


Dissecting Genotype by Environment Interactions in Moroccan Wheat: An Advanced Biplot and Heatmap Analysis Unveiling Agronomic, Quality Traits, and Genotypic Stability for Tailored Breeding Strategies
Abstract
1. Introduction
2. Materials and Methods
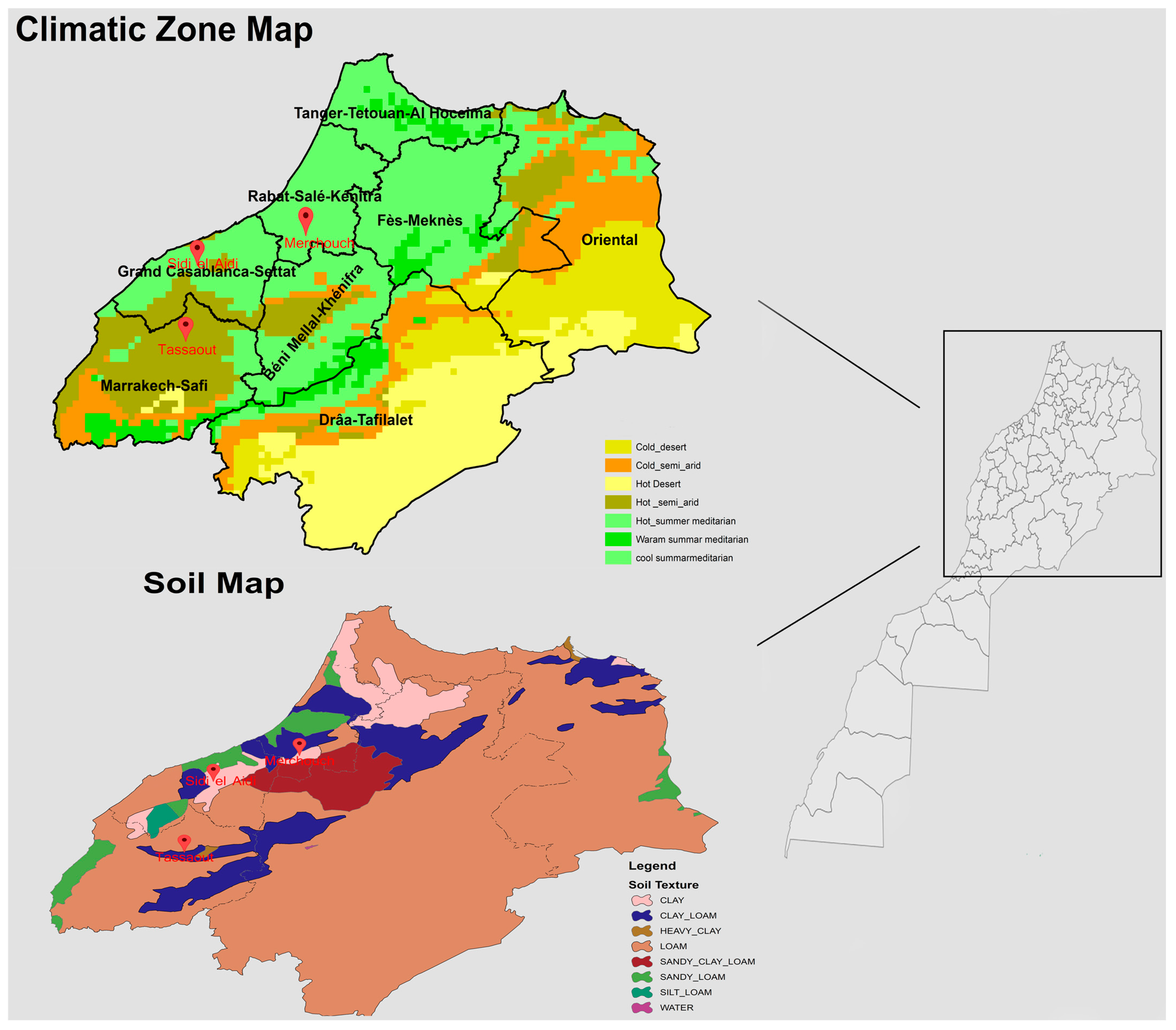
2.1. Research Locations and Climatic Variables
2.2. Experimental Framework
Characteristics of Experimental Plots and Agronomic Protocols
2.3. Data Collection and Trait Measurement
2.4. Statistical Analysis
3. Results
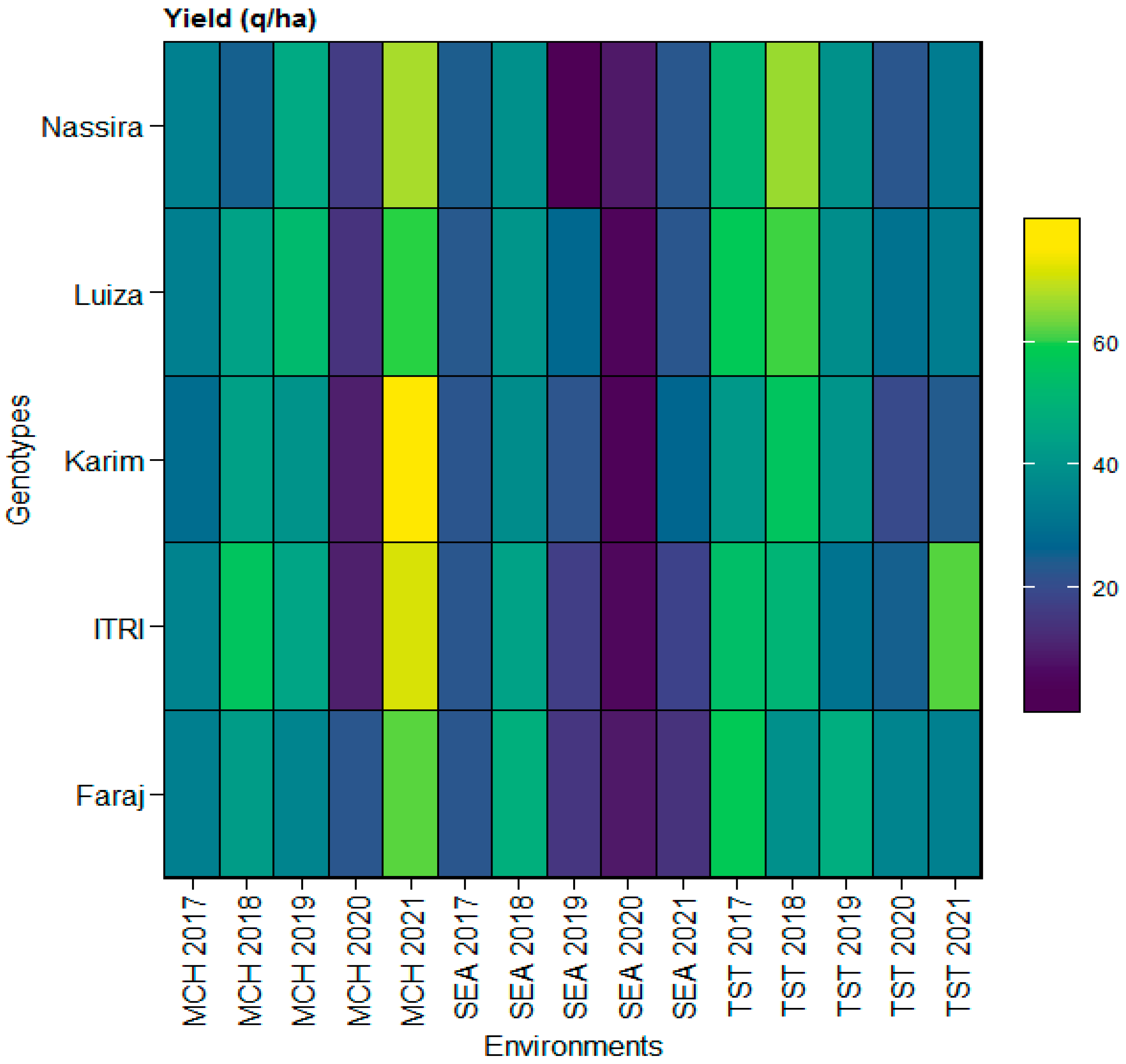
3.1. Descriptive Statistics for Yield and Yield Components across Multiple Locations
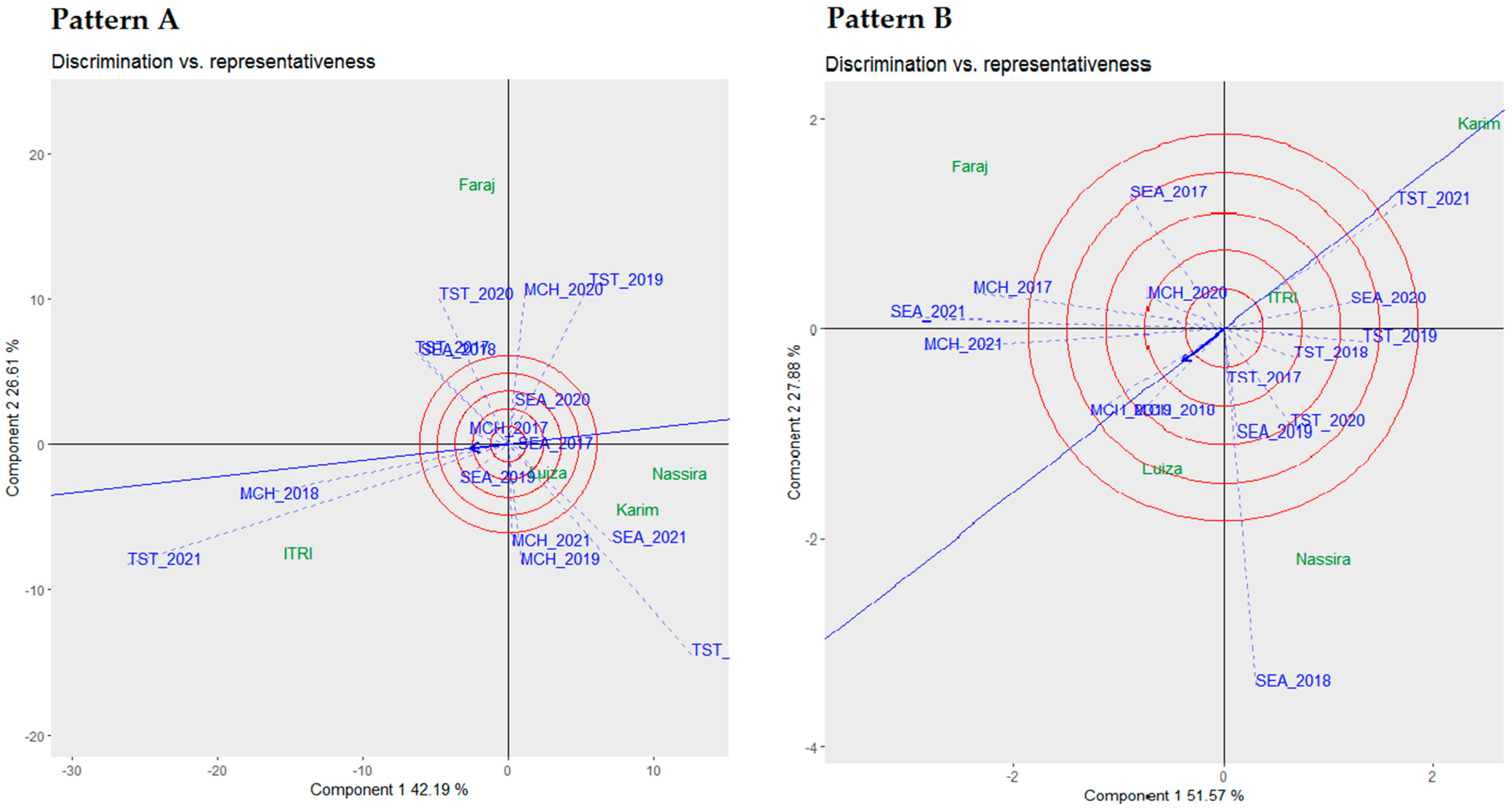
3.2. Discrimination vs. Representativeness Biplot Analysis
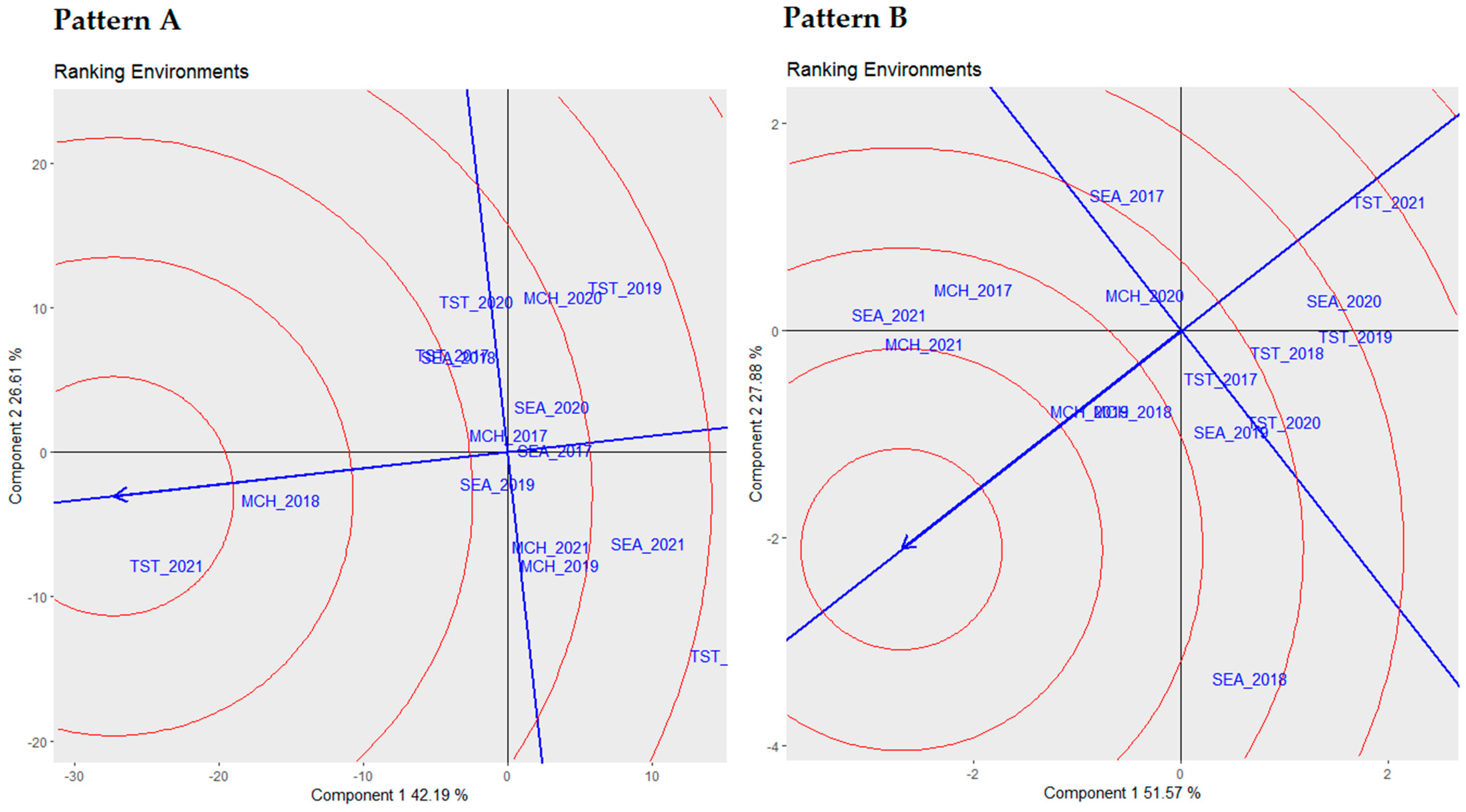
3.3. Ranking Environments via Biplot Analysis
3.4. Comprehensive Stability Analysis
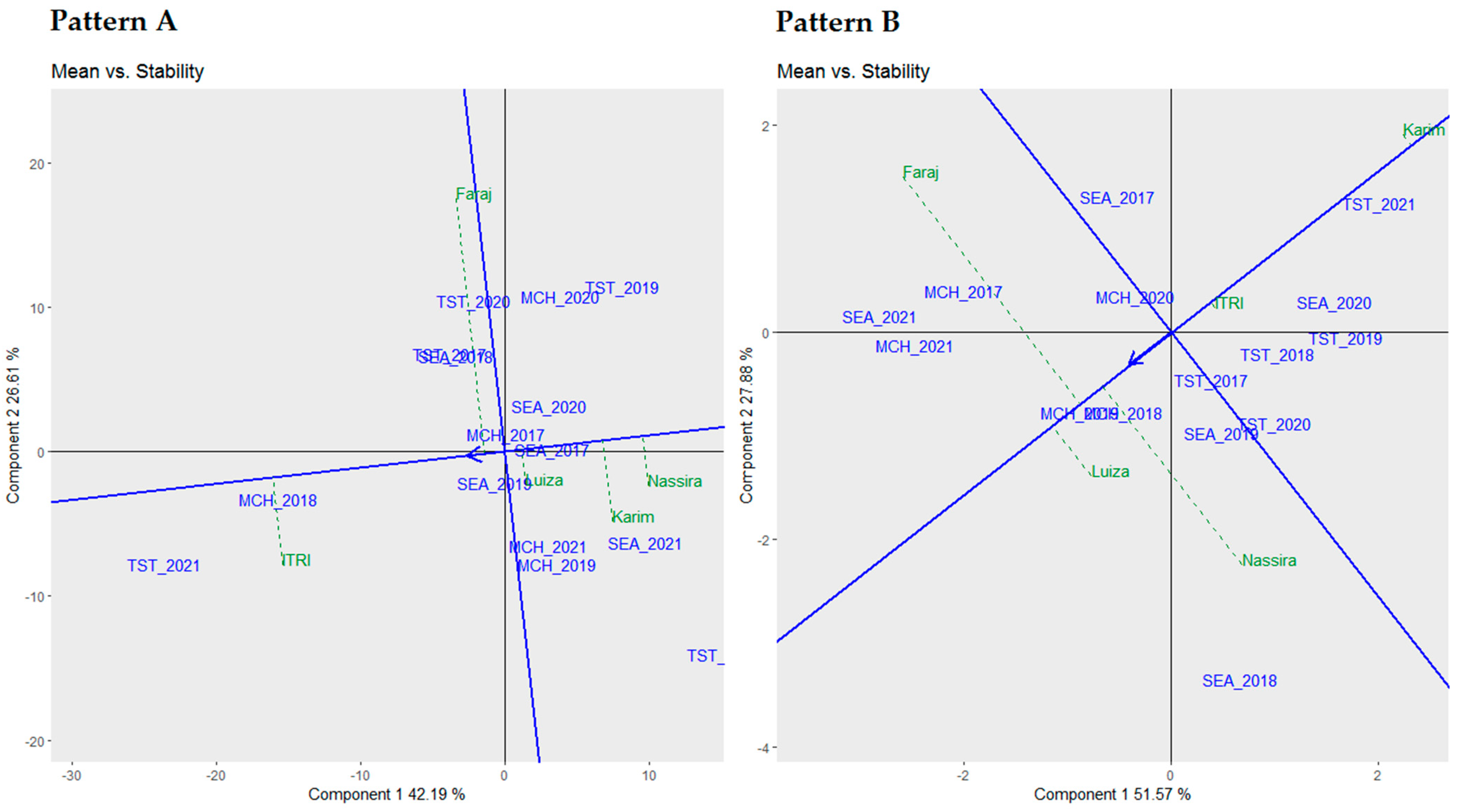
3.5. Mean vs. Stability Biplot Analysis
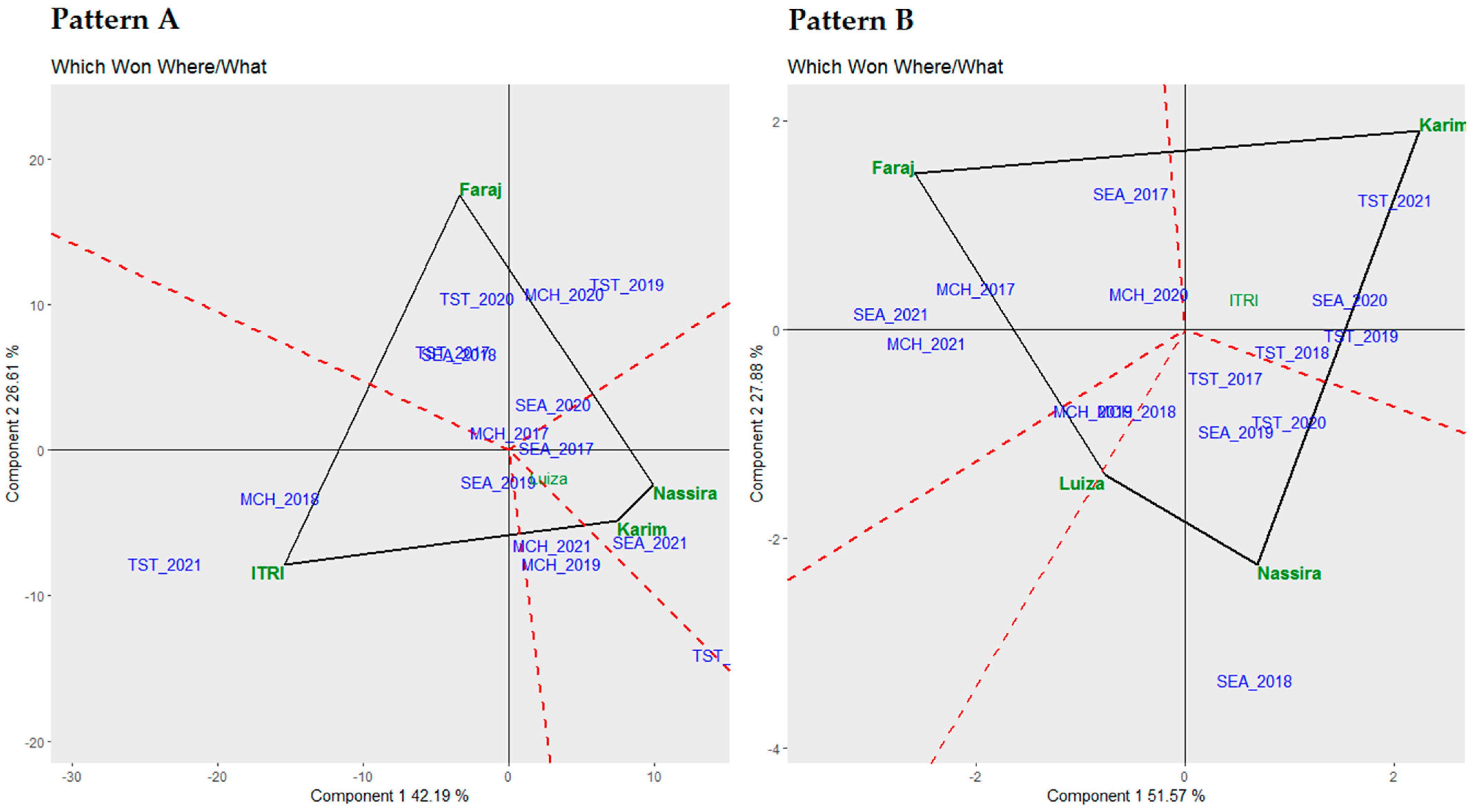
3.6. Which Won Where/What Biplot Analysis
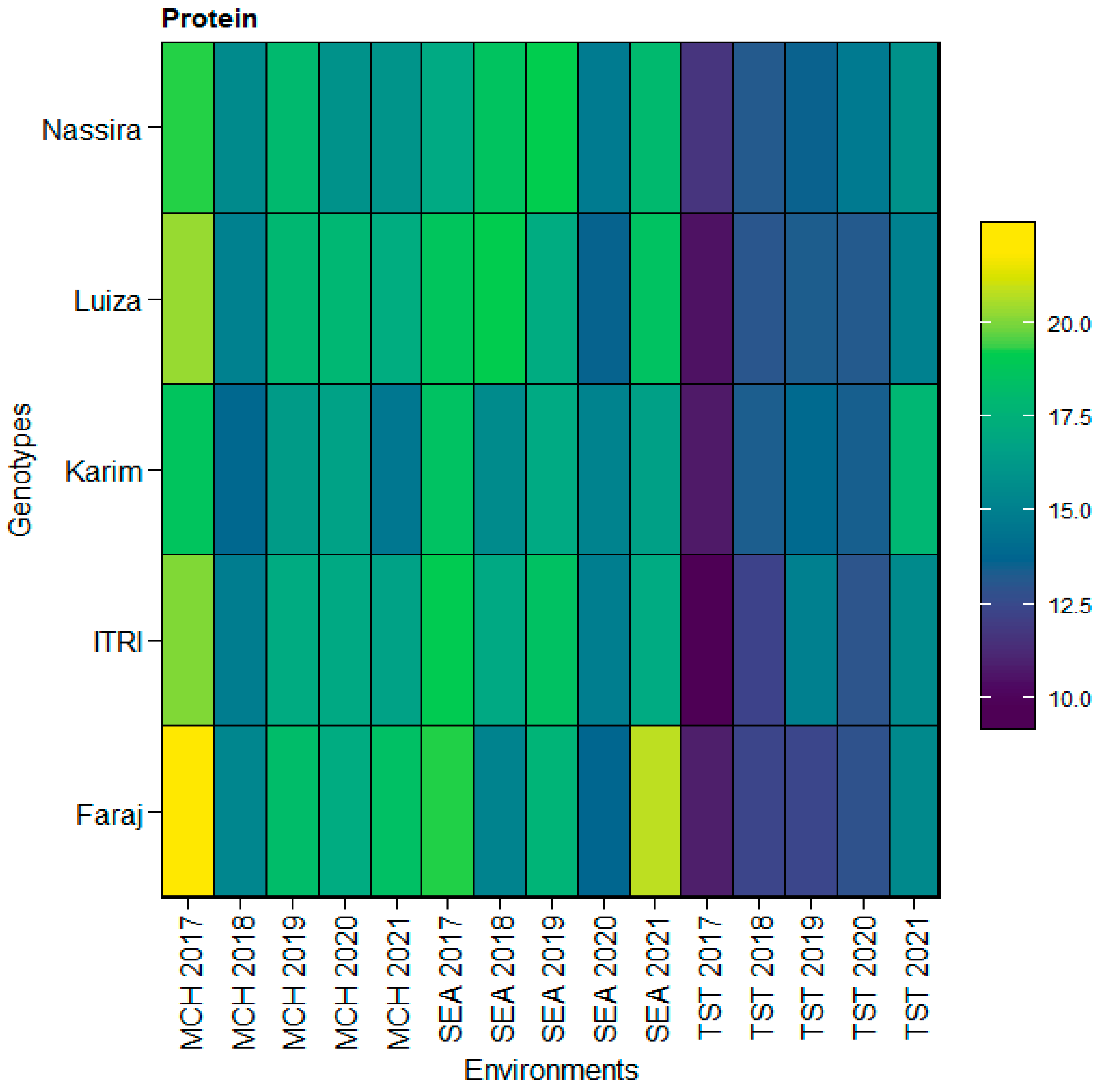
3.7. Descriptive Statistics for Quality Traits across Environments
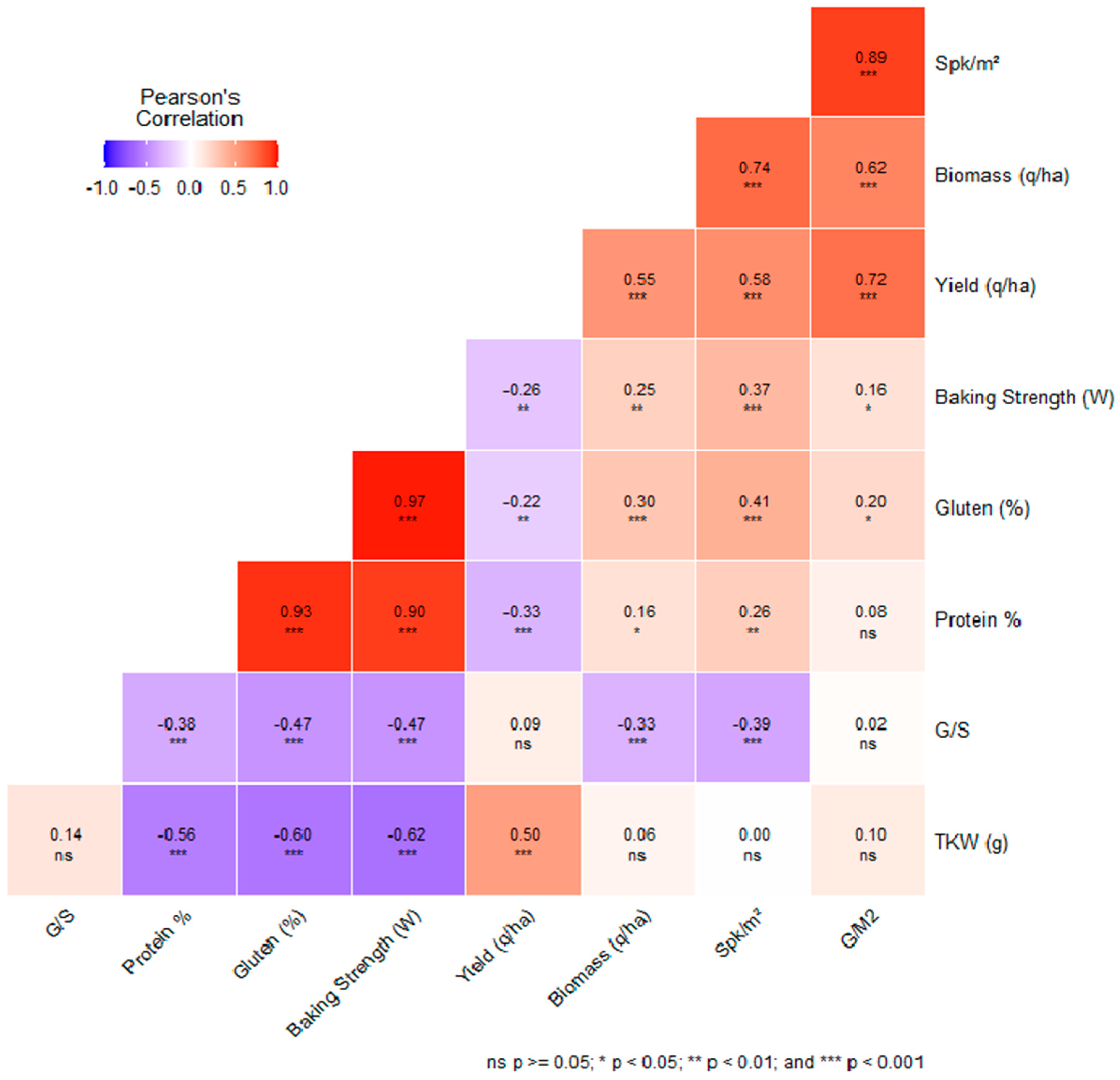
3.8. Correlations among Agronomic and Quality Traits in the Context of Genotype–Environment Interactions
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rossini, F.; Provenzano, M.E.; Sestili, F.; Ruggeri, R. Synergistic effect of sulfur and nitrogen in the organic and mineral fertilization of durum wheat: Grain yield and quality traits in the Mediterranean environment. Agronomy 2018, 8, 189. [Google Scholar] [CrossRef]
- Beres, B.L.; Rahmani, E.; Clarke, J.M.; Grassini, P.; Pozniak, C.J.; Geddes, C.M.; Porker, K.D.; May, W.E.; Ransom, J.K. A systematic review of durum wheat: Enhancing production systems by exploring genotype, environment, and management (G × E × M) synergies. Front. Plant Sci. 2020, 11, 568657. [Google Scholar] [CrossRef] [PubMed]
- Jaisi, S.; Thapa, A.; Poudel, M.R. Study of Correlation Coefficient and Path Analysis Among Yield Parameters of Wheat: A Review. INWASCON Technol. Mag. (I-TECH MAG) 2021, 3, 1–4. [Google Scholar] [CrossRef]
- Turuspekov, Y.; Baibulatova, A.; Yermekbayev, K.; Tokhetova, L.; Chudinov, V.; Sereda, G.; Ganal, M.; Griffiths, S.; Abugalieva, S. GWAS for plant growth stages and yield components in spring wheat (Triticum aestivum L.) harvested in three regions of Kazakhstan. BMC Plant Biol. 2017, 17, 190. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, R.; Armion, M.; Zadhasan, E.; Ahmadi, M.M.; Amri, A. The use of AMMI model for interpreting genotype × environment interaction in durum wheat. Exp. Agric. 2018, 54, 670–683. [Google Scholar] [CrossRef]
- Mohammadi, R.; Armion, M.; Sadeghzadeh, D.; Amri, A.; Nachit, M. Analysis of genotype-by-environment interaction for agronomic traits of durum wheat in Iran. Plant Prod. Sci. 2011, 14, 15–21. [Google Scholar] [CrossRef]
- Tanin, M.J.; Sharma, A.; Saini, D.K.; Singh, S.; Kashyap, L.; Srivastava, P.; Mavi, G.; Kaur, S.; Kumar, V.; Kumar, V. Ascertaining yield and grain protein content stability in wheat genotypes having the Gpc-B1 gene using univariate, multivariate, and correlation analysis. Front. Genet. 2022, 13, 1001904. [Google Scholar] [CrossRef] [PubMed]
- Kendal, E. Comparing durum wheat cultivars by genotype × yield × trait and genotype × trait biplot method. Chil. J. Agric. Res. 2019, 79, 512–522. [Google Scholar] [CrossRef]
- Slim, A.; Piarulli, L.; Chennaoui Kourda, H.; Rouaissi, M.; Robbana, C.; Chaabane, R.; Pignone, D.; Montemurro, C.; Mangini, G. Genetic structure analysis of a collection of Tunisian durum wheat germplasm. Int. J. Mol. Sci. 2019, 20, 3362. [Google Scholar] [CrossRef]
