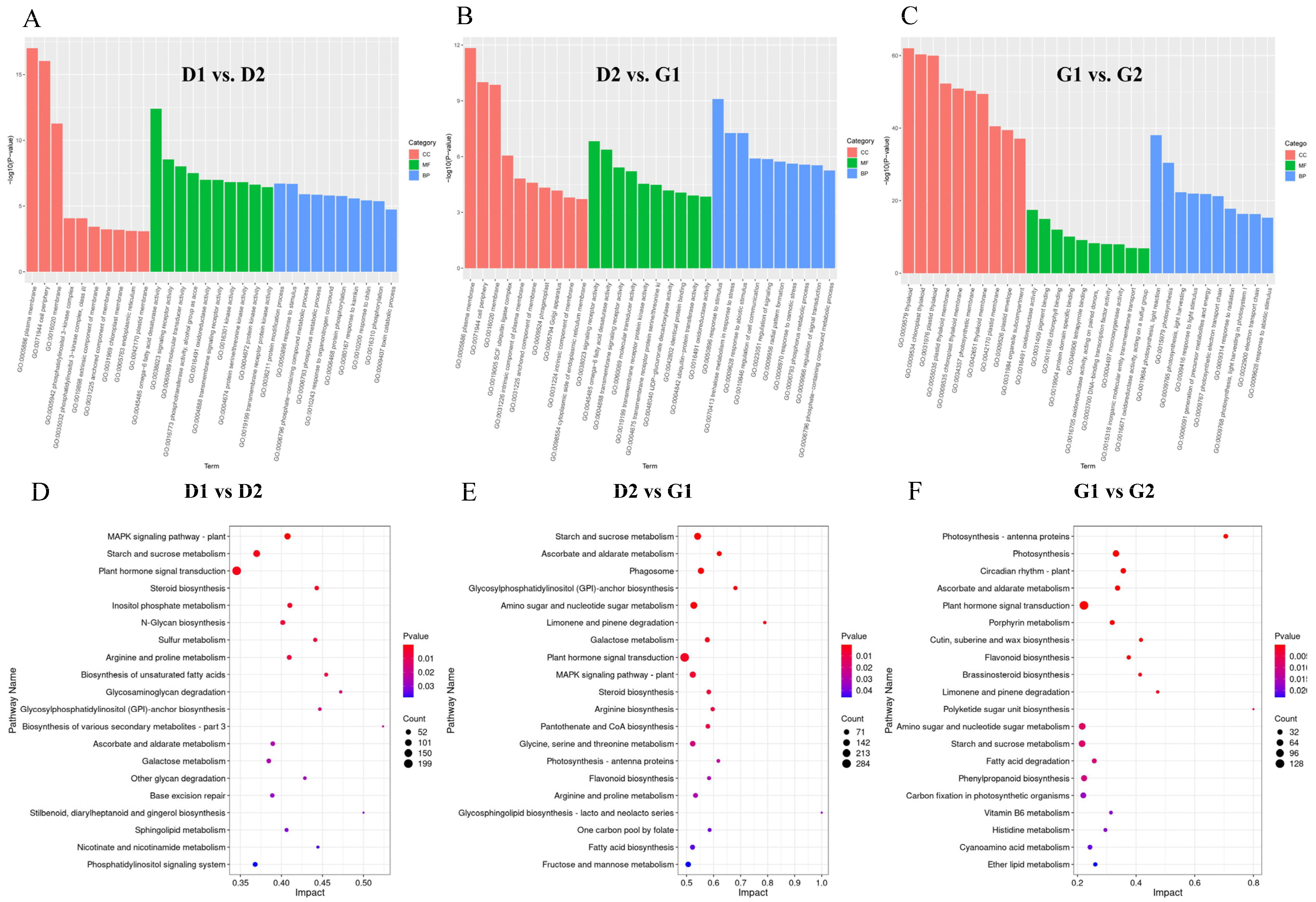
2.2.3. KEGG Pathway Analysis of DAM during Ginseng Overwintering Bud Germination
2.3. Result of Transcriptomic Analysis
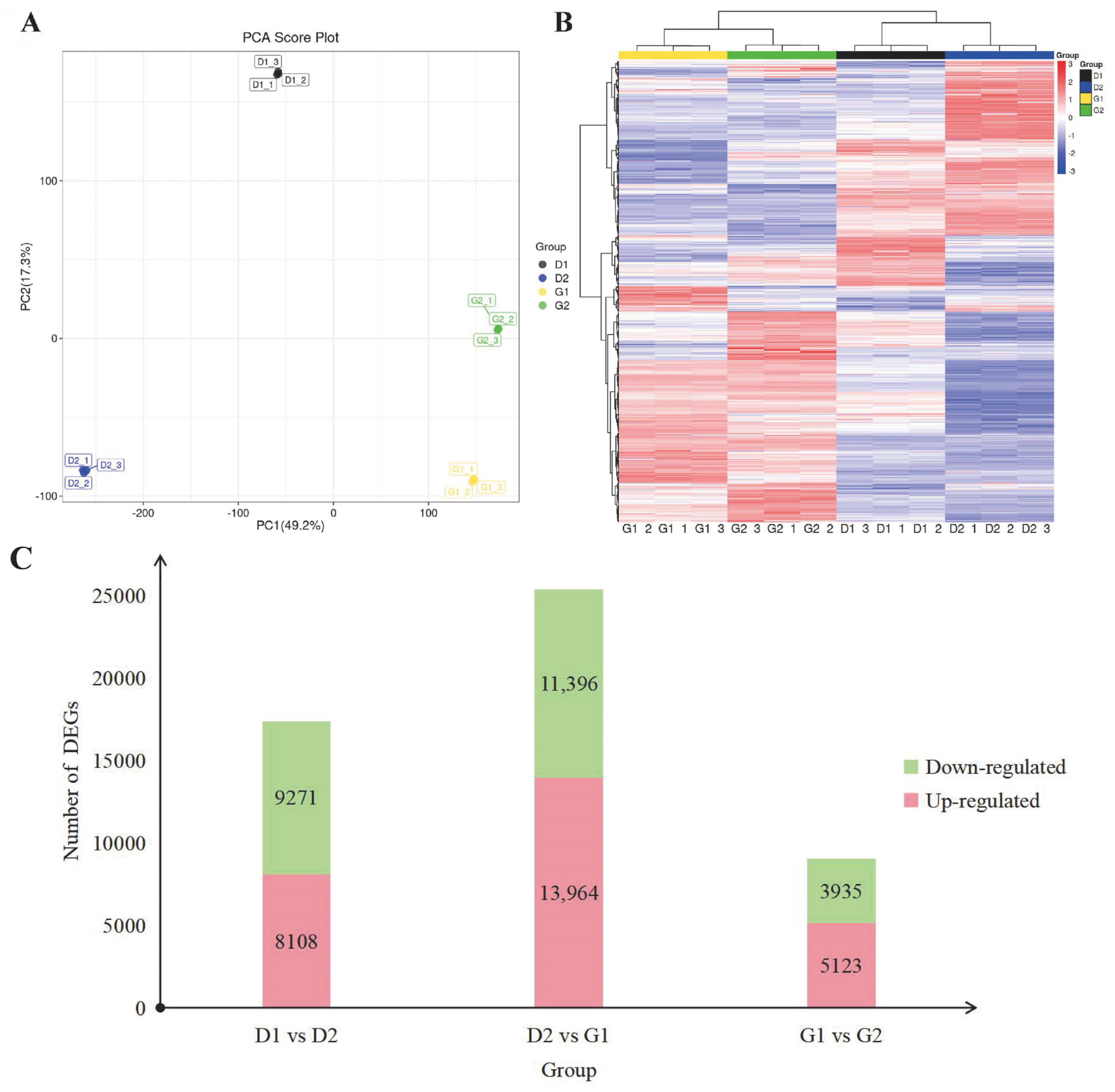
2.3.1. Overview of Transcriptome Data
2.3.2. Enrichment Analysis of DEGs
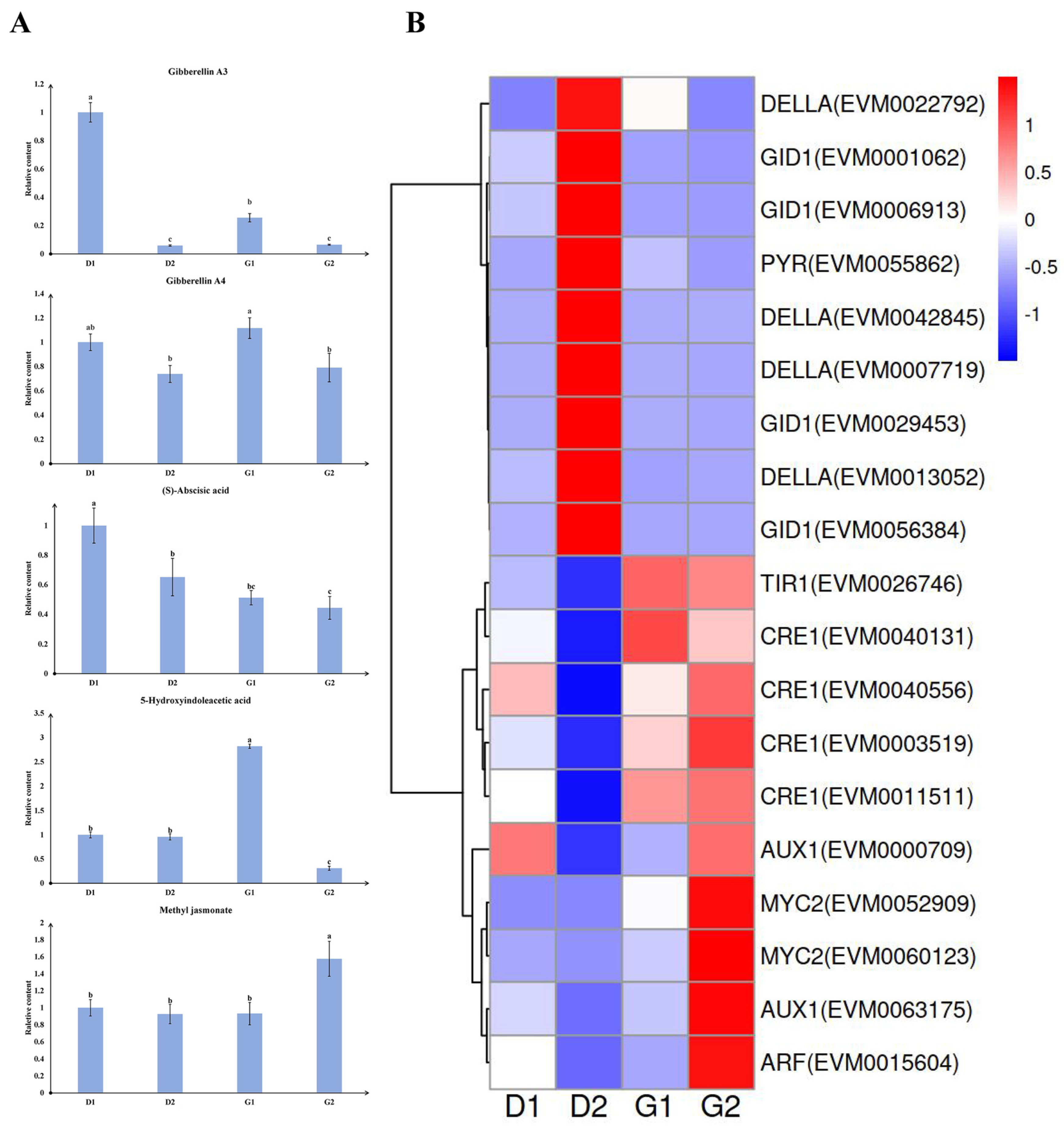
2.4. Plant Hormones and Related Genes during Germination
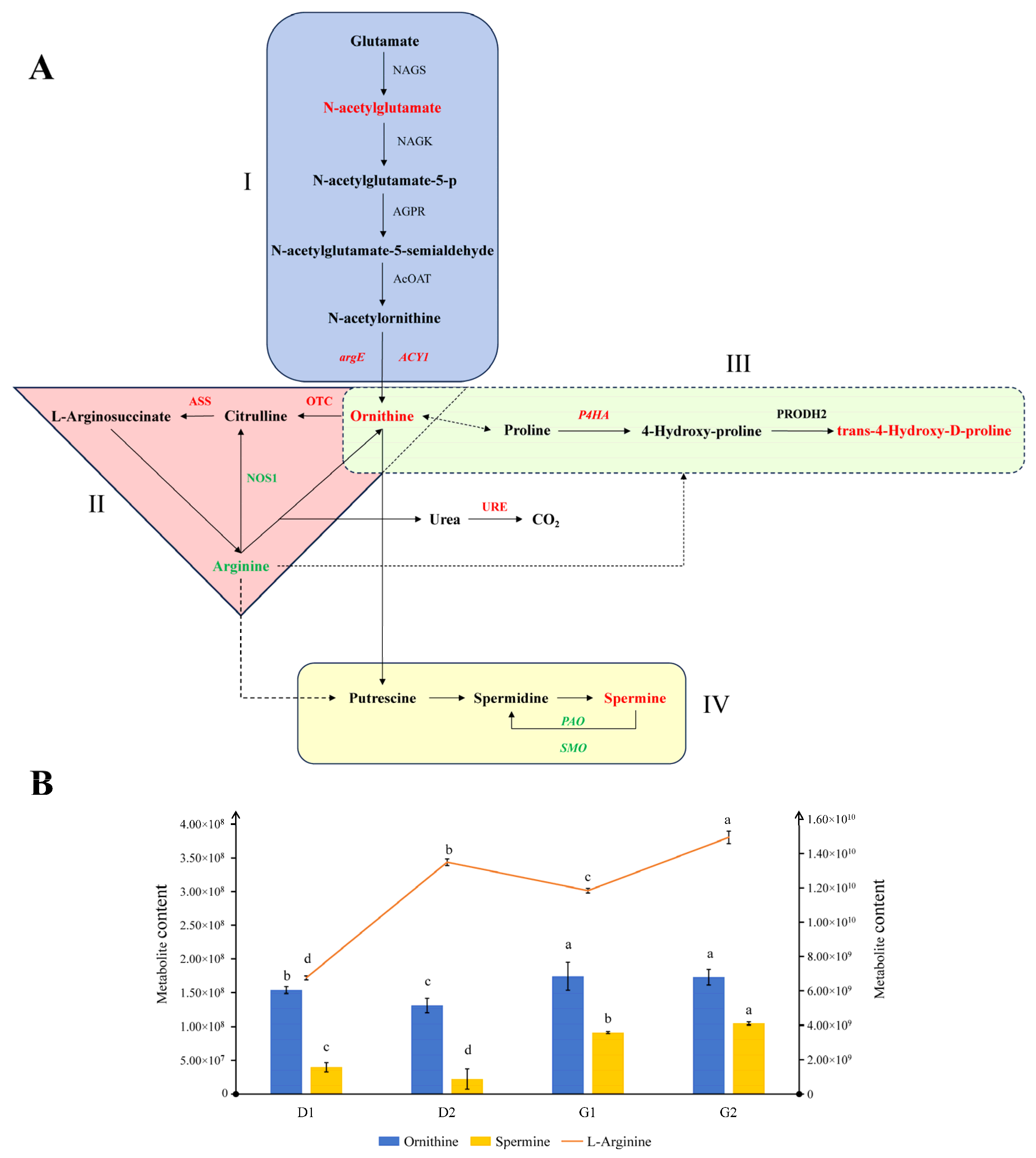
2.5. Arginine-Related Pathways during Ginseng Overwintering Bud Germination
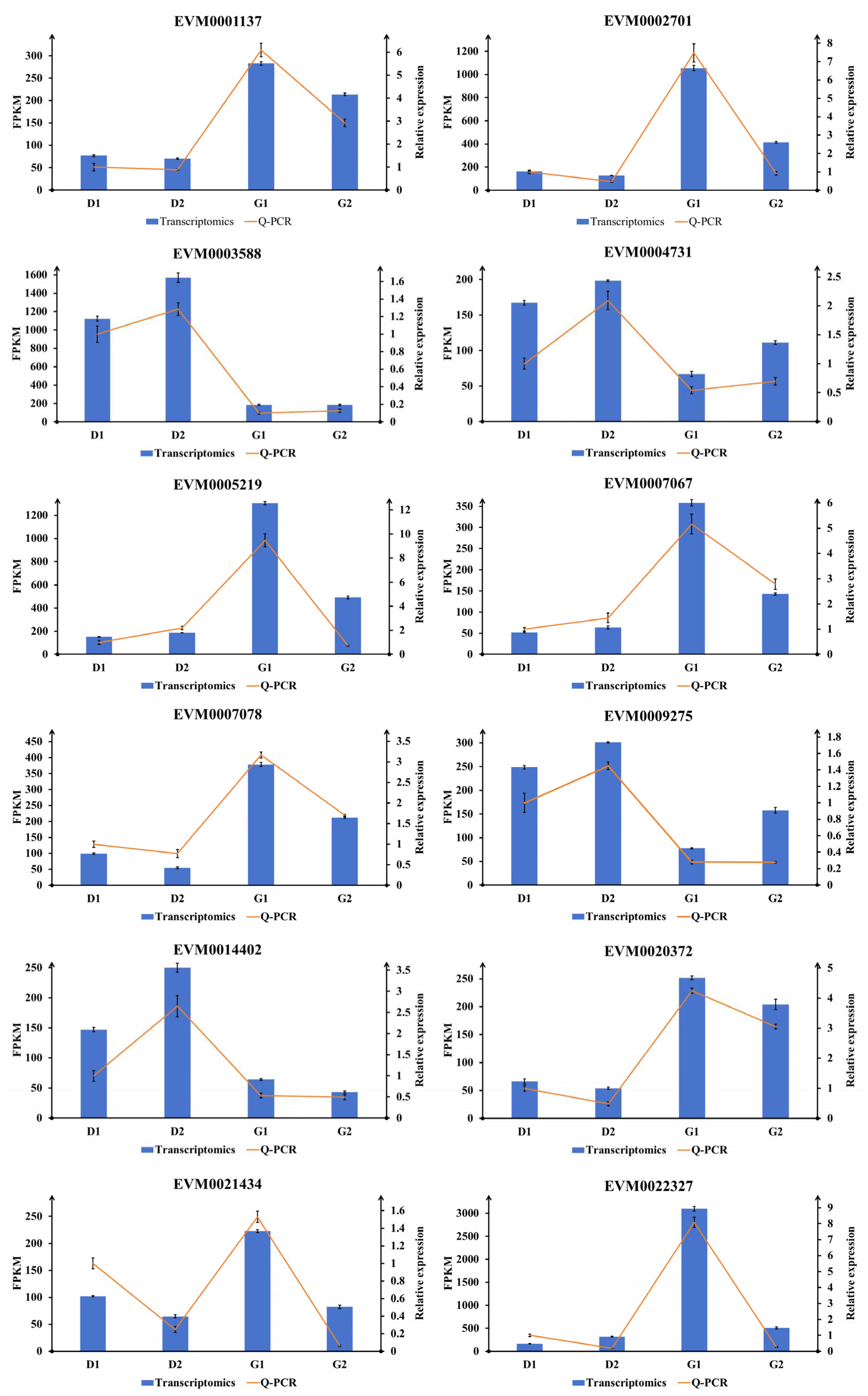
2.6. Validation of Transcriptomic Results Using Q-PCR
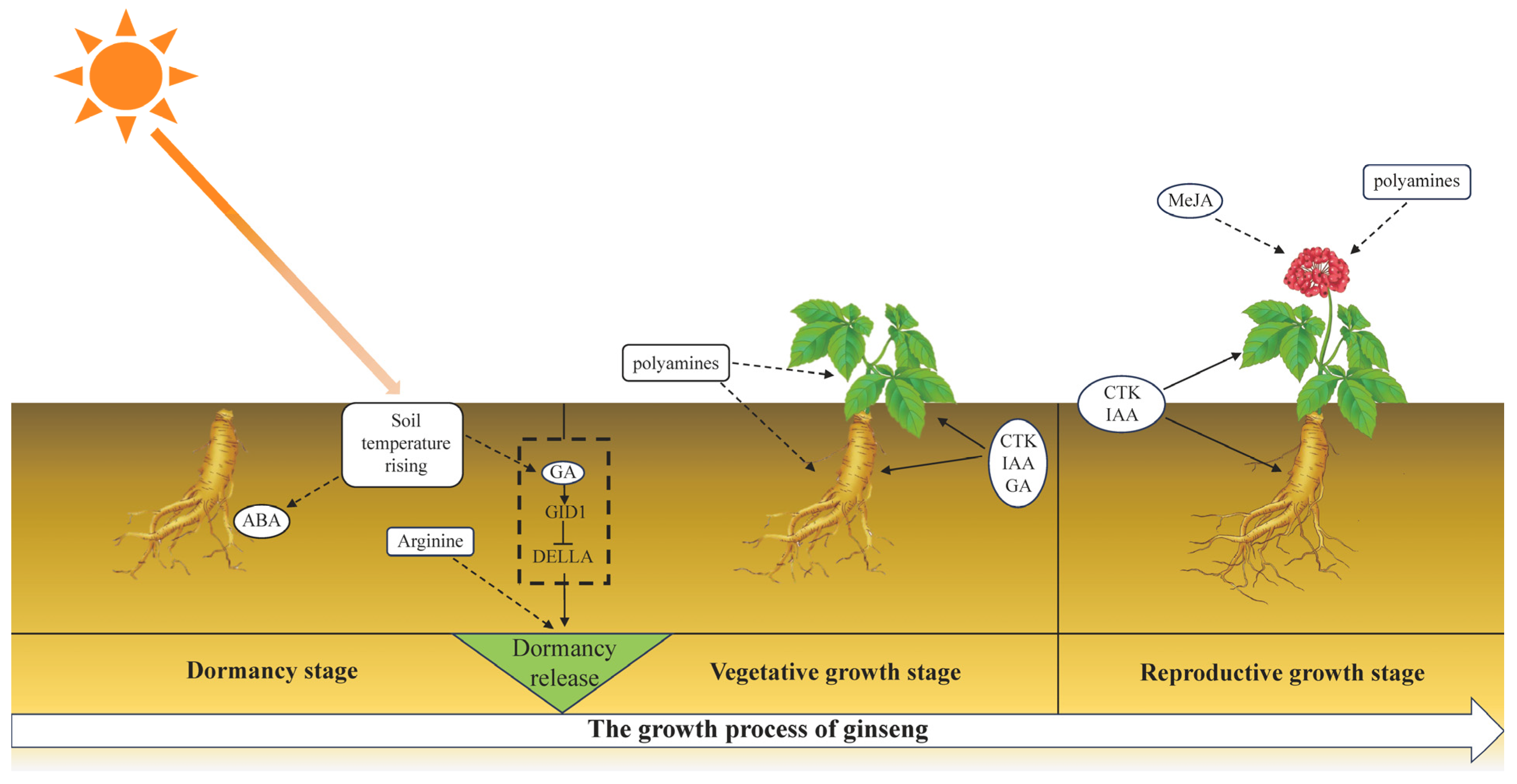
3. Discussion
3.1. The Role of Phytohormones in the Germination of Ginseng Overwintering Buds
3.2. Arginine and Polyamine Metabolism during the Germination of Ginseng Overwintering Buds
4. Materials and Methods
4.1. Plant Material and Treatment
4.2. Paraffin Section
4.3. Non-Targeted Metabolomics Analysis
4.3.1. Samples Preparation
4.3.2. Liquid Chromatography Conditions
4.3.3. Mass Spectrum Conditions
4.3.4. Data Analysis
4.4. RNA Sequencing
4.4.1. RNA Extraction and Detection
4.4.2. Library Construction and Quality Inspection
4.4.3. Data Analysis
4.5. Q-PCR Verification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endo-, para-, and ecodormancy: Physiological terminology and classification for dormancy research. Hortscience 1987, 22, 371–377. [Google Scholar] [CrossRef]
