
Comprehensive Genome-Wide Natural Variation and Expression Analysis of Tubby-like Proteins Gene Family in Brachypodium distachyon

Abstract
:1. Introduction
2. Material and Methods
2.1. Sequence Identification of TLP Genes in Arabidopsis, Rice, Sorghum, and B. distachyon
2.2. Phylogenetic and Sequence Analysis of TLP Genes
2.3. Expression Analysis
2.4. Natural Variation and 3D Modeling of BdTLPs’ Variant Proteins
3. Results
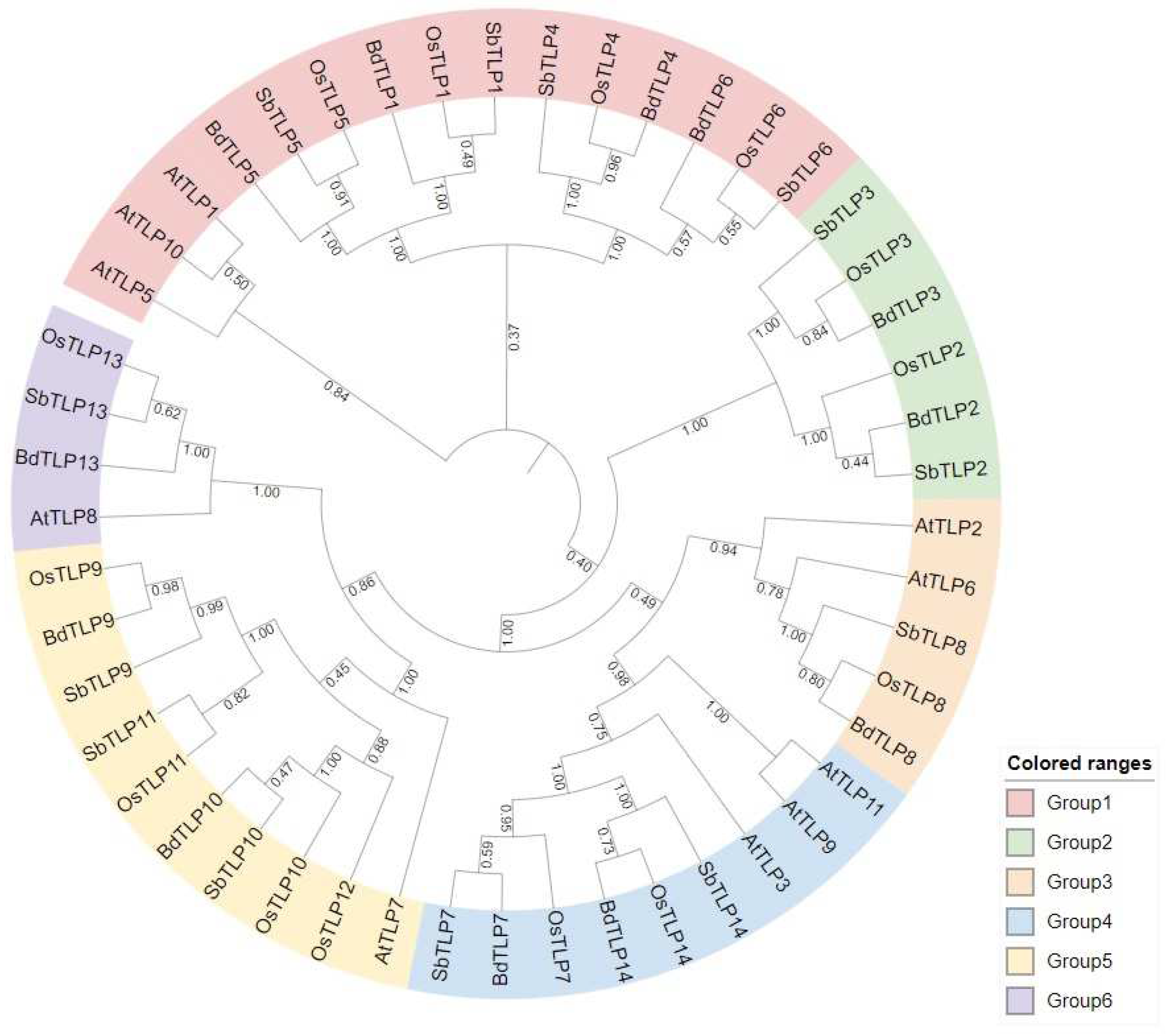
3.1. Classification and Phylogenetic Analysis of Arabidopsis thaliana, Oryza sativa, Sorghum bicolor, and Brachypodium distachyon TLP Genes
3.2. Multiple Sequence Alignment Analysis and Amino Acid Composition Bias of AtTLPs, OsTLPs, SbTLPs, and BdTLPs
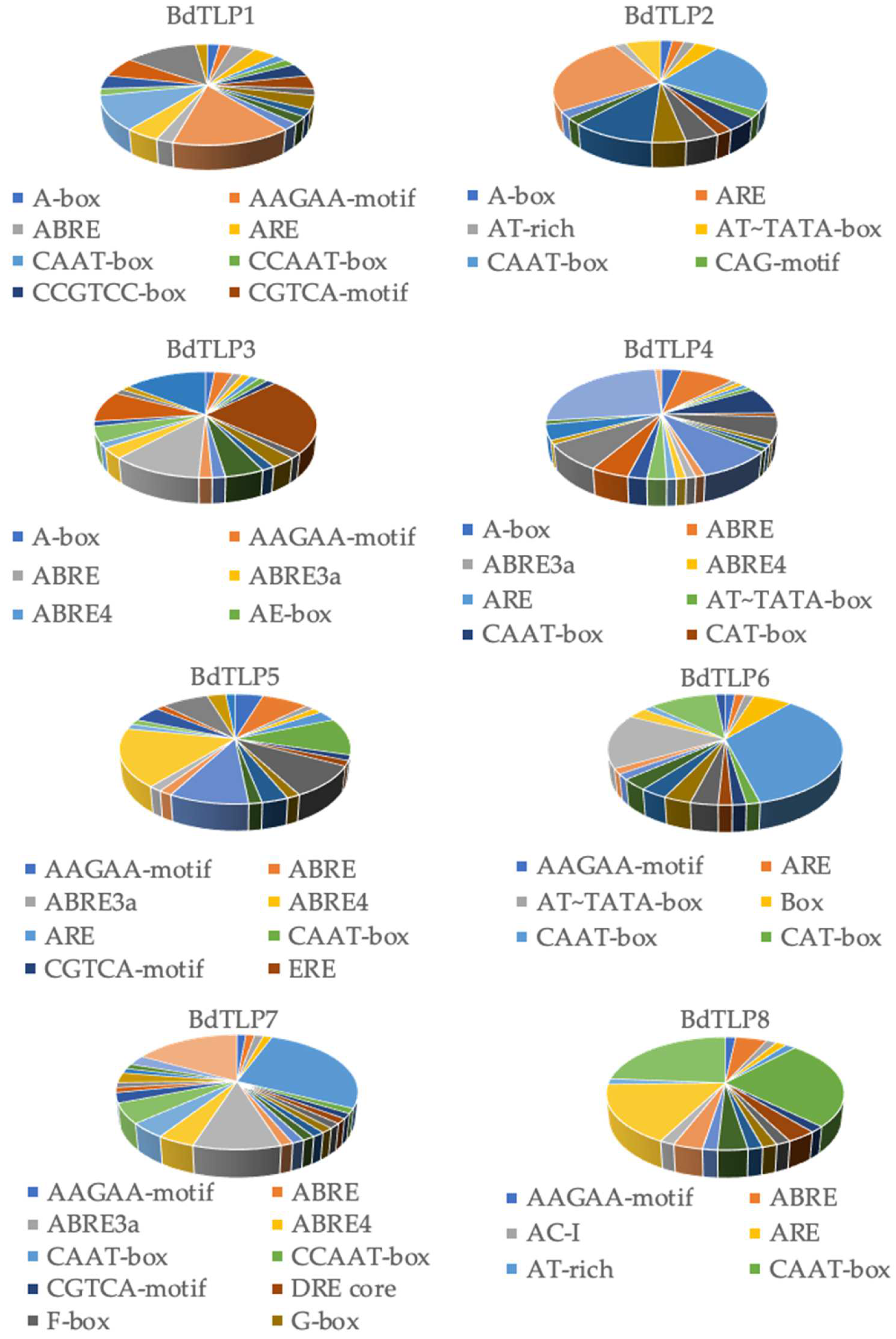
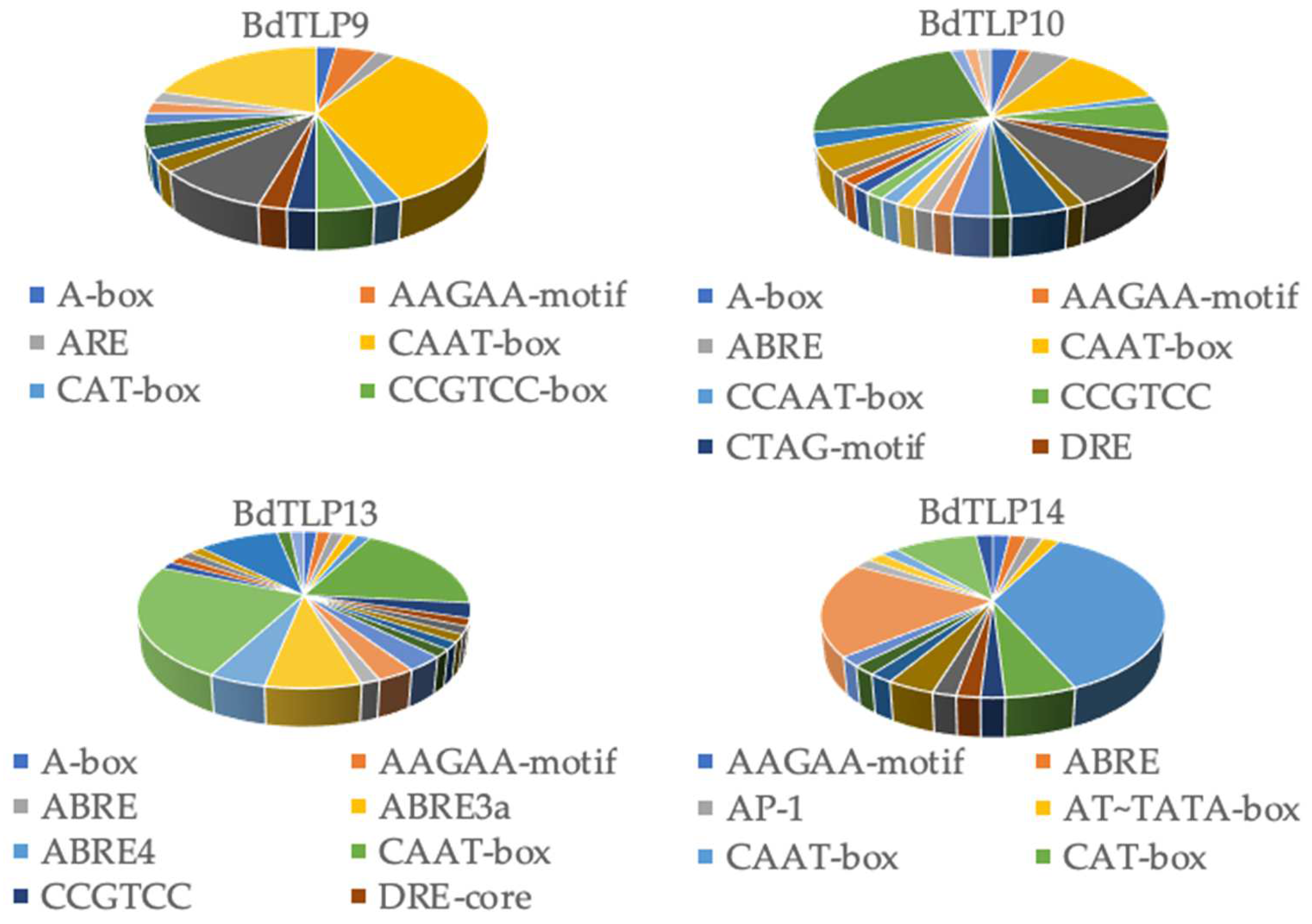
3.3. Cis Elements in BdTLP Promoters and Gene Expression during Developmental Stages
3.4. Natural Variation DNA and Protein Sequences and 3D Structures of BdTLPs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, M.; Xu, Z.; Kong, Y. The tubby-like proteins kingdom in animals and plants. Gene 2018, 642, 16–25. [Google Scholar] [CrossRef]
- Bano, N.; Aalam, S.; Bag, S.K. Tubby-like proteins (TLPs) transcription factor in different regulatory mechanism in plants: A review. Plant Mol. Biol. 2022, 110, 455–468. [Google Scholar] [CrossRef]
- Kleyn, P.W.; Fan, W.; Kovats, S.G.; Lee, J.J.; Pulido, J.C.; Wu, Y.; Berkemeier, L.R.; Misumi, D.J.; Holmgren, L.; Charlat, O.; et al. Identification and characterization of the mouse obesity gene tubby: A member of a novel gene family. Cell 1996, 85, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Noben-Trauth, K.; Naggert, J.K.; North, M.A.; Nishina, P.M. A candidate gene for the mouse mutation tubby. Nature 1996, 380, 534–538. [Google Scholar] [CrossRef]
- Ohlemiller, K.K.; Hughes, R.M.; Mosinger-Ogilvie, J.; Speck, J.D.; Grosof, D.H.; Silverman, M.S. Cochlear and retinal degeneration in the tubby mouse. Neuroreport 1995, 6, 845–849. [Google Scholar] [CrossRef] [PubMed]