- Newton, I. Minitab Cookbook; Packt Publishing Ltd.: Birmingham, UK, 2014. [Google Scholar]
- Ryan, B.F.; Joiner, B.L.; Cryer, J.D. MINITAB Handbook: Update for Release; Cengage Learning: Boston, MA, USA, 2012. [Google Scholar]
- Francis, T.; Kannenberg, L. Yield stability studies in short-season maize. I. A descriptive method for grouping genotypes. Can. J. Plant Sci. 1978, 58, 1029–1034. [Google Scholar] [CrossRef]
- Wricke, G. Uber eine methode zur erfassung der okologischen streubreite in feldversucen. Z. Pflanzenzüchtung 1962, 47, 92–96. [Google Scholar]
- Shukla, G. Some statistical aspects of partitioning genotype-environmental components of variability. Heredity 1972, 29, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mi, J. Applying statistical methods to library data analysis. Ser. Libr. 2019, 76, 195–200. [Google Scholar] [CrossRef]
- Tonk, F.A.; Ilker, E.; Tosun, M. Evaluation of genotype x environment interactions in maize hybrids using GGE biplot analysis. Crop Breed. Appl. Biotechnol. 2011, 11, 1–9. [Google Scholar] [CrossRef]
- Lopez-Cruz, M.; Crossa, J.; Bonnett, D.; Dreisigacker, S.; Poland, J.; Jannink, J.-L.; Singh, R.P.; Autrique, E.; de los Campos, G. Increased prediction accuracy in wheat breeding trials using a marker × environment interaction genomic selection model. G3 Genes Genomes Genet. 2015, 5, 569–582. [Google Scholar] [CrossRef]
- El Haddad, N.; Sanchez-Garcia, M.; Visioni, A.; Jilal, A.; El Amil, R.; Sall, A.T.; Lagesse, W.; Kumar, S.; Bassi, F.M. Crop wild relatives crosses: Multi-location assessment in durum wheat, barley, and lentil. Agronomy 2021, 11, 2283. [Google Scholar] [CrossRef]
- Bassi, F.; Sanchez-Garcia, M. Adaptation and stability analysis of ICARDA durum wheat elites across 18 countries. Crop Sci. 2017, 57, 2419–2430. [Google Scholar] [CrossRef]
- Mulugeta, B.; Tesfaye, K.; Geleta, M.; Johansson, E.; Hailesilassie, T.; Hammenhag, C.; Hailu, F.; Ortiz, R. Multivariate analyses of Ethiopian durum wheat revealed stable and high yielding genotypes. PLoS ONE 2022, 17, e0273008. [Google Scholar] [CrossRef]
- Anastasi, U.; Corinzia, S.A.; Cosentino, S.L.; Scordia, D. Performances of durum wheat varieties under conventional and no-chemical input management systems in a semiarid Mediterranean environment. Agronomy 2019, 9, 788. [Google Scholar] [CrossRef]
- Jokar, F.; Karimizadeh, R.; Masoumiasl, A.; Fahliani, R.A. Canopy temperature and chlorophyll content are effective measures of drought stress tolerance in durum wheat. Not. Sci. Biol. 2018, 10, 575–583. [Google Scholar] [CrossRef]
- van der Bom, F.J.; Williams, A.; Raymond, N.S.; Alahmad, S.; Hickey, L.T.; Singh, V.; Bell, M.J. Root angle, phosphorus, and water: Interactions and effects on durum wheat genotype performance in drought-prone environments. Plant Soil 2023, 1–21. [Google Scholar] [CrossRef]
- Arjona, J.M.; Royo, C.; Dreisigacker, S.; Ammar, K.; Villegas, D. Effect of Ppd-A1 and Ppd-B1 allelic variants on grain number and thousand kernel weight of durum wheat and their impact on final grain yield. Front. Plant Sci. 2018, 9, 888. [Google Scholar] [CrossRef] [PubMed]
- Negisho, K.; Shibru, S.; Matros, A.; Pillen, K.; Ordon, F.; Wehner, G. Association mapping of drought tolerance indices in Ethiopian durum wheat (Triticum turgidum ssp. durum). Front. Plant Sci. 2022, 13, 838088. [Google Scholar] [CrossRef] [PubMed]