- Rohde, A.; Bhalerao, R.P. Plant dormancy in the perennial context. Trends Plant Sci. 2007, 12, 217–223. [Google Scholar] [CrossRef]
- Kim, J.H.; Yi, Y.-S.; Kim, M.-Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef]
- Ji, Y.; Ai, J.; Wang, Y.; Wang, Z. A Study on Morphogenesis of Flower Bud Differentiation in Panax ginseng C. A. Mey. Spec. Wild Econ. Anim. Plant Res. 2009, 31, 40–43. [Google Scholar]
- Yang, L.; Song, M.; Zhang, J. Effect of light and temperature on seed germination of 6 species of Lilium L. J. Plant Resour. Environ. 2000, 9, 14–18. [Google Scholar]
- Fan, W.N.; Sun, X.G.; Ni, J.X.; Du, H.Q.; Shi, Y.H.; Yan, X.B.; Wang, C.Z. Effect of photoperiod on phytochromes and endogenous hormones of alfalfa with different fall-dormancies. Acta Prataculturae Sin. 2014, 23, 177–184. [Google Scholar]
- Feng, Y.; Zhu, L.; Pan, T.; Guo, Z.; Zhong, X.; Ding, A.; Pan, D. Characterization of summer dormancy in Narcissus tazetta var. Chinensis and the role of NtFTs in summer dormancy and flower differentiation. Sci. Hortic-Amst. 2015, 183, 109–117. [Google Scholar] [CrossRef]
- Delvallée, I.; Paffen, A.; De Klerk, G.J. The development of dormancy in bulblets of Lilium speciosum generated in vitro. II. The effect of temperature. Physiol. Plant. 1990, 80, 431–436. [Google Scholar] [CrossRef]
- Langens-Gerrits, M.M.; Miller, W.B.; Croes, A.F.; De Klerk, G.J. Effect of low temperature on dormancy breaking and growth after planting in lily bulblets regenerated in vitro. Plant Growth Regul. 2003, 40, 267–275. [Google Scholar] [CrossRef]
