
A Review with a Focus on Vaccinium-Berries-Derived Bioactive Compounds for the Treatment of Reproductive Cancers


Abstract
:1. Introduction
2. Methods
3. Vaccinium Berries as Sources of Bioactive Compounds
4. The Therapeutic Role of Blueberry Bioactives in Reproductive Cancers
5. The Therapeutic Role of Cranberry Bioactives in Reproductive Cancers
5.1. Experimental Studies
5.2. Clinical Trials
6. The Therapeutic Role of Bilberry and Lingonberry Bioactives in Reproductive Cancers
7. Limitations
8. Conclusions and Future Directions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| ACNs | Anthocyanins |
| Akt | Threonine kinase |
| Anthos | Anthocyanidin aglycone |
| AR | Androgen receptor |
| ARID1A | AT-rich interaction domain 1A |
| Bax | Bcl-2-associated X protein |
| BC | Bladder cancer |
| CC | Cervical cancer |
| CDK | Cyclin-dependent kinase |
| COX-2 | Cyclooxygenase-2 |
| DNA-PK | DNA-dependent protein kinase |
| DPn | Polymerization |
| EGFR | Epidermal growth factor receptor |
| EMMPRIN | Extracellular matrix metalloproteinase inducer |
| ERK | Extracellular-signal regulated kinase |
| ExoAnthos | Exosomal Anthos |
| FGFR3 | Fibroblast growth factor receptor 3 |
| FOXA1 | Forkhead box A1 |
| HIF1α | Hypoxia inducible factor-1α |
| Iso | Isorhamnetin |
| KDM6 | Lysine (K)-specific demethylase 6 |
| Km | Kaempferol |
| KRAS | Kirsten rat sarcoma viral oncogene homologue |
| MAP | Mitogen-activated protein |
| MMPs | Matrix metalloproteinases |
| Myr | Myricetin |
| NFkB | Nuclear factor kappa-B |
| OC | Ovarian cancer |
| p21 | Protein 21 |
| p27 | Protein 27 |
| p38 MAPK | Protein 38 mitogen-activated protein kinases |
| p53 | Protein 53 |
| PACs | Proanthocyanidins |
| PAM50 | 50-gene signature |
| Par-4 | Prostate apoptosis response-4 gene |
| PARP | Poly(ADP-ribose) polymerase |
| PCa | Prostate cancer |
| PCNA | Proliferating cell nuclear antigen |
| PCS | Prostate cancer 14-pathway |
| PgP | p-glycoproteins |
| PI3K | Phosphoinositide 3-kinase |
| PIK3CA | P110alpha catalytic subunit of PI3K |
| PKA | Protein kinase A |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| PR | Progesterone receptor |
| PTEN | Phosphatase and tensin homolog |
| Qu | Quercetin |
| RAF | Retinoblastoma tumour suppressor protein-proto-oncogene |
| RB1 | Retinoblastoma gene 1 |
| RCs | Reproductive cancers |
| RCTs | Randomized controlled trials |
| ROS | Intracellular reactive oxygen species |
| SCCC | Small-cell carcinoma of the cervix |
| STAT3 | Signal transducer and activator of transcription3 |
| tBid | Truncated Bid |
| TCGA | Cancer genome atlas |
| TIMPs | Metalloproteinases |
| TNF-α | Tumour necrosis factor |
| TP53 | p53 tumour-suppressor gene |
| TRAIL | Tumour necrosis factor-related apoptosis-induced ligand |
| TUNEL+ | TUNEL-positive tumour cells |
| UTIs | Urinary tract infections |
| VEGF | Vascular endothelial growth factor |
| WT1 | Wilms’ tumour gene 1 |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Wang, L.; Lu, B.; He, M.; Wang, Y.; Wang, Z.; Du, L. Prostate cancer incidence and mortality: Global status and temporal trends in 89 countries from 2000 to 2019. Front. Public Health 2022, 10, 811044. [Google Scholar] [CrossRef] [PubMed]
- Withrow, D.; Pilleron, S.; Nikita, N.; Ferlay, J.; Sharma, S.; Nicholson, B.; Rebbeck, T.R.; Lu-Yao, G. Current and projected number of years of life lost due to prostate cancer: A global study. Prostate 2022, 82, 1088–1097. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Rumgay, H.; Li, M.; Yu, H.; Pan, H.; Ni, J. The global landscape of bladder cancer incidence and mortality in 2020 and projections to 2040. J. Glob Health 2023, 13, 04109. [Google Scholar] [CrossRef]
- Bergengren, O.; Pekala, K.R.; Matsoukas, K.; Fainberg, J.; Mungovan, S.F.; Bratt, O.; Bray, F.; Brawley, O.; Luckenbaugh, A.N.; Mucci, L.; et al. 2022 Update on prostate cancer epidemiology and risk factors-A systematic review. Eur. Urol. 2023, 84, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Jubber, I.; Ong, S.; Bukavina, L.; Black, P.C.; Compérat, E.; Kamat, A.M.; Kiemeney, L.; Lawrentschuk, N.; Lerner, S.P.; Meeks, J.J.; et al. Epidemiology of bladder cancer in 2023: A systematic review of risk factors. Eur. Urol. 2023, 84, 176–190. [Google Scholar] [CrossRef]