- Rabti, A.-b.; Mekaoussi, R.; Fellahi, Z.E.A.; Hannachi, A.; Benbelkacem, A.; Benmahammed, A.; Bouzerzour, H. Characterization of old and recent durum wheat [Triticum turgidum (L.) Tell. convar. durum (Desf.) Mackey] varieties assessed under South Mediterranean conditions. Egypt. J. Agron. 2020, 42, 307–320. [Google Scholar] [CrossRef]
- Eltaher, S.; Baenziger, P.S.; Belamkar, V.; Emara, H.A.; Nower, A.A.; Salem, K.F.; Alqudah, A.M.; Sallam, A. GWAS revealed effect of genotype × environment interactions for grain yield of Nebraska winter wheat. BMC Genom. 2021, 22, 2. [Google Scholar] [CrossRef] [PubMed]
- Farokhzadeh, S.; Shahsavand Hassani, H.; Mohammadi-Nejad, G.; Zinati, Z. Evaluation of grain yield stability of tritipyrum as a novel cereal in comparison with triticale lines and bread wheat varieties through univariate and multivariate parametric methods. PLoS ONE 2022, 17, e0274588. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, S.; Jarquin, D.; Crossa, J.; Reynolds, M. Genomic-enabled prediction accuracies increased by modeling genotype × environment interaction in durum wheat. Plant Genome 2018, 11, 170112. [Google Scholar] [CrossRef]
- Hacini, N.; Djelloul, R.; Desclaux, D. Study of genotype × environment interactions and agro-technological behavior of durum wheat varieties applied in different agro-climatic zones of algeria. Plant Arch. 2022, 22, 193–200. [Google Scholar] [CrossRef]
- Ayed, S.; Bouhaouel, I.; Othmani, A.; Bassi, F.M. Use of wild relatives in durum wheat (Triticum turgidum L. var. durum Desf.) breeding program: Adaptation and stability in context of contrasting environments in Tunisia. Agronomy 2021, 11, 1782. [Google Scholar] [CrossRef]
- Megahed, E.M.; Awaad, H.A.; Ramadan, I.E.; Abdul-Hamid, M.I.; Sweelam, A.A.; El-Naggar, D.R.; Mansour, E. Assessing performance and stability of yellow rust resistance, heat tolerance, and agronomic performance in diverse bread wheat genotypes for enhancing resilience to climate change under Egyptian conditions. Front. Plant Sci. 2022, 13, 1014824. [Google Scholar] [CrossRef]
- Habash, D.Z.; Baudo, M.; Hindle, M.; Powers, S.J.; Defoin-Platel, M.; Mitchell, R.; Saqi, M.; Rawlings, C.; Latiri, K.; Araus, J.L. Systems responses to progressive water stress in durum wheat. PLoS ONE 2014, 9, e108431. [Google Scholar] [CrossRef] [PubMed]
- Tomić, J.M.; Torbica, A.M.; Popović, L.M.; Rakita, S.M.; Živančev, D.R. Breadmaking potential and proteolytic activity of wheat varieties from two production years with different climate conditions. Food Feed Res. 2015, 42, 83–90. [Google Scholar] [CrossRef]
- Gagliardi, A.; Carucci, F.; Masci, S.; Flagella, Z.; Gatta, G.; Giuliani, M.M. Effects of genotype, growing season and nitrogen level on gluten protein assembly of durum wheat grown under mediterranean conditions. Agronomy 2020, 10, 755. [Google Scholar] [CrossRef]
- Niu, N.; Arief, V.N.; DeLacy, I.H.; Lush, D.; Sheppard, J.; Zhang, G.; Dieters, M.J. Genetic gain in yield and protein over two cycles of a wheat recurrent selection program. Breed. Sci. 2010, 60, 181–186. [Google Scholar] [CrossRef][Green Version]
- Kebrom, T.H.; Chandler, P.M.; Swain, S.M.; King, R.W.; Richards, R.A.; Spielmeyer, W. Inhibition of tiller bud outgrowth in the tin mutant of wheat is associated with precocious internode development. Plant Physiol. 2012, 160, 308–318. [Google Scholar] [CrossRef]
- Vicente, R.; Pérez, P.; Martínez-Carrasco, R.; Feil, R.; Lunn, J.E.; Watanabe, M.; Arrivault, S.; Stitt, M.; Hoefgen, R.; Morcuende, R. Metabolic and transcriptional analysis of durum wheat responses to elevated CO2 at low and high nitrate supply. Plant Cell Physiol. 2016, 57, 2133–2146. [Google Scholar] [CrossRef]