- Badgandi, H.B.; Hwang, S.H.; Shimada, I.S.; Loriot, E.; Mukhopadhyay, S. Tubby family proteins are adapters for ciliary trafficking of integral membrane proteins. J. Cell Biol. 2017, 216, 743–760. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Wen, X.; Ratti, N.; Loktev, A.; Rangell, L.; Scales, S.J.; Jackson, P.K. The ciliary G-protein-coupled receptor Gpr161 negatively regulates the Sonic hedgehog pathway via cAMP signaling. Cell 2013, 152, 210–223. [Google Scholar] [CrossRef] [PubMed]
- Abd-Hamid, N.A.; Ahmad-Fauzi, M.I.; Zainal, Z.; Ismail, I. Diverse and dynamic roles of F-box proteins in plant biology. Planta 2020, 251, 68. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.J.; Kim, D.Y.; Seo, Y.W. Interactions between wheat Tubby-like and SKP1-like proteins. Genes Genet. Syst. 2015, 90, 293–304. [Google Scholar] [CrossRef]
- Bao, Y.; Song, W.M.; Jin, Y.L.; Jiang, C.M.; Yang, Y.; Li, B.; Huang, W.J.; Liu, H.; Zhang, H.X. Characterization of Arabidopsis Tubby-like proteins and redundant function of AtTLP3 and AtTLP9 in plant response to ABA and osmotic stress. Plant Mol. Biol. 2014, 86, 471–483. [Google Scholar] [CrossRef]
- Reitz, M.U.; Bissue, J.K.; Zocher, K.; Attard, A.; Huckelhoven, R.; Becker, K.; Imani, J.; Eichmann, R.; Schafer, P. The subcellular localization of Tubby-like proteins and participation in stress signaling and root colonization by the mutualist Piriformospora indica. Plant Physiol. 2012, 160, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Santagata, S.; Boggon, T.J.; Baird, C.L.; Gomez, C.A.; Zhao, J.; Shan, W.S.; Myszka, D.G.; Shapiro, L. G-protein signaling through tubby proteins. Science 2001, 292, 2041–2050. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xing, S.; Sun, Q.; Zhan, C.; Liu, X.; Zhang, S.; Wang, X. The expression of a tubby-like protein from Malus domestica (MdTLP7) enhances abiotic stress tolerance in Arabidopsis. BMC Plant Biol. 2019, 19, 60. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.N.; Xing, S.S.; Zhang, Z.R.; Chen, X.S.; Wang, X.Y. Genome-Wide Identification and Expression Analysis of the Tubby-like Protein Family in the Malus domestica Genome. Front. Plant Sci. 2016, 7, 1693. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.P.; Lee, C.L.; Chen, P.H.; Wu, S.H.; Yang, C.C.; Shaw, J.F. Molecular analyses of the Arabidopsis TUBBY-like protein gene family. Plant Physiol. 2004, 134, 1586–1597. [Google Scholar] [CrossRef]
- Zhang, Y.; He, X.; Su, D.; Feng, Y.; Zhao, H.; Deng, H.; Liu, M. Comprehensive Profiling of Tubby-Like Protein Expression Uncovers Ripening-Related TLP Genes in Tomato (Solanum lycopersicum). Int. J. Mol. Sci. 2020, 21, 1000. [Google Scholar] [CrossRef]
- Yang, Z.; Zhou, Y.; Wang, X.; Gu, S.; Yu, J.; Liang, G.; Yan, C.; Xu, C. Genomewide comparative phylogenetic and molecular evolutionary analysis of tubby-like protein family in Arabidopsis, rice, and poplar. Genomics 2008, 92, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Bano, N.; Fakhrah, S.; Mohanty, C.S.; Bag, S.K. Genome-Wide Identification and Evolutionary Analysis of Gossypium Tubby-like Protein (TLP) Gene Family and Expression Analyses during Salt and Drought Stress. Front. Plant Sci. 2021, 12, 667929. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.Y.; Fan, X.W.; Pang, X.Y.; Li, Y.Z. Decrypting tubby-like protein gene family of multiple functions in starch root crop cassava. AoB Plants 2019, 11, plz075. [Google Scholar] [CrossRef]
- Liu, Q. Identification of rice TUBBY-like genes and their evolution. FEBS J. 2008, 275, 163–171. [Google Scholar] [CrossRef]
- Chen, Y.; Dai, W.; Sun, B.; Zhao, Y.; Ma, Q. Genome-wide identification and comparative analysis of the TUBBY-like protein gene family in maize. Genes Genom. 2016, 38, 25–36. [Google Scholar]
- Xu, H.R.; Liu, Y.; Yu, T.F.; Hou, Z.H.; Zheng, J.C.; Chen, J.; Zhou, Y.B.; Chen, M.; Fu, J.D.; Ma, Y.Z.; et al. Comprehensive Profiling of Tubby-like Proteins in Soybean and Roles of the GmTLP8 Gene in Abiotic Stress Responses. Front. Plant Sci. 2022, 13, 844545. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Wen, J.; Fu, J.; Geng, H.; Dan, Z.; Zhao, W.; Xu, W.; Huang, W. Genome-wide identification and comprehensive analysis of tubby-like protein gene family in multiple crops. Front. Plant Sci. 2022, 13, 1093944. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hu, J.; Ma, X.; Li, C.; Yang, Q.; Feng, S.; Li, M.; Li, N.; Song, X. Identification, evolution and expression analyses of whole genome-wide TLP gene family in Brassica napus. BMC Genom. 2020, 21, 264. [Google Scholar] [CrossRef]
- Renak, D.; Dupl’akova, N.; Honys, D. Wide-scale screening of T-DNA lines for transcription factor genes affecting male gametophyte development in Arabidopsis. Sex. Plant Reprod. 2012, 25, 39–60. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Ni, W.; Feng, B.; Han, T.; Petrasek, M.G.; Ma, H. Members of the Arabidopsis-SKP1-like gene family exhibit a variety of expression patterns and may play diverse roles in Arabidopsis. Plant Physiol. 2003, 133, 203–217. [Google Scholar] [CrossRef]
- Li, S.; Wang, Z.; Wang, F.; Lv, H.; Cao, M.; Zhang, N.; Li, F.; Wang, H.; Li, X.; Yuan, X.; et al. A tubby-like protein CsTLP8 acts in the ABA signaling pathway and negatively regulates osmotic stresses tolerance during seed germination. BMC Plant Biol. 2021, 21, 340. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, X.; Cao, X.; Chen, B.; Ma, C.; Lv, J.; Sun, Z.; Qiao, K.; Zhu, L.; Zhang, C.; et al. GhTULP34, a member of tubby-like proteins, interacts with GhSKP1A to negatively regulate plant osmotic stress. Genomics 2021, 113, 462–474. [Google Scholar] [CrossRef] [PubMed]
- The International Brachypodium Initiative. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar] [CrossRef]
- Tyler, L.; Fangel, J.U.; Fagerstrom, A.D.; Steinwand, M.A.; Raab, T.K.; Willats, W.G.; Vogel, J.P. Selection and phenotypic characterization of a core collection of Brachypodium distachyon inbred lines. BMC Plant Biol. 2014, 14, 25. [Google Scholar] [CrossRef]