- Weiderpass, E.; Labrèche, F. Malignant tumors of the female reproductive system. Saf. Health Work 2012, 3, 166–180. [Google Scholar] [CrossRef]
- Cabasag, C.J.; Fagan, P.J.; Ferlay, J.; Vignat, J.; Laversanne, M.; Liu, L.; van der Aa, M.A.; Bray, F.; Soerjomataram, I. Ovarian cancer today and tomorrow: A global assessment by world region and Human Development Index using GLOBOCAN 2020. Int. J. Cancer 2022, 151, 1535–1541. [Google Scholar] [CrossRef]
- Tanha, K.; Mottaghi, A.; Nojomi, M.; Moradi, M.; Rajabzadeh, R.; Lotfi, S.; Janani, L. Investigation on factors associated with ovarian cancer: An umbrella review of systematic review and meta-analyses. J. Ovarian Res. 2021, 14, 153. [Google Scholar] [CrossRef]
- Singh, D.; Vignat, J.; Lorenzoni, V.; Eslahi, M.; Ginsburg, O.; Lauby-Secretan, B.; Arbyn, M.; Basu, P.; Bray, F.; Vaccarella, S. Global estimates of incidence and mortality of cervical cancer in 2020: A baseline analysis of the WHO Global Cervical Cancer Elimination Initiative. Lancet Glob. Health 2023, 11, e197–e206. [Google Scholar] [CrossRef]
- Pimple, S.; Mishra, G. Cancer cervix: Epidemiology and disease burden. Cytojournal 2022, 19, 21. [Google Scholar] [CrossRef]
- Yu, S.; Katz, O.; Fang, W.; Li, D.; Sang, W.; Liu, C. Shift of fleshy fruited species along elevation: Temperature, canopy coverage, phylogeny and origin. Sci. Rep. 2017, 7, 40417. [Google Scholar] [CrossRef] [PubMed]
- Fenn, M.A.; Giovannoni, J.J. Phytohormones in fruit development and maturation. Plant J. 2021, 105, 446–458. [Google Scholar] [CrossRef]
- Obroucheva, N.V. Hormonal regulation during plant fruit development. Biol. Plant Dev. 2014, 45, 11–21. [Google Scholar] [CrossRef]
- Martău, G.A.; Bernadette-Emőke, T.; Odocheanu, R.; Soporan, D.A.; Bochis, M.; Simon, E.; Vodnar, D.C. Vaccinium species (Ericaceae): Phytochemistry and biological properties of medicinal plants. Molecules 2023, 28, 1533. [Google Scholar] [CrossRef]
- Kopystecka, A.; Kozioł, I.; Radomska, D.; Bielawski, K.; Bielawska, A.; Wujec, M. Vaccinium uliginosum and Vaccinium myrtillus-two species-one used as a functional food. Nutrients 2023, 15, 4119. [Google Scholar] [CrossRef] [PubMed]
- Albert, N.W.; Iorizzo, M.; Mengist, M.F.; Montanari, S.; Zalapa, J.; Maule, A.; Edger, P.P.; Yocca, A.E.; Platts, A.E.; Pucker, B.; et al. Vaccinium as a comparative system for understanding of complex flavonoid accumulation profiles and regulation in fruit. Plant Physiol. 2023, 192, 1696–1710. [Google Scholar] [CrossRef] [PubMed]
- Skrovankova, S.; Sumczynski, D.; Mlcek, J.; Jurikova, T.; Sochor, J. Bioactive compounds and antioxidant activity in different types of berries. Int. J. Mol. Sci. 2015, 16, 24673–24706. [Google Scholar] [CrossRef]
- Karppinen, K.; Zoratti, L.; Nguyenquynh, N.; Häggman, H.; Jaakola, L. On the developmental and environmental regulation of secondary metabolism in Vaccinium spp. berries. Front. Plant Sci. 2016, 7, 655. [Google Scholar] [CrossRef]
- Määttä-Riihinen, K.R.; Kähkönen, M.P.; Törrönen, A.R.; Heinonen, I.M. Catechins and procyanidins in berries of vaccinium species and their antioxidant activity. J. Agric. Food Chem. 2005, 53, 8485–8491. [Google Scholar] [CrossRef]
- Suvanto, J.; Karppinen, K.; Riihinen, K.; Jaakola, L.; Salminen, J. Changes in the proanthocyanidin composition and related gene expression in bilberry (Vaccinium myrtillus L.) tissues. J. Agric. Food Chem. 2020, 68, 7378–7386. [Google Scholar] [CrossRef] [PubMed]
- Dare, A.P.; Günther, C.S.; Grey, A.C.; Guo, G.; Demarais, N.J.; Cordiner, S.; McGhie, T.K.; Boldingh, H.; Hunt, M.; Deng, C.; et al. Resolving the developmental distribution patterns of polyphenols and related primary metabolites in bilberry (Vaccinium myrtillus) fruit. Food Chem. 2021, 374, 131703. [Google Scholar] [CrossRef] [PubMed]