- Mohammadi, R.; Sadeghzadeh, B.; Ahmadi, M.M. Evaluation of genotype × environment interaction in durum wheat (Triticum turgidum var. durum L.) regional yield trials. Iran. J. Crop Sci. 2020, 22, 15–31. [Google Scholar] [CrossRef]
- Sakin, M.A.; Akinci, C.; Duzdemir, O.; Donmez, E. Assessment of genotype x environment interaction on yield and yield components of durum wheat genotypes by multivariate analyses. Afr. J. Biotechnol. 2011, 10, 2875–2885. [Google Scholar]
- Mohamed, M.M.; Darwish, M.; El-Rady, A.; Ghalab, E.; Elfanah, A. Estimation of AMMI and GGE biplots for some bread and durum wheat genotypes. J. Plant Prod. 2022, 13, 75–83. [Google Scholar] [CrossRef]
- Verma, A.; Chatrath, R.; Singh, G. Adaptability of wheat genotypes under multi-environment trials for Northern Hills Zone. Int. J. Bio-Resour. Stress Manag. 2020, 11, 304–310. [Google Scholar] [CrossRef]
- Kamara, M.M.; Rehan, M.; Mohamed, A.M.; El Mantawy, R.F.; Kheir, A.M.; Abd El-Moneim, D.; Safhi, F.A.; ALshamrani, S.M.; Hafez, E.M.; Behiry, S.I. Genetic potential and inheritance patterns of physiological, agronomic and quality traits in bread wheat under normal and water deficit conditions. Plants 2022, 11, 952. [Google Scholar] [CrossRef] [PubMed]
- Ionut, R.; Hiriscau, D.; Kadar, R.; Berindean, I. Phenotypic response of spring wheat perspective line genotypes to different environmental conditions. Life Sci. Sustain. Dev. 2022, 3, 81–86. [Google Scholar]
- Roostaei, M.; Jafarzadeh, J.; Roohi, E.; Nazary, H.; Rajabi, R.; Mohammadi, R.; Khalilzadeh, G.R.; Seif, F.; Mirfatah, S.M.M.; Amiri, S.S. Genotype × environment interaction and stability analyses of grain yield in rainfed winter bread wheat. Exp. Agric. 2022, 58, e37. [Google Scholar] [CrossRef]
- Chairi, F.; Aparicio, N.; Serret, M.D.; Araus, J.L. Breeding effects on the genotype × environment interaction for yield of durum wheat grown after the Green Revolution: The case of Spain. Crop J. 2020, 8, 623–634. [Google Scholar] [CrossRef]
- Dabi, A.; Alemu, G.; Sime, B.; Geleta, N.; Delessa, A.; Solomon, T.; Zegaye, H.; Asnake, D.; Asefa, B.; Duga, R. Genotype × Environment Interaction and Stability Analysis Using GGE Biplot for Grain Yield of Bread Wheat (Triticum aestivum) Genotypes under Low Moisture Stress Areas of Ethiopia. Int. J. Bio-Resour. Stress Manag. 2023, 14, 1089–1098. [Google Scholar] [CrossRef]
- Lin, C.-S.; Binns, M.R. A superiority measure of cultivar performance for cultivar × location data. Can. J. Plant Sci. 1988, 68, 193–198. [Google Scholar] [CrossRef]
- Martínez-Peña, R.; Schlereth, A.; Höhne, M.; Encke, B.; Morcuende, R.; Nieto-Taladriz, M.T.; Araus, J.L.; Aparicio, N.; Vicente, R. Source-sink dynamics in field-grown durum wheat under contrasting nitrogen supplies: Key role of non-foliar organs during grain filling. Front. Plant Sci. 2022, 13, 869680. [Google Scholar] [CrossRef] [PubMed]
- Al-Sayaydeh, R.; Shtaya, M.; Qubbaj, T.; Al-Rifaee, M.; Alabdallah, M.; Migdadi, O.; Gammoh, I.; Al-Abdallat, A. Performance and Stability Analysis of Selected Durum Wheat Genotypes Differing in Their Kernel Characteristics. Plants 2023, 12, 2664. [Google Scholar] [CrossRef]
- Johnson, M.; Kumar, A.; Oladzad-Abbasabadi, A.; Salsman, E.; Aoun, M.; Manthey, F.A.; Elias, E.M. Association mapping for 24 traits related to protein content, gluten strength, color, cooking, and milling quality using balanced and unbalanced data in durum wheat [Triticum turgidum L. var. durum (Desf).]. Front. Genet. 2019, 10, 717. [Google Scholar] [CrossRef]
- Plavšin, I.; Gunjača, J.; Šimek, R.; Novoselović, D. Capturing GEI patterns for quality traits in biparental wheat populations. Agronomy 2021, 11, 1022. [Google Scholar] [CrossRef]