- Petrić, M.; Jevremović, S.; Trifunović, M.; Tadić, V.; Milošević, S.; Dragićević, M.; Subotić, A. The effect of low temperature and GA3 treatments on dormancy breaking and activity of antioxidant enzymes in Fritillaria meleagris bulblets cultured in vitro. Acta Physiol. Plant 2013, 35, 3223–3236. [Google Scholar] [CrossRef]
- Wu, L.Q.; Li, A.X. Effects of different low temperature storage on bulb growth and development of lily. For. Sci. Technol. 2022, 9, 43–46. [Google Scholar]
- Sun, Y.; Liu, P.; Zhao, Y.; Liu, C.; Su, C. Extraction, isolation and identification of germination inhibitory substances from corm of Amorphophallus konjac K. Koch. Physiol. Mol. Biol. Plants 1996, 22, 277–283. [Google Scholar]
- Ge, H.; Li, Q.; Chen, G.; Zhang, G.; Li, S. Changes in endogenous plant hormones during dormancy of strawberry. Acta Hortic. Sin. 1998, 25, 90–91. [Google Scholar]
- Jiang, N.N.; Zhang, Q.X.; Wang, Y.; Sun, Y.; Fang, Y.F.; Xu, J.G. Effects of GA3 on dormancy release, endogenous hormones levels and sugar metabolism in Paeonia lactiflora ‘Da Fugui’. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2020, 44, 26–32. [Google Scholar]
- Shi, Z.W.; Hao, J.J.; Zhen, H.X.; Zhao, H.M.; Niu, Y.B. Establishment of tissue culture system of Fritillaria cirrhosa and the influence of ABA and GA3 on bulb germination. Mol. Plant Breed. 2022, 20, 3347–3354. [Google Scholar]