- Karppinen, K.; Hirvelä, E.; Nevala, T.; Sipari, N.; Suokas, M.; Jaakola, L. Changes in the abscisic acid levels and related gene expression during fruit development and ripening in bilberry (Vaccinium myrtillus L.). Phytochemistry 2013, 95, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Karppinen, K.; Tegelberg, P.; Häggman, H.; Jaakola, L. Abscisic acid regulates anthocyanin biosynthesis and gene expression associated with cell wall modificationin ripening bilberry (Vaccinium myrtillus L.) fruits. Front. Plant Sci. 2018, 9, 1259. [Google Scholar] [CrossRef] [PubMed]
- Zifkin, M.; Jin, A.; Ozga, J.A.; Zaharia, L.I.; Schernthaner, J.P.; Gesell, A.; Abrams, S.R.; Kennedy, J.A.; Constabel, P.C. Gene expression and metabolite profiling of developing highbush blueberry fruit indicates transcriptional regulation of flavonoid metabolism and activation of abscisic acid metabolism. Plant Physiol. 2012, 158, 200–224. [Google Scholar] [CrossRef] [PubMed]
- Lätti, A.K.; Jaakola, L.; Riihinen, K.R.; Kainulainen, P.S. Anthocyanin and flavonol variation in bog bilberries (Vaccinium uliginosum L.) in Finland. J. Agric. Food Chem. 2010, 58, 427–433. [Google Scholar] [CrossRef]
- Cesonienė, L.; Daubaras, R.; Jasutienė, I.; Venclovienė, J.; Miliauskienė, I. Evaluation of the biochemical components and chromatic properties of the juice of Vaccinium macrocarpon Aiton and Vaccinium oxycoccos L. Plant Foods Hum. Nutr. 2011, 66, 238–244. [Google Scholar] [CrossRef]
- Ge, Q.; Li, J.; Yang, F.; Tian, X.; Zhang, M.; Hao, Z.; Liang, C.; Meng, J. Molecular classifications of prostate cancer: Basis for individualized risk stratification and precision therapy. Ann. Med. 2023, 55, 2279235. [Google Scholar] [CrossRef] [PubMed]
- Fong, M.H.Y.; Feng, M.; McConkey, D.J.; Choi, W. Update on bladder cancer molecular subtypes. Transl. Androl. Urol. 2020, 9, 2881–2889. [Google Scholar] [CrossRef]
- Köbel, M.; Kang, E.Y. The evolution of ovarian carcinoma subclassification. Cancers 2022, 14, 416. [Google Scholar] [CrossRef]
- Marchocki, Z.; Swift, B.; Covens, A. Small cell and other rare histologic types of cervical cancer. Curr. Oncol. Rep. 2022, 24, 1531–1539. [Google Scholar] [CrossRef] [PubMed]
- Alsharairi, N.A. Experimental studies on the therapeutic potential of Vaccinium berries in breast cancer—A Review. Plants 2024, 13, 153. [Google Scholar] [CrossRef] [PubMed]
- Weh, K.M.; Clarke, J.; Kresty, L.A. Cranberries and cancer: An update of preclinical studies evaluating the cancer inhibitory potential of cranberry and cranberry derived constituents. Antioxidants 2016, 5, 27. [Google Scholar] [CrossRef] [PubMed]
- Boivin, D.; Blanchette, M.; Barrette, S.; Moghrabi, A.; Béliveau, R. Inhibition of cancer cell proliferation and suppression of TNF-induced activation of NFkappaB by edible berry juice. Anticancer. Res. 2007, 27, 937–948. [Google Scholar] [PubMed]
- Matchett, M.D.; MacKinnon, S.L.; Sweeney, M.I.; Gottschall-Pass, K.T.; Hurta, R.A.R. Blueberry flavonoids inhibit matrix metalloproteinase activity in DU145 human prostate cancer cells. Biochem. Cell Biol. 2005, 83, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Matchett, M.D.; MacKinnon, S.L.; Sweeney, M.I.; Gottschall-Pass, K.T.; Hurta, R.A.R. Inhibition of matrix metalloproteinase activity in DU145 human prostate cancer cells by flavonoids from lowbush blueberry (Vaccinium angustifolium): Possible roles for protein kinase C and mitogen-activated protein-kinase-mediated events. J. Nutr. Biochem. 2006, 17, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.M.; Howell, A.B.; McEniry, B.; Knight, C.T.; Seigler, D.; Erdman, J.W., Jr.; Lila, M.A. Effective separation of potent antiproliferation and antiadhesion components from wild blueberry (Vaccinium angustifolium Ait.) fruits. J. Agric. Food Chem. 2004, 52, 6433–6442. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.M.; Erdman, J.W., Jr.; Lila, M.A. Differential effects of blueberry proanthocyanidins on androgen sensitive and insensitive human prostate cancer cell lines. Cancer Lett. 2006, 231, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Adams, L.S.; Zhang, Y.; Lee, R.; Sand, D.; Scheuller, H.S.; Heber, D. Blackberry, black raspberry, blueberry, cranberry, red raspberry, and strawberry extracts inhibit growth and stimulate apoptosis of human cancer cells in vitro. J. Agric. Food Chem. 2006, 54, 9329–9339. [Google Scholar] [CrossRef]