- Vida, G.; Cséplő, M.; Rakszegi, M.; Bányai, J. Effect of multi-year environmental and meteorological factors on the quality traits of winter durum wheat. Plants 2021, 11, 113. [Google Scholar] [CrossRef] [PubMed]
- Pacuta, V.; Rasovsky, M.; Michalska-Klimczak, B.; Wyszynski, Z. Grain yield and quality traits of durum wheat (Triticum durum Desf.) treated with seaweed-and humic acid-based biostimulants. Agronomy 2021, 11, 1270. [Google Scholar] [CrossRef]
- Kyratzis, A.C.; Pallides, A.; Katsiotis, A. Investigating stability parameters for agronomic and quality traits of durum wheat grown under Mediterranean conditions. Agronomy 2022, 12, 1774. [Google Scholar] [CrossRef]
- Bnejdi, F.; Gazzah, M.E. Epistasis and genotype-by-environment interaction of grain protein content in durum wheat. Genet. Mol. Biol. 2010, 33, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ma, X.; Li, Y.; Ju, H. Geographical detector-based wheat quality attribution under genotype, environment, and crop management frameworks. Front. Environ. Sci. 2022, 10, 1037979. [Google Scholar] [CrossRef]
- Nigro, D.; Fortunato, S.; Giove, S.L.; Paradiso, A.; Gu, Y.Q.; Blanco, A.; De Pinto, M.C.; Gadaleta, A. Glutamine synthetase in durum wheat: Genotypic variation and relationship with grain protein content. Front. Plant Sci. 2016, 7, 971. [Google Scholar] [CrossRef] [PubMed]
- Muqaddasi, Q.H.; Brassac, J.; Ebmeyer, E.; Kollers, S.; Korzun, V.; Argillier, O.; Stiewe, G.; Plieske, J.; Ganal, M.W.; Röder, M.S. Prospects of GWAS and predictive breeding for European winter wheat’s grain protein content, grain starch content, and grain hardness. Sci. Rep. 2020, 10, 12541. [Google Scholar] [CrossRef]
- Ruan, Y.; Yu, B.; Knox, R.E.; Singh, A.K.; DePauw, R.; Cuthbert, R.; Zhang, W.; Piche, I.; Gao, P.; Sharpe, A. High density mapping of quantitative trait loci conferring gluten strength in Canadian durum wheat. Front. Plant Sci. 2020, 11, 170. [Google Scholar] [CrossRef]
- Hao, S.; Lou, H.; Wang, H.; Shi, J.; Liu, D.; Tao, J.; Miao, S.; Pei, Q.; Yu, L.; Wu, M. Genome-wide association study reveals the genetic basis of five quality traits in Chinese wheat. Front. Plant Sci. 2022, 13, 835306. [Google Scholar] [CrossRef]
- Nazco, R.; Peña, R.J.; Ammar, K.; Villegas, D.; Crossa, J.; Royo, C. Durum wheat (Triticum durum Desf.) Mediterranean landraces as sources of variability for allelic combinations at Glu-1/Glu-3 loci affecting gluten strength and pasta cooking quality. Genet. Resour. Crop Evol. 2014, 61, 1219–1236. [Google Scholar] [CrossRef]
- Kirouani, A.; Taghouti, M.; Boukhalfoun, L.; Henkrar, F.; Udupa, S. Bread making possibility of Algerian durum wheat (Triticum turgidum L. var. Durum) varieties. J. Fundam. Appl. Sci. 2020, 12, 1223–1234. [Google Scholar] [CrossRef]
- Al-Khayri, J.M.; Alshegaihi, R.M.; Mahgoub, E.I.; Mansour, E.; Atallah, O.O.; Sattar, M.N.; Al-Mssallem, M.Q.; Alessa, F.M.; Aldaej, M.I.; Hassanin, A.A. Association of High and Low Molecular Weight Glutenin Subunits with Gluten Strength in Tetraploid Durum Wheat (Triticum turgidum spp. Durum L.). Plants 2023, 12, 1416. [Google Scholar] [CrossRef] [PubMed]
- Huertas-García, A.B.; Guzmán, C.; Ibba, M.I.; Rakszegi, M.; Sillero, J.C.; Alvarez, J.B. Processing and bread-making quality profile of spanish spelt wheat. Foods 2023, 12, 2996. [Google Scholar] [CrossRef]
- Maich, R.; Di Rienzo, J. Genotype × tillage interaction in a recurrent selection program in wheat. Cereal Res. Commun. 2014, 42, 525–533. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Station | Soil Type | Precipitations (mm) for the Five Growing Seasons | Geographical Location | Altitude (Meters) | |||||