- Carter, C.E.; Partis, M.D.; Thomas, B. The Expression of Histone 2A in Onion (Allium cepa) during the Onset of Dormancy, Storage and Emergence from Dormancy. New Phytol. 1999, 143, 461–470. [Google Scholar] [CrossRef]
- Freeman, D.; Riou-Khamlichi, C.; Oakenfull, E.A.; Murray, J.A. Isolation, characterization and expression of cyclin and cyclin-dependent kinase genes in Jerusalem artichoke (Helianthus tuberosus L.). J. Exp. Bot. 2003, 54, 303–308. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Gao, S.Y. Brief report on the experiment of breaking the dormancy of Panax ginseng in winter. J. Northeast Agric. Sci. 1964, 2, 60. [Google Scholar]
- Chen, Z.; Hao, L.; Qin, G. Effects of gibberellin and low temperature on breaking dormancy of Panax ginseng and Panax quinquefolium. J. Chin. Med. Mater. 1986, 4, 1–4. [Google Scholar] [CrossRef]
- John, C.S.; Proctor, T.A.; Bernadine, J.C.L.; Li, G. Effect of temperature on dormancy and growth of Panax quinquefolium. For. By-Prod. Spec. China 1987, 2, 32–34. [Google Scholar]
- Kim, D.H.; Kim, Y.C.; Bang, K.H.; Kim, J.U.; Lee, J.W.; Cho, I.H.; Kim, Y.B.; Son, S.W.; Park, J.B.; Kim, K.H. Effects of GA3 and Alternating Temperature on Breaking Bud Dormancy of Panax ginseng C. A. Meyer Seedling. Korean J. Med. Crop Sci. 2015, 23, 379–384. [Google Scholar] [CrossRef]
- Tanino, K.K. Hormones and Endodormancy Induction in Woody Plants. J. Crop Improv. 2004, 10, 157–199. [Google Scholar] [CrossRef]