- Wedge, D.E.; Meepagala, K.M.; Magee, J.B.; Smith, S.H.; Huang, G.; Larcom, L.L. Anticarcinogenic activity of strawberry, blueberry, and raspberry extracts to breast and cervical cancer cells. J. Med. Food 2001, 4, 49–51. [Google Scholar] [CrossRef]
- Davidson, K.T.; Zhu, Z.; Bai, Q.; Xiao, H.; Wakefield, M.R.; Fang, Y. Blueberry as a potential radiosensitizer for treating cervical cancer. Pathol. Oncol. Res. 2019, 25, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.; Liu, Y.; Liu, J.; Wang, E. Stability of blueberry anthocyanin, anthocyanidin and pyranoanthocyanidin pigments and their inhibitory effects and mechanisms in human cervical cancer HeLa cells. RSC Adv. 2019, 9, 10842–10853. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Wang, J.; Wang, W.; Lyu, L.; Wu, W.; Li, W. The extraction and high antiproliferative effect of anthocyanin from Gardenblue blueberry. Molecules 2023, 28, 2850. [Google Scholar] [CrossRef] [PubMed]
- Diaconeasa, Z.; Leopold, L.; Rugină, D.; Ayvaz, H.; Socaciu, C. Antiproliferative and antioxidant properties of anthocyanin rich extracts from blueberry and blackcurrant juice. Int. J. Mol. Sci. 2015, 16, 2352–2365. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Li, Z. Blueberries inhibit cyclooxygenase-1 and cyclooxygenase-2 activity in human epithelial ovarian cancer. Oncol. Lett. 2017, 13, 4897–4904. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, P.J.; Kurowska, E.; Freeman, D.J.; Chambers, A.F.; Koropatnick, D.J. A flavonoid fraction from cranberry extract inhibits proliferation of human tumor cell lines. J. Nutr. 2004, 134, 1529–1535. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Adams, L.S.; Hardy, M.L.; Heber, D. Total cranberry extract versus its phytochemical constituents: Antiproliferative and synergistic effects against human tumor cell lines. J. Agric. Food Chem. 2004, 52, 2512–2517. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, P.J.; Kurowska, E.M.; Freeman, D.J.; Chambers, A.F.; Koropatnick, J. In vivo inhibition of growth of human tumor lines by flavonoid fractions from cranberry extract. Nutr. Cancer 2006, 56, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Déziel, B.A.; Patel, K.; Neto, C.; Gottschall-Pass, K.; Hurta, R.A.R. Proanthocyanidins from the American Cranberry (Vaccinium macrocarpon) inhibit matrix metalloproteinase-2 and matrix metalloproteinase-9 activity in human prostate cancer cells via alterations in multiple cellular signalling pathways. J. Cell Biochem. 2010, 111, 742–754. [Google Scholar] [CrossRef]
- Déziel, B.; MacPhee, J.; Patel, K.; Catalli, A.; Kulka, M.; Neto, C.; Gottschall-Pass, K.; Hurta, R. American cranberry (Vaccinium macrocarpon) extract affects human prostate cancer cell growth via cell cycle arrest by modulating expression of cell cycle regulators. Food Funct. 2012, 3, 556–564. [Google Scholar] [CrossRef]
- MacLean, M.A.; Scott, B.E.; Deziel, B.A.; Nunnelley, M.C.; Liberty, A.M.; Gottschall-Pass, K.T.; Neto, C.C.; Hurta, R.A.R. North American cranberry (Vaccinium macrocarpon) stimulates apoptotic pathways in DU145 human prostate cancer cells in vitro. Nutr. Cancer 2011, 63, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; MacKinnon, S.L.; Craft, C.C.; Matchett, M.D.; Hurta, R.A.R.; Neto, C.C. Ursolic acid and its esters: Occurrence in cranberries and other Vaccinium fruit and effects on matrix metalloproteinase activity in DU145 prostate tumor cells. J. Sci. Food Agric. 2011, 91, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Prasain, J.K.; Jones, K.; Moore, R.; Barnes, S.; Leahy, M.; Roderick, R.; Juliana, M.M.; Grubbs, C.J. Effect of cranberry juice concentrate on chemically-induced urinary bladder cancers. Oncol. Rep. 2008, 19, 1565–1570. [Google Scholar] [PubMed]