- Hasterok, R.; Catalan, P.; Hazen, S.P.; Roulin, A.C.; Vogel, J.P.; Wang, K.; Mur, L.A.J. Brachypodium: 20 years as a grass biology model system; the way forward? Trends Plant Sci. 2022, 27, 1002–1016. [Google Scholar] [CrossRef] [PubMed]
- Bettgenhaeuser, J.; Corke, F.M.; Opanowicz, M.; Green, P.; Hernandez-Pinzon, I.; Doonan, J.H.; Moscou, M.J. Natural Variation in Brachypodium Links Vernalization and Flowering Time Loci as Major Flowering Determinants. Plant Physiol. 2017, 173, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.P.; Bednarek, R.; Bouche, F.; Gordon, S.P.; Vogel, J.P.; Garvin, D.F.; Amasino, R.M. Genetic Architecture of Flowering-Time Variation in Brachypodium distachyon. Plant Physiol. 2017, 173, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Davidson, R.M.; Gowda, M.; Moghe, G.; Lin, H.; Vaillancourt, B.; Shiu, S.H.; Jiang, N.; Robin Buell, C. Comparative transcriptomics of three Poaceae species reveals patterns of gene expression evolution. Plant J. 2012, 71, 492–502. [Google Scholar] [CrossRef] [PubMed]
- Kakei, Y.; Mochida, K.; Sakurai, T.; Yoshida, T.; Shinozaki, K.; Shimada, Y. Transcriptome analysis of hormone-induced gene expression in Brachypodium distachyon. Sci. Rep. 2015, 5, 14476. [Google Scholar] [CrossRef] [PubMed]
- Saada, S.; Solomon, C.U.; Drea, S. Programmed Cell Death in Developing Brachypodium distachyon Grain. Int. J. Mol. Sci. 2021, 22, 9086. [Google Scholar] [CrossRef] [PubMed]
- Coudert, E.; Gehant, S.; de Castro, E.; Pozzato, M.; Baratin, D.; Neto, T.; Sigrist, C.J.A.; Redaschi, N.; Bridge, A.; UniProt, C. Annotation of biologically relevant ligands in UniProtKB using ChEBI. Bioinformatics 2023, 39, btac793. [Google Scholar] [CrossRef]
- Sigrist, C.J.; de Castro, E.; Cerutti, L.; Cuche, B.A.; Hulo, N.; Bridge, A.; Bougueleret, L.; Xenarios, I. New and continuing developments at PROSITE. Nucleic Acids Res. 2013, 41, D344–D347. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Eddy, S.R. HMMER User’s Guide; Department of Genetics, Washington University School of Medicine: St. Louis, MO, USA, 1992. [Google Scholar]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, K.B.; Nicholas, H.B., Jr.; Deerfield, D.W., II. GeneDoc: Analysis and Visualization of Genetic Variation. Embnet News 1997, 4, 14. [Google Scholar]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Smedley, D.; Haider, S.; Durinck, S.; Pandini, L.; Provero, P.; Allen, J.; Arnaiz, O.; Awedh, M.H.; Baldock, R.; Barbiera, G.; et al. The BioMart community portal: An innovative alternative to large, centralized data repositories. Nucleic Acids Res. 2015, 43, W589–W598. [Google Scholar] [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- George, N.; Fexova, S.; Fuentes, A.M.; Madrigal, P.; Bi, Y.; Iqbal, H.; Kumbham, U.; Nolte, N.F.; Zhao, L.; Thanki, A.S.; et al. Expression Atlas update: Insights from sequencing data at both bulk and single cell level. Nucleic Acids Res. 2023, 52, D107–D114. [Google Scholar] [CrossRef] [PubMed]
- Moreno, P.; Fexova, S.; George, N.; Manning, J.R.; Miao, Z.; Mohammed, S.; Munoz-Pomer, A.; Fullgrabe, A.; Bi, Y.; Bush, N.; et al. Expression Atlas update: Gene and protein expression in multiple species. Nucleic Acids Res. 2022, 50, D129–D140. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, M.C.; Konate, M.M.; Chen, L.; Das, B.; Karlovich, C.; Williams, P.M.; Evrard, Y.A.; Doroshow, J.H.; McShane, L.M. TPM, FPKM, or Normalized Counts? A Comparative Study of Quantification Measures for the Analysis of RNA-seq Data from the NCI Patient-Derived Models Repository. J. Transl. Med. 2021, 19, 269. [Google Scholar] [CrossRef]
- Randle, A.; Villanueva, M.; Chen, Z.J. ggplot2: Elegant Graphics for Data Analysis. Meas. Interdiscip. Res. Perspect. 2019, 17, 160–167. [Google Scholar]
- Patro, S.G.K.; Sahu, K.K. Normalization: A Preprocessing Stage. arXiv 2015, arXiv:1503:06462. [Google Scholar] [CrossRef]
- Gordon, S.P.; Contreras-Moreira, B.; Woods, D.P.; Des Marais, D.L.; Burgess, D.; Shu, S.; Stritt, C.; Roulin, A.C.; Schackwitz, W.; Tyler, L.; et al. Extensive gene content variation in the Brachypodium distachyon pan-genome correlates with population structure. Nat. Commun. 2017, 8, 2184. [Google Scholar] [CrossRef]