- Nagar, P.K. Changes in endogenous abscisic acid and phenols during winter dormancy in tea (Camellia sinensis L. (O) Kuntze ). Acta Physiol. Plant 1996, 18, 33–38. [Google Scholar]
- Or, E.; Belausov, E.; Popilevsky, I.; Bental, Y. Changes in endogenous ABA level in relation to the dormancy cycle in grapevines grown in a hot climate. J. Hortic. Sci. Biotechnol. 2000, 75, 190–194. [Google Scholar] [CrossRef]
- Zhang, T.; Yuan, Y.; Zhan, Y.; Cao, X.; Liu, C.; Zhang, Y.; Gai, S. Metabolomics analysis reveals Embden Meyerhof Parnas pathway activation and flavonoids accumulation during dormancy transition in tree peony. Bmc Plant Biol. 2020, 20, 484. [Google Scholar] [CrossRef]
- Tuan, P.A.; Kumar, R.; Rehal, P.K.; Toora, P.K.; Ayele, B.T. Molecular Mechanisms Underlying Abscisic Acid/Gibberellin Balance in the Control of Seed Dormancy and Germination in Cereals. Front. Plant Sci. 2018, 9, 362906. [Google Scholar] [CrossRef]
- Arana, M.V.; Marín-de la Rosa, N.; Maloof, J.N.; Blázquez, M.A.; Alabadí, D. Circadian oscillation of gibberellin signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 9292–9297. [Google Scholar] [CrossRef]
- Ionescu, I.A.; López-Ortega, G.; Burow, M.; Bayo-Canha, A.; Junge, A.; Gericke, O.; Møller, B.L.; Sánchez-Pérez, R. Transcriptome and Metabolite Changes during Hydrogen Cyanamide-Induced Floral Bud Break in Sweet Cherry. Front. Plant Sci. 2017, 8, 270556. [Google Scholar] [CrossRef]