- Prasain, J.K.; Rajbhandari, R.; Keeton, A.B.; Piazza, G.A.; Barnes, S. Metabolism and growth inhibitory activity of cranberry derived flavonoids in bladder cancer cells. Food Funct. 2016, 7, 4012–4019. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.P.; Singh, R.K.; Kim, K.K.; Satyan, K.S.; Nussbaum, R.; Torres, M.; Brard, L.; Vorsa, N. Cranberry proanthocyanidins are cytotoxic to human cancer cells and sensitize platinum-resistant ovarian cancer cells to paraplatin. Phytother. Res. 2009, 23, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Singh, A.P.; Singh, R.K.; Demartino, A.; Brard, L.; Vorsa, N.; Lange, T.S.; Moore, R.G. Anti-angiogenic activity of cranberry proanthocyanidins and cytotoxic properties in ovarian cancer cells. Int. J. Oncol. 2012, 40, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Han, A.; Chen, E.; Singh, R.K.; Chichester, C.O.; Moore, R.G.; Singh, A.P.; Vorsa, N. The cranberry flavonoids PAC DP-9 and quercetin aglycone induce cytotoxicity and cell cycle arrest and increase cisplatin sensitivity in ovarian cancer cells. Int. J. Oncol. 2015, 46, 1924–1934. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.; Pickles, T.; D’yachkova, Y. A randomised trial of cranberry versus apple juice in the management of urinary symptoms during external beam radiation therapy for prostate cancer. Clin. Oncol. (R Coll. Radiol.) 2003, 15, 322–328. [Google Scholar] [CrossRef]
- Herst, P.M.; Aumata, A.; Sword, V.; Jones, R.; Purdie, G.; Costello, S. Cranberry capsules are not superior to placebo capsules in managing acute non-haemorrhagic radiation cystitis in prostate cancer patients: A phase III double blinded randomised placebo controlled clinical trial. Radiother. Oncol. 2020, 149, 117–123. [Google Scholar] [CrossRef]
- Cowan, C.C.; Hutchison, C.; Cole, T.; Barry, S.J.E.; Paul, J.; Reed, N.S.; Russell, J.M. A randomised double-blind placebo-controlled trial to determine the effect of cranberry juice on decreasing the incidence of urinary symptoms and urinary tract infections in patients undergoing radiotherapy for cancer of the bladder or cervix. Clin. Oncol. (R Coll. Radiol.) 2012, 24, e31–e38. [Google Scholar] [CrossRef]
- Hamilton, K.; Bennett, N.C.; Purdie, G.; Herst, P.M. Standardized cranberry capsules for radiation cystitis in prostate cancer patients in New Zealand: A randomized double blinded, placebo controlled pilot study. Support. Care Cancer 2015, 23, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Bonetta, A.; Roviello, G.; Generali, D.; Zanotti, L.; Cappelletti, M.R.; Pacifico, C.; Di Pierro, F. Enteric-coated and highly standardized cranberry extract reduces antibiotic and nonsteroidal anti-inflammatory drug use for urinary tract infections during radiotherapy for prostate carcinoma. Res. Rep. Urol. 2017, 9, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Student, V.; Vidlar, A.; Bouchal, J.; Vrbkova, J.; Kolar, Z.; Kral, M.; Kosina, P.; Vostalova, J. Cranberry intervention in patients with prostate cancer prior to radical prostatectomy. Clinical, pathological and laboratory findings. Biomed. Pap. Med. Fac. Univ. Palacky. Olomouc Czech Repub. 2016, 160, 559–565. [Google Scholar] [CrossRef]
- Del Bubba, M.; Di Serio, C.; Renai, L.; Scordo, C.V.A.; Checchini, L.; Ungar, A.; Tarantini, F.; Bartoletti, R. Vaccinium myrtillus L. extract and its native polyphenol-recombined mixture have anti-proliferative and pro-apoptotic effects on human prostate cancer cell lines. Phytother. Res. 2021, 35, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Agrawal, A.K.; Mudd, A.M.; Kyakulaga, A.; Singh, I.P.; Vadhanam, M.V.; Gupta, R.C. Exosomal formulation of anthocyanidins against multiple cancer types. Cancer Lett. 2017, 393, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Aqil, F.; Jeyabalan, J.; Agrawal, A.K.; Kyakulaga, A.; Munagala, R.; Parker, L.; Gupta, R.C. Exosomal delivery of berry anthocyanidins for the management of ovarian cancer. Food Funct. 2017, 8, 4100–4107. [Google Scholar] [CrossRef]
- McDougall, G.J.; Ross, H.A.; Ikeji, M.; Stewart, D. Berry extracts exert different antiproliferative effects against cervical and colon cancer cells grown in vitro. J. Agric. Food Chem. 2008, 56, 3016–3023. [Google Scholar] [CrossRef]


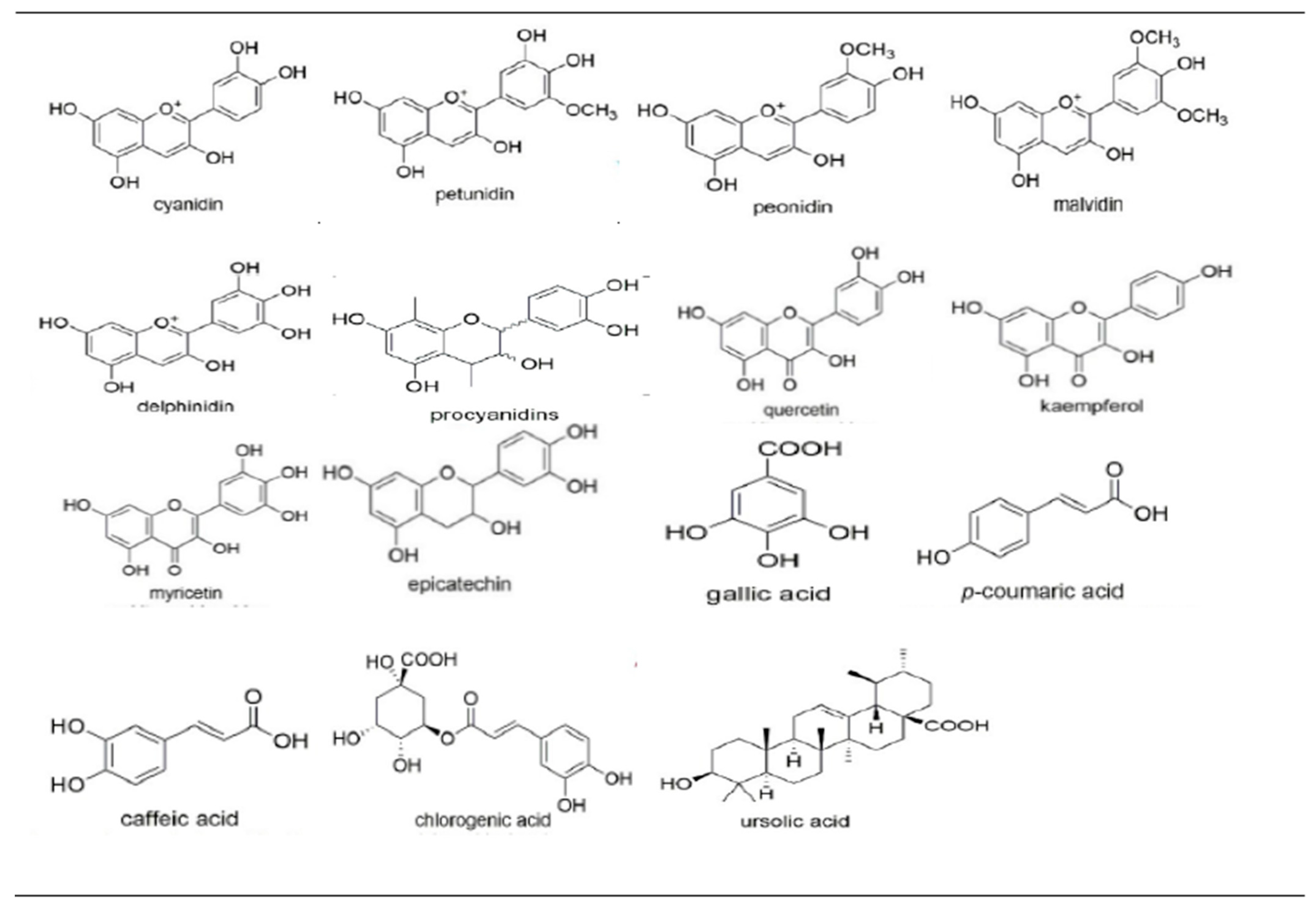{kind=link}
| Cancer Models | Cancer Types | Cell Lines | Bioactive Extracts | Treatment | Therapeutic Role | Mechanisms of Action | Ref. |
|---|---|---|---|---|---|---|---|