|---|---|---|---|---|---|---|---|---|---|
| 2016–2017 | 2017–2018 | 2018–2019 | 2019–2020 | 2020–2021 | Latitude | Longitude | |||
| Sidi El Aidi (SEA) | Vertisol | 290 | 505 | 210 | 242 | 467 | 33.12218° N | 7.63315° W | 235 |
| Merchouch (MCH) | Cambisol | 348 | 579 | 179 | 249 | 518 | 33.60499° N | 6.71000° W | 399 |
| Tassaout (TST) | Alfisol | 216 | 305 | 200 | 247 | 304 | 31.82021° N | 7.43806° W | 591 |
| Cultivar | Code | Year of Registration | Pedigree and Breeding History |
|---|---|---|---|
| Faraj | V1 | 2007 | Hybrid Nassira, Qarmal, Lahn (ICARDA) |
| Itri | V2 | 2017 | RISSA/GAN//POHO_1/3/PLATA_3//CREX/ALLA/x Karim |
| Karim | V3 | 1985 | Bittern ‘S’ « JO’S’. AA”:S’//FG’S’ » |
| Luiza | V4 | 2011 | RASCON_39/TILO_1 |
| Nassira | V5 | 2003 | INRA Selection on CIMMYT EII, 12 TA14/BD3//Isly # CF41530–1548 |
| Env (E): (Combinations of Location * Season) | Gen (G) | YLD (q/ha) | Biomass (q/ha) | TKW (g) | Spk/m2 | G/S | G/m2 |
|---|---|---|---|---|---|---|---|
| SEA 2017 | V1 | 23.01 abc | 105 a | 37.3 bcdef | 324 ab | 25 a | 8300 ab |
| V2 | 23.21 abc | 77.5 a | 33.45 def | 284 abcd | 26 a | 7347 ab | |
| V3 | 22.93 abc | 78.33 a | 43.1 abcde | 219 abcdef | 29 a | 6431 ab | |
| V4 | 23.66 abc | 104.17 a | 33.9 def | 297 abc | 35 a | 9715 a | |
| V5 | 24.65 abc | 111.67 a | 47.45 abc | 302 ab | 25 a | 7622 ab | |
| Mean | 23.49 | 95.33 | 39.04 | 284 | 28 | 7883 | |
| SEA 2018 | V1 | 48.82 a | 120.83 a | 49.13 ab | 284 abcd | 28 a | 8092 ab |
| V2 | 44.78 ab | 106.67 a | 47.13 abc | 373 a | 21 a | 7690 ab | |
| V3 | 38.28 abc | 103.33 a | 50.5 a | 213 abcdef | 29 a | 6143 ab | |
| V4 | 40.95 abc | 122.5 a | 47 abc | 316 ab | 16 a | 5071 ab | |
| V5 | 39.33 abc | 96.67 a | 45.88 abcd | 273 abcd | 14 a | 3677 ab | |
| Mean | 42.43 | 110 | 47.93 | 289 | 22 | 6135 | |
| SEA 2019 | V1 | 15 abc | 87.5 a | 35.4 cdef | 203 abcdefg | 28 a | 5549 ab |
| V2 | 16.73 abc | 94.17 a | 31.4 ef | 270 abcd | 27 a | 6596 ab | |
| V3 | 22.75 abc | 97.5 a | 33 ef | 273 abcd | 26 a | 7155 ab | |
| V4 | 27.78 abc | 105.83 a | 38.6 abcdef | 278 abcd | 26 a | 6869 ab | |
| V5 | 3.32 c | 39.17 a | 34.49 def | 113 defg | 22 a | 2319 ab | |
| Mean | 17.12 | 84.83 | 34.58 | 227 | 26 | 5698 | |
| SEA 2020 | V1 | 8.58 abc | 95.37 a | 32.55 ef | 73 efg | 27 a | 1982 b |
| V2 | 5.49 bc | 82.3 a | 30.45 f | 49 efg | 25 a | 1204 b | |
| V3 | 4.22 bc | 97.85 a | 31.35 ef | 32 g | 27 a | 842 b | |
| V4 | 4.72 bc | 105.64 a | 30.25 f | 41 fg | 25 a | 1034 b | |
| V5 | 8.51 abc | 82.7 a | 31.85 ef | 73 efg | 21 a | 1544 b | |
| Mean | 6 | 93 | 31 | 54 | 25 | 1321 | |
| SEA 2021 | V1 | 14.07 abc | 68.15 a | 31.2 ef | 111 defg | 32 a | 3289 ab |
| V2 | 17.76 abc | 50.85 a | 32.6 ef | 122 cdefg | 33 a | 3966 ab | |
| V3 | 27.2 abc | 112.22 a | 40 abcdef | 224 abcde | 19 a | 5295 ab | |
| V4 | 23.09 abc | 90.07 a | 35.6 cdef | 170 bcdefg | 28 a | 4717 ab | |
| V5 | 23.26 abc | 83.3 a | 35.6 cdef | 184 bcdefg | 26 a | 4776 ab | |
| Mean | 21 | 81 | 35 | 162 | 28 | 4409 | |
| Analysis of variance | |||||||
| Gen (G) | ns | ns | * | ns | ns | ns | |
| Env (E) | *** | ns | *** | *** | ns | *** | |
| Gen (G) * Env (E) | ns | ns | * | ** | ns | ns | |
| Env (E): (Combinations of Location * Season) | Gen (G) | YLD (q/ha) | Biomass (q/ha) | TKW (g) | Spk/m2 | G/S | G/m2 |
|---|---|---|---|---|---|---|---|
| MCH 2017 | V1 | 34.3 bcdef | 247.5 ab | 34.9 abcd | 818 abcde | 12 d | 10,006 bcdefg |
| V2 | 35.46 bcdef | 229.5 ab | 33.35 abcd | 726 bcdefgh | 20 cd | 13,740 bcdef | |
| V3 | 29.17 cdef | 216 ab | 39.05 abcd | 624 cdefghi | 16 d | 9785 bcdefg | |