- Lea, P.J.; Azevedo, R.A. Nitrogen use efficiency. 2. Amino acid metabolism. Ann. Appl. Biol. 2007, 151, 269–275. [Google Scholar] [CrossRef]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Nesi, A.N.; Araújo, W.L.; Braun, H.P. Amino Acid Catabolism in Plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, F.M.; Avila, C.; Canton, F.R.; Canas, R.A.; de la Torre, F. Ammonium assimilation and amino acid metabolism in conifers. J. Exp. Bot. 2007, 58, 2307–2318. [Google Scholar] [CrossRef] [PubMed]
- Durzan, D.J. Nitrogen metabolism of Picea glauca. I. Seasonal changes of free amino acids in buds, shoot apices, and leaves, and the metabolism of uniformly labelled C-L-arginine by buds during the onset of dormancy. Can. J. Bot. 1968, 46, 909–919. [Google Scholar] [CrossRef]
- Chen, H.; McCaig, B.C.; Melotto, M.; He, S.Y.; Howe, G.A. Regulation of Plant Arginase by Wounding, Jasmonate, and the Phytotoxin Coronatine. J. Biol. Chem. 2004, 279, 45998–46007. [Google Scholar] [CrossRef]
- Ruamrungsri, S.; Ohtake, N.; Kuni, S.; Suwanthada, C.; Apavatjrut, P.; Ohyama, T. Changes in nitrogenous compounds, carbohydrates and abscisic acid in Curcuma alismatifolia Gagnep. during dormancy. J. Hortic. Sci. Biotechnol. 2001, 76, 48–51. [Google Scholar] [CrossRef]
- Sun, H.M.; Li, T.L.; Li, Y.F. Changes in free amino acid content and composition in lily bulb. Plant Physiol. J. 2004, 40, 414–418. [Google Scholar]