| In vitro | PCa | PC-3 | NA | PC-3 cells were treated with 0, 10, 20, 30, 40, and 50 μL/mL of three different blueberries and incubated for 48 h | Inhibition of cell proliferation and inflammation Induction of cell cycle arrest | cdk4/6, cyclin D1/D3, TNFα, COX-2, NFκB ↓ | [34] |
| In vitro | PCa | DU145 | ACNs (cyanidin, peonidin, petunidin, delphinidin, and malvidin), flavonol (Qu), PACs | DU145 cells were treated with 0.1, 0.5, and 1.0 mg/mL fractions of lowbush blueberries and incubated for 24 h | Inhibition of metastasis formation Induction of cell apoptosis | caspase-3 ↑, MMP-2/9 ↓ | [35] |
| In vitro | PCa | DU145 | ACNs (cyanidin, peonidin, petunidin, delphinidin, and malvidin), flavonol (Qu), PACs | DU145 cells were treated with 0.1, 0.5, and 1.0 mg/mL fractions of lowbush blueberries and incubated for 24 h | Inhibition of metastasis formation | TIMP-1/2 ↑, MMP-2/9, PKA, MAP ↓ | [36] |
| In vitro | PCa | LNCaP | PACs | LNCaP cells were treated with 5, 10, 12, 15, 20, 30, and 40 μg/mL of PAC-rich fractions of wild blueberry and 2.38 mM QU standard and incubated for 24 h | Inhibition of cell proliferation and adhesion | NA | [37] |
| In vitro | PCa | LNCaP, DU145 | PACs | PCa cells were treated with 0, 20, 40, 60, 80, 100, and 120 μg/mL of PAC-rich fractions of two blueberry fruits and 2.38 mM QU standard and incubated for 24 h | Inhibition of cell proliferation | NA | [38] |
| In vitro | PCa | LNCaP | ACNs (cyanidin, peonidin, petunidin, delphinidin, and malvidin), flavonol (QU, Km), PACs | LNCaP cells were treated with blueberry extracts ranging from 25 to 200 μg/mL in concentration and incubated for 48 h | Inhibition of cell proliferation | NA | [39] |
| In vitro | CC | CaSki, SiHa | NA | CC cells were treated with different concentrations of four extracts (ethyl acetate, hexane/ethyl acetate, acetone/water, and ethanol) of Tifblue and Premier blueberries and incubated for 48 h. | Inhibition of mutagenesis | Carcinogen formation by methyl methanesulfonate ↓ | [40] |
| In vitro | CC | SiHa | NA | SiHa cells were treated with 50 mg/mL blueberry extracts for 24 h, followed by radiotherapy at 4 Gy for 48 h | Inhibition of cell proliferation Induction of cell apoptosis | caspase-3, TUNEL+ cells, TRAIL ↑, cyclin D/E ↓ | [41] |
| In vitro | CC | HeLa | ACNs (cyanidin, peonidin, petunidin, delphinidin, and malvidin), anthocyanidin, proanthocyanidin | HeLa cells were treated with 0, 100, 200, 400, and 600 μg/mL−1 of lowbush blueberry extracts and incubated for 24 h | Inhibition of cell proliferation and viability Induction of apoptosis and cell cycle arrest | caspase-3, p53, p38 MAPK ↑, cyclin D1 ↓ | [42] |
| In vitro | CC | HeLa | ACNs (cyanidin, peonidin, petunidin, delphinidin, and malvidin) | HeLa cells were treated with 52 μg/mL of Gardenblue blueberry ACNs and chemotherapeutic drugs (cisplatin and doxorubicin) | Inhibition of cell proliferation | NA | [43] |
| In vitro | OC and CC | A2780 (OC), HeLa (CC) | ACNs (cyanidin, peonidin, petunidin, delphinidin, and malvidin) | Cells were treated with 0, 50, 100, 150, 200, 250, 300, 350, 400, 450, and 500 μg/mL of ACN-rich fractions of blueberry juice and incubated for 24 h | Inhibition of cell proliferation and viability | NA | [44] |
| In vivo | OC | SKOV3 | NA | BALB/c mice were fed daily with 100, 200, and 400 mg/kg of blueberry juice. SKOV3 cells were treated with 0, 1, 2, 4, 8, or 16 mg/mL of blueberry juice and incubated for 24, 48, or 72 h | Inhibition of cell proliferation | COX-1/2 ↓ | [45] |
| Cancer Models | Cancer Types | Cell Lines | Bioactive Extracts | Treatment | Therapeutic Role | Mechanisms of Action | Ref. |
|---|---|---|---|---|---|---|---|
| In vitro | PCa | LNCaP, DU145 | ACNs, flavonols, PACs, flavan-3-ols (epicatechin) | PCa cells were treated with fractions of cranberry press cake extract (0, 100, 200, 300, 400, 500, and 600 mg/mL) and incubated for 4d | Inhibition of cell proliferation | NA | [46] |
| In vitro | PCa | RWPE-1, RWPE-2, 22Rv1 | ACNs (cyanidin, peonidin), PACs, total polyphenols | PCa cells were treated with fractions of total cranberry extract (50, 100, and 200 μg/mL) and incubated for 48 h | Inhibition of cell proliferation | NA | [47] |