| V4 | 34.25 bcdef | 236.25 ab | 37.15 abcd | 726 bcdefgh | 19 cd | 13,794 bcdef | |
| V5 | 34.64 bcdef | 258 a | 29.75 d | 875 abc | 19 cd | 16,262 abcd | |
| Mean | 33.56 | 237.45 | 34.84 | 751 | 17 | 12,717 | |
| MCH 2018 | V1 | 42.97 abcdef | 163.33 abcde | 46.75 ab | 240 hi | 34 a | 8046 cdefg |
| V2 | 56.6 abcd | 166.67 abcde | 45.75 abc | 281 fghi | 29 abc | 8168 cdefg | |
| V3 | 44 abcdef | 148.33 abcde | 38.88 abcd | 278 fghi | 32 ab | 8710 cdefg | |
| V4 | 44.47 abcdef | 137.5 abcde | 46.88 a | 281 fghi | 28 abc | 7698 cdefg | |
| V5 | 25.73 cdef | 143.33 abcde | 44.38 abcd | 265 ghi | 19 cd | 5068 fg | |
| Mean | 43 | 152 | 45 | 267 | 28 | 7538 | |
| MCH 2019 | V1 | 35.81 bcdef | 154.17 abcde | 39 abcd | 338 defghi | 12 d | 4217 fg |
| V2 | 45.58 abcdef | 258.33 a | 39.15 abcd | 429 cdefghi | 13 d | 5449 efg | |
| V3 | 40.47 abcdef | 208.33 abc | 37.8 abcd | 394 cdefghi | 13 d | 5108 efg | |
| V4 | 53.1 abcde | 260 a | 37.45 abcd | 527 cdefghi | 14 d | 7117 defg | |
| V5 | 46.85 abcdef | 210 abc | 40.95 abcd | 410 cdefghi | 20 cd | 8300 cdefg | |
| Mean | 44 | 218 | 39 | 419 | 14 | 6038 | |
| MCH 2020 | V1 | 23.1 def | 129.44 abcde | 39.7 abcd | 327 efghi | 17 cd | 5505 efg |
| V2 | 9.83 f | 38.31 e | 31.3 cd | 100 i | 22 abcd | 2395 g | |
| V3 | 9.62 f | 47.1 de | 30.45 d | 116 i | 21 bcd | 2406 g | |
| V4 | 14.22 f | 65.89 cde | 31.85 bcd | 178 i | 21 bcd | 4096 g | |
| V5 | 16.22 ef | 108.38 bcde | 37.35 abcd | 308 efghi | 19 cd | 5729 efg | |
| Mean | 14.6 | 77.83 | 34.13 | 205 | 20 | 4026 | |
| MCH 2021 | V1 | 62.98 abc | 166.82 abcde | 39.15 abcd | 859 abcd | 17 cd | 14,693 abcde |
| V2 | 71.82 ab | 154.19 abcde | 35.1 abcd | 783 abcdefg | 22 abcd | 17,099 abc | |
| V3 | 76.03 a | 190.06 abcd | 44.4 abcd | 940 ab | 21 bcd | 19,270 ab | |
| V4 | 61.48 abc | 146.81 abcde | 38.55 abcd | 788 abcdef | 21 bcd | 16,554 abcd | |
| V5 | 67.7 ab | 228.1 ab | 41.75 abcd | 1293 a | 19 cd | 23,895 a | |
| Mean | 68 | 177.2 | 39.79 | 932 | 20 | 18,302 | |
| Analysis of Variance | |||||||
| Gen (G) | ns | ns | ns | ns | ns | * | |
| Env (E) | *** | *** | *** | *** | *** | *** | |
| Gen (G) * Env (E) | ns | ns | ns | ns | ** | ns | |
| Env (E): (Combinations of Location * Season) | Gen (G) | YLD (q/ha) | Biomass (q/ha) | TKW (g) | Spk/m2 | G/S | G/m2 |
|---|---|---|---|---|---|---|---|
| TST 2017 | V1 | 58.45 ab | 136.5 a | 58.35 a | 302 abc | 27 a | 8400 ab |
| V2 | 54.58 ab | 113.25 abcd | 46.1 abcde | 235 abc | 36 a | 8370 ab | |
| V3 | 41.93 ab | 79.5 abcd | 53 ab | 192 abc | 31 a | 5770 ab | |
| V4 | 58.22 ab | 85.5 abcd | 50 abcd | 197 abc | 34 a | 6488 ab | |
| V5 | 51.97 ab | 111.75 bcd | 51.4 abc | 238 abc | 31 a | 7320 ab | |
| Mean | 53.03 | 105.3 | 51.77 | 232 | 32 | 7270 | |
| TST 2018 | V1 | 39.37 ab | 191.67 a | 47.75 abcde | 402 a | 17 a | 6661 ab |
| V2 | 50.73 ab | 83.33 abcd | 43.5 bcde | 362 ab | 26 a | 9447 ab | |
| V3 | 56.62 ab | 152.5 abcd | 49.25 abcde | 383 a | 24 a | 9315 ab | |
| V4 | 62 ab | 162.5 abcd | 47.38 abcde | 356 ab | 30 a | 10,562 ab | |
| V5 | 66.57 a | 114.17 bcd | 49.25 abcde | 311 abc | 27 a | 8659 ab | |
| Mean | 55.06 | 140.83 | 47.43 | 362 | 25 | 8929 | |
| TST 2019 | V1 | 48.34 ab | 155 ab | 41.25 bcde | 373 ab | 23 a | 7803 ab |
| V2 | 30.79 ab | 107.5 abcd | 37.15 e | 167 bc | 33 a | 5513 ab | |
| V3 | 40.54 ab | 134.17 abcd | 45.5 bcde | 265 abc | 26 a | 6774 ab | |
| V4 | 38.66 ab | 116.67 bcd | 38.35 de | 208 abc | 28 a | 5873 ab | |
| V5 | 39.62 ab | 86.67 cd | 44.6 bcde | 208 abc | 27 a | 5589 ab | |
| Mean | 39.59 | 120 | 41.37 | 243 | 27 | 6310 | |
| TST 2020 | V1 | 35.8 ab | 123.49 abc | 50.75 abcd | 257 abc | 23 a | 5889 ab |