- Heby, O.; Marton, L.J.; Wilson, C.B.; Martinez, H.M. Polyamine metabolism in a rat brain tumor cell line: Its relationship to the growth rate. J. Cell. Physiol. 1975, 86, 511–521. [Google Scholar] [CrossRef]
- Mamont, P.S.; Boehlen, P.; McCann, P.P.; Bey, P.; Schuber, F.; Tardif, C. Alpha-methyl ornithine, a potent competitive inhibitor of ornithine decarboxylase, blocks proliferation of rat hepatoma cells in culture. Proc. Natl. Acad. Sci. USA 1976, 73, 1626–1630. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhong, B.; Li, W.; Deng, J.; Zhang, S.; Tang, Q. Variation of polyamines on shoot and flower bud of ‘Hosui’ Pear (Pyrus pyrifolia Nakai) during reflorescence period. Southwest China J. Agric. Sci. 2016, 29, 2221–2224. [Google Scholar]
- Kushad, M.M.; Orvos, A.R.; Yelenosky, G. Relative changes in polyamines during citrus flower development. Hortscience 1990, 25, 946–948. [Google Scholar] [CrossRef]
- Kaur-Sawhney, R.; Tiburcio, A.F.; Galston, A.W. Spermidine and flower-bud differentiation in thin-layer explants of tobacco. Planta 1988, 173, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Rey, M.; Díaz-Sala, C.; Rodríguez, R. Comparison of endogenous polyamine content in hazel leaves and buds between the annual dormancy and flowering phases of growth. Physiol. Plant. 1994, 91, 45–50. [Google Scholar] [CrossRef]
- Alvarez, M.E.; Savouré, A.; Szabados, L. Proline metabolism as regulatory hub. Trends Plant Sci. 2022, 27, 39–55. [Google Scholar] [CrossRef]