- Baek, M.; DiMaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D.; et al. Accurate prediction of protein structures and interactions using a three-track neural network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef]
- Zhang, C.; Shine, M.; Pyle, A.M.; Zhang, Y. US-align: Universal structure alignments of proteins, nucleic acids, and macromolecular complexes. Nat. Methods 2022, 19, 1109–1115. [Google Scholar] [CrossRef]
- Zhou, Y.; Pan, Q.; Pires, D.E.V.; Rodrigues, C.H.M.; Ascher, D.B. DDMut: Predicting effects of mutations on protein stability using deep learning. Nucleic Acids Res. 2023, 51, W122–W128. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.E.; Ascher, D.B.; Blundell, T.L. DUET: A server for predicting effects of mutations on protein stability using an integrated computational approach. Nucleic Acids Res. 2014, 42, W314–W319. [Google Scholar] [CrossRef] [PubMed]
- Orr, H.A. Fitness and its role in evolutionary genetics. Nat. Rev. Genet. 2009, 10, 531–539. [Google Scholar] [CrossRef]
- Qu, L.; Sun, M.; Li, X.; He, R.; Zhong, M.; Luo, D.; Liu, X.; Zhao, X. The Arabidopsis F-box protein FOF2 regulates ABA-mediated seed germination and drought tolerance. Plant Sci. 2020, 301, 110643. [Google Scholar] [CrossRef]
- Majee, M.; Kumar, S.; Kathare, P.K.; Wu, S.; Gingerich, D.; Nayak, N.R.; Salaita, L.; Dinkins, R.; Martin, K.; Goodin, M.; et al. KELCH F-BOX protein positively influences Arabidopsis seed germination by targeting PHYTOCHROME-INTERACTING FACTOR1. Proc. Natl. Acad. Sci. USA 2018, 115, E4120–E4129. [Google Scholar] [CrossRef]
- Chae, E.; Tan, Q.K.; Hill, T.A.; Irish, V.F. An Arabidopsis F-box protein acts as a transcriptional co-factor to regulate floral development. Development 2008, 135, 1235–1245. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Ito, S.; Song, Y.H.; Strait, A.A.; Kiba, T.; Lu, S.; Henriques, R.; Pruneda-Paz, J.L.; Chua, N.H.; Tobin, E.M.; et al. F-box proteins FKF1 and LKP2 act in concert with ZEITLUPE to control Arabidopsis clock progression. Plant Cell 2010, 22, 606–622. [Google Scholar] [CrossRef] [PubMed]