| V2 | 25.65 ab | 59.75 abcd | 45.65 bcde | 135 c | 33 a | 4528 ab | |
| V3 | 19.87 b | 60.73 abcd | 48.75 abcde | 130 c | 26 a | 3391 b | |
| V4 | 30.5 ab | 76.42 bcd | 49.85 abcd | 165 bc | 28 a | 4620 ab | |
| V5 | 23.08 ab | 54.54 cd | 51.8 abc | 122 c | 27 a | 3299 b | |
| Mean | 26.98 | 74.99 | 49.36 | 159 | 27 | 4345 | |
| TST 2021 | V1 | 34.62 ab | 130 abcd | 45.6 bcde | 254 abc | 26 a | 6661 ab |
| V2 | 62.77 ab | 194.31 abcd | 46.45 abcde | 289 abc | 41 a | 11,821 a | |
| V3 | 24.15 ab | 144.92 abcd | 40.45 cde | 224 abc | 25 a | 5311 ab | |
| V4 | 33.69 ab | 127.38 bcd | 47.9 abcde | 259 abc | 22 a | 5967 ab | |
| V5 | 33.23 ab | 134.31 d | 47.4 abcde | 262 abc | 24 a | 6261 ab | |
| Mean | 37.69 | 146.18 | 45.56 | 257 | 28 | 7204 | |
| Analysis of variance | |||||||
| Gen (G) | ns | ** | ** | ** | ** | ns | |
| Env (E) | *** | *** | *** | *** | ns | *** | |
| Gen (G) * Env (E) | ns | * | ns | ns | ns | ns | |
| Genotype | Francis Cumulative Values | GR (Francis) | Wricke’s Ecovalence (W) | GR (Wricke) | Shukla’s Stability Variance (σ2) | GR (Shukla) |
|---|---|---|---|---|---|---|
| Faraj (V1) | 48 | 3 | 3,607,727 | 1 | 457 | 2 |
| ITRI (V2) | 48 | 3 | 5,763,770 | 5 | 580 | 5 |
| Karim (V3) | 34 | 1 | 4,630,654 | 3 | 400 | 1 |
| Luiza (V4) | 49 | 5 | 4,084,894 | 2 | 333 | 3 |
| Nassira (V5) | 46 | 2 | 5,041,188 | 4 | 602 | 4 |
| Env (E): (Combinations of Location * Season) | Gen (G) | Protein (%) | Gluten (%) | Baking Strength (W) |
|---|---|---|---|---|
| SEA 2017 | V1 | 19.46 ab | 49.63 a | 494.29 a |
| V2 | 19.17 ab | 49.9 a | 466.61 ab | |
| V3 | 18.6 abcd | 48.39 ab | 454.14 ab | |
| V4 | 18.8 abc | 48.3 abc | 451.57 abc | |
| V5 | 17.12 bcdef | 42.03 abcdef | 378.54 bcdefgh | |
| Mean | 18.63 | 47.65 | 449.03 | |
| SEA 2018 | V1 | 15.18 def | 34.06 fg | 324.94 gh |
| V2 | 17.06 bcdef | 41.29 abcdefg | 370.62 bcdefgh | |
| V3 | 15.62 cdef | 36.56 defg | 310.27 h | |
| V4 | 19.24 ab | 48.44 ab | 453.43 ab | |
| V5 | 18.69 abcd | 46.16 abcd | 426.9 abcdef | |
| Mean | 17.16 | 41.3 | 377.23 | |
| SEA 2019 | V1 | 17.73 abcde | 40.06 abcdefg | 392.15 bcdefgh |
| V2 | 18.57 abcd | 47.53 abc | 436.36 abcd | |
| V3 | 17.12 bcdef | 42.9 abcdef | 385.59 bcdefgh | |
| V4 | 17.23 bcdef | 44.11 abcdef | 412.67 abcdefg | |
| V5 | 19.26 ab | 47.32 abc | 432.03 abcde | |
| Mean | 17.98 | 44.38 | 411.76 | |
| SEA 2020 | V1 | 13.8 f | 31.26 g | 306.14 h |
| V2 | 14.95 ef | 38.24 bcdefg | 351.07 cdefgh | |
| V3 | 15.2 def | 38.13 cdefg | 343.56 defgh | |
| V4 | 13.7 f | 35.07 efg | 328.13 fgh | |
| V5 | 14.8 ef | 36.37 defg | 332.01 efgh | |
| Mean | 14 | 36 | 332 | |
| SEA 2021 | V1 | 20.9 a | 47.19 abc | 461.83 ab |
| V2 | 17.2 bcdef | 43.96 abcdef | 403.53 abcdefgh | |
| V3 | 16.6 bcdef | 41.63 abcdef | 374.56 bcdefgh | |
| V4 | 18.65 abcd | 47.7 abc | 446.14 abc | |
| V5 | 18.1 abcde | 44.38 abcde | 405.47 abcdefgh | |
| Mean | 18 | 45 | 418 | |
| Analysis of variance | ||||
| Gen (G) | ns | ** | ** | |
| Env (E) | *** | *** | *** | |
| Gen (G) * Env (E) | *** | *** | *** | |
| Env (E): (Combinations of Location * Season) | Gen (G) | Protein (%) | Gluten (%) | Baking Strength (W) |
|---|---|---|---|---|
| MCH 2017 | V1 | 22 a | 64.82 a | 622.86 a |
| V2 | 20.15 ab | 59.12 abc | 524.59 bc | |
| V3 | 18.75 bcd | 55.42 cde | 494 cd | |
| V4 | 20.4 ab | 63 ab | 582.47 ab | |
| V5 | 19.5 abc | 56.45 bcd | 522.16 bc | |
| Mean | 20.16 | 59.76 | 549.22 | |
| MCH 2018 | V1 | 15.34 ghi | 36.54 jk | 330.03 hi |
| V2 | 14.88 hi | 36.03 jk | 297.86 i | |
| V3 | 13.92 i | 32.43 k | 300.97 i | |
| V4 | 15.1 hi | 36.71 jk | 307.25 i | |
| V5 | 15.58 fghi | 36.98 ijk | 345.66 ghi | |
| Mean