- Tian, W.N.; Braunstein, L.D.; Pang, J.; Stuhlmeier, K.M.; Xi, Q.C.; Tian, X.; Stanton, R.C. Importance of glucose-6-phosphate dehydrogenase activity for cell growth. J. Biol. Chem. 1998, 273, 10609–10617. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent Proportion and Code Name | A | B | C | D1 | D2 |
|---|---|---|---|---|---|
| Distilled water:70% ethanol:TBA | 3:5:2 | 2:2:4 | 0:2.5:7.5 | TBA | TBA |
| Dehydration time | 1–2 h | 1–2 h | 1–2 h | 1 h | 1 h |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, R.; Li, Y.; Tang, M.; Qu, Z.; Shao, C.; Zheng, P.; Hou, W. Transcriptomic and Metabolomic Research on the Germination Process of Panax ginseng Overwintering Buds. Plants 2024, 13, 1041. https://doi.org/10.3390/plants13071041
Li R, Li Y, Tang M, Qu Z, Shao C, Zheng P, Hou W. Transcriptomic and Metabolomic Research on the Germination Process of Panax ginseng Overwintering Buds. Plants. 2024; 13(7):1041. https://doi.org/10.3390/plants13071041
Chicago/Turabian StyleLi, Ranqi, Yashu Li, Miaomiao Tang, Zhengyi Qu, Cai Shao, Peihe Zheng, and Wei Hou. 2024. "Transcriptomic and Metabolomic Research on the Germination Process of Panax ginseng Overwintering Buds" Plants 13, no. 7: 1041. https://doi.org/10.3390/plants13071041
APA StyleLi, R., Li, Y., Tang, M., Qu, Z., Shao, C., Zheng, P., & Hou, W. (2024). Transcriptomic and Metabolomic Research on the Germination Process of Panax ginseng Overwintering Buds. Plants, 13(7), 1041. https://doi.org/10.3390/plants13071041