| In vivo | PCa | DU145 | PACs | Female Balb/c mice were injected with 100 mg/kg PACs 3, 5, 7, 10, 12, 14, 17, 19, and 21 days after tumour implant. | Inhibition of tumour growth | NA | [48] |
| In vitro | PCa | DU145 | PACs | DU145 cells were treated with 0–25 μg/mL of cranberry PACs and incubated for 24 h | Inhibition of cell viability | TIMP-2, p38, ERK 1/2, c-jun ↑, MMP-2/9, EMMPRIN, PI3K, Akt, NFkB p65, c-fos ↓ | [49] |
| In vitro | PCa | DU145 | ACNs (cyanidin, peonidin), flavonols (Qu), PACs | DU145 cells were treated with whole cranberry extracts (10, 25, and 50 μg/mL−1) and incubated for 24 h | Inhibition of cell viability Induction of cell cycle arrest | p27 ↑, CDK4, cyclin A/B1/D1/E ↓ | [50] |
| In vitro | PCa | DU145 | ACNs (cyanidin, peonidin), flavonols (Qu, Myr), PACs | DU145 cells were treated with flavonol and PAC-enriched fractions of cranberry (10, 25, 5, and 100 μg/mL) and incubated for 24 h | Induction of cell apoptosis | caspase-8/9, par-4, cytochrome-C, tBid, Bax ↑ | [51] |
| In vitro | PCa | DU145 | Ursolic acid | DU145 cells were treated with 1 μg/mL−1 and 10 μg/mL−1 of ursolic acid and its cis and trans-3-O-p-hydroxycinnamoyl esters extracted from cranberries and incubated for 24 h | Inhibition of tumour growth, invasion, and metastasis | MMP-2/9 ↓ | [52] |
| In vivo | BC | NA | NA | Female Fischer rats were injected with 1.0 or 0.5 mL/day of cranberry juice concentrate for a period of six weeks | Inhibition of tumour growth and cell proliferation | NA | [53] |
| In vivo | BC | RT4, SCABER, SW-780 | Flavonols (Qu, aglycone Qu, Myr) | BC cells were treated with cranberry-derived flavonoid concentrations ranging from 0.3–200 μM and incubated for 72 h | Inhibition of tumour growth and cell proliferation | NA | [54] |
| In vitro | OC | SKOV-3 | PACs | SKOV-3 cells were treated with cranberry PAC fractions (concentrations ranging from 0–150 μg/mL) and 4.5 μg/mL paraplatin and incubated for 48 h | Inhibition of cell proliferation and viability Induction of cell apoptosis | NA | [55] |
| In vitro | OC | SKOV-3 | PACs | SKOV-3 cells were treated with cranberry PACs (concentrations ranging from 12.5–100 μg/mL) and incubated for 24 h | Inhibition of cell proliferation, viability, and angiogenesis Induction of cell cycle arrest and apoptosis | ROS, caspase-3/7/8 ↑, Akt, PARP, VEGF ↓ | [56] |
| In vitro | OC | SKOV-3, OVCAR-8 | PACs, Qu aglycone | OC cells were treated with cranberry PACs and Qu aglycone concentrations ranging from 0–200 μg/mL and incubated for 24–48 h | Inhibition of cell viability Induction of cell cycle arrest and apoptosis | caspase-3, p21, p27, CDK-2 ↑, PARP, EGFR, MAPK, ERK, RAF, cyclin D1, phospho-histone H3, DNA-PK ↓ | [57] |
| Study Characteristics | Study Focus | Intervention | Effects | Ref. |
|---|---|---|---|---|
| Total subjects = 112 PCa patients (cranberry group = 55, apple group = 57) | Urinary symptoms | Patients consumed 354 mL of cranberry juice or apple juice a day over two weeks during radiation treatment | No significant effects on urinary symptoms were observed | [58] |
| Total subjects = 101 PCa patients (cranberry capsules group = 51, placebo = 50) | Cystitis symptoms | Patients received two capsules containing cranberry PACs or a beetroot-containing placebo twice a day during radiation treatment and two weeks after | Patients in the cranberry arm developed worse radiation cystitis symptoms compared with those in the placebo arm | [59] |
| Total subjects = 128 BC and CC patients (cranberry group = 64, placebo = 64) | Urinary symptoms, UTIs | Patients received ≥16,000 mL of cranberry juice or a placebo beverage twice a day over four weeks during radiation treatment and two weeks after | Patients in the placebo arm experienced more urinary symptoms and UTIs during and after radiation treatment compared with those in the treatment arm | [60] |