- Ariizumi, T.; Lawrence, P.K.; Steber, C.M. The role of two f-box proteins, SLEEPY1 and SNEEZY, in Arabidopsis gibberellin signaling. Plant Physiol. 2011, 155, 765–775. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Kim, H.S.; Kim, S.D.; Park, J.Y. Insulin Phosphorylates Tyrosine Residue 464 of Tub and Translocates Tubby into the Nucleus in HIRcB Cells. Endocrinol. Metab. 2014, 29, 163–168. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light | Hormone | Environmental Stress | Developmental | Promoter | Other | Total | |
|---|---|---|---|---|---|---|---|
| BdTLP1 | 6 | 7 | 3 | 0 | 6 | 24 | 46 |
| BdTLP2 | 2 | 1 | 1 | 3 | 23 | 17 | 47 |
| BdTLP3 | 5 | 7 | 2 | 1 | 21 | 27 | 63 |
| BdTLP4 | 13 | 11 | 3 | 1 | 14 | 47 | 89 |
| BdTLP5 | 9 | 10 | 4 | 0 | 18 | 23 | 64 |
| BdTLP6 | 2 | 4 | 3 | 4 | 32 | 18 | 63 |
| BdTLP7 | 5 | 7 | 5 | 0 | 23 | 31 | 71 |
| BdTLP8 | 5 | 4 | 3 | 0 | 25 | 22 | 59 |
| BdTLP9 | 1 | 1 | 3 | 2 | 17 | 20 | 44 |
| BdTLP10 | 9 | 4 | 4 | 1 | 9 | 41 | 68 |
| BdTLP13 | 3 | 2 | 4 | 0 | 27 | 28 | 64 |
| BdTLP14 | 3 | 4 | 3 | 3 | 29 | 11 | 53 |
| Total | 63 | 62 | 38 | 15 | 244 | 309 | 731 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mejia, S.; Santos, J.L.B.; Noutsos, C. Comprehensive Genome-Wide Natural Variation and Expression Analysis of Tubby-like Proteins Gene Family in Brachypodium distachyon. Plants 2024, 13, 987. https://doi.org/10.3390/plants13070987
Mejia S, Santos JLB, Noutsos C. Comprehensive Genome-Wide Natural Variation and Expression Analysis of Tubby-like Proteins Gene Family in Brachypodium distachyon. Plants. 2024; 13(7):987. https://doi.org/10.3390/plants13070987
Chicago/Turabian StyleMejia, Sendi, Jose Lorenzo B. Santos, and Christos Noutsos. 2024. "Comprehensive Genome-Wide Natural Variation and Expression Analysis of Tubby-like Proteins Gene Family in Brachypodium distachyon" Plants 13, no. 7: 987. https://doi.org/10.3390/plants13070987