| Total subjects = 40 PCa patients (cranberry capsules group = 20, placebo = 20) | Cystitis symptoms | Patients received one capsule containing cranberry PACs or a magnesium stearate, gelatin, colloidal silica, and gelatin-containing placebo once a day during radiation treatment and two weeks after | Patients in the cranberry arm had fewer severe cystitis symptoms compared with those in the placebo arm during and after radiation treatment | [61] |
| Total subjects = 924 PCa patients (enteric-coated, highly standardized cranberry group = 489, untreated group = 435) | UTIs | Patients treated with one tablet/day of 200 mg of highly standardized cranberry extract over 6–7 weeks of radiation treatment | Patients demonstrated a significant reduction in urinary discomfort and the use of antibiotics/anti-inflammatory drugs | [62] |
| Total subjects = 64 PCa patients (cranberry group = 32, placebo = 32) | Serum prostate-specific antigen; blood, urine, and prostate tissue markers | Patients received 1500 mg of cranberry fruit powder daily for one month, or one capsule containing 500 mg of canola oil, Blue 1 Lake, sodium aluminium silicate, STAR-DRI® 1015A (Tate & Lyle Solutions, Sycamore, IL, USA) maltodextrin (placebo) 21 days before surgery | Patients in the cranberry arm showed a significant reduction in serum prostate-specific antigen, serum gamma-glutamyltranspeptidase, and urinary beta-microseminoprotein, along with upregulated insulin-like growth factor-1 compared with those in the placebo arm | [63] |
| Cancer Models | Cancer Types | Cell Lines | Vaccinium Species | Bioactive Extracts | Treatment | Therapeutic Role | Mechanisms of Action | Ref. |
|---|---|---|---|---|---|---|---|---|
| In vitro | PCa | LNCaP, DU145, PC3 | Bilberry | ACNs (cyanidin, peonidin, petunidin, delphinidin, and malvidin), flavonol (Qu and Myr), flavanol (epicatechin), phenolic acids (Gallic, p-Coumaric, caffeic, and chlorogenic acids) | PCa cells were treated with bilberry extract and a recombined standard mixture at distinct dilutions (1/100 v/v dilution, (1/400 v/v, and 1/800 v/v dilution), and incubated for 7d | Inhibition of tumour growth and cell proliferation Induction of cell apoptosis | Hypoxia ↑ | [64] |
| In vitro/vivo | PCa and OC | DU145, PC3 (PCa); OVCA432 (OC) | Bilberry | Anthos | Mice were randomized into vehicle (PBS), ExoAnthos (5 mg Anthos and 50 mg Exo protein/kg b.wt.), and Anthos (10 mg/kg b. wt.) groups PCa and OC cells were treated with various concentrations of ExoAnthos, Anthos, or exosomes and incubated for 72 h | Inhibition of tumour growth and cell proliferation | NA | [65] |
| In vitro/vivo | OC | A2780, A2780/CP70, OVCA432, OVCA433 | Bilberry | Anthos | Mice were treated, through oral gavage, with Anthos (6 and 30 mg kg−1 b. wt) and exosomal formula (mg kg−1 Anthos and 60 mg kg−1 Exo) (study 1); Mice were also treated with paclitaxel (4 mg kg−1 b. wt) and exosomal formula of paclitaxel (4 mg PAC kg−1 b. wt and 60 mg Exo kg−1 b. wt) OC cells were treated with different concentrations (0, 50 100, 150, 200, and 300 µM) of Anthos and paclitaxel and incubated for 72 h | Inhibition of tumour growth and cell proliferation | PgP ↓ | [66] |
| In vitro | CC | HeLa | Lingonberry | ACNs (cyanidin), tannin (procyanidins) | HeLa cells were treated with fractions of lingonberry extract (concentrations ranging from 25–75 μg/mL) and incubated for 72 h | Inhibition of cell proliferation and viability | NA | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsharairi, N.A. A Review with a Focus on Vaccinium-Berries-Derived Bioactive Compounds for the Treatment of Reproductive Cancers. Plants 2024, 13, 1047. https://doi.org/10.3390/plants13071047
Alsharairi NA. A Review with a Focus on Vaccinium-Berries-Derived Bioactive Compounds for the Treatment of Reproductive Cancers. Plants. 2024; 13(7):1047. https://doi.org/10.3390/plants13071047
Chicago/Turabian StyleAlsharairi, Naser A. 2024. "A Review with a Focus on Vaccinium-Berries-Derived Bioactive Compounds for the Treatment of Reproductive Cancers" Plants 13, no. 7: 1047. https://doi.org/10.3390/plants13071047
APA StyleAlsharairi, N. A. (2024). A Review with a Focus on Vaccinium-Berries-Derived Bioactive Compounds for the Treatment of Reproductive Cancers. Plants, 13(7), 1047. https://doi.org/10.3390/plants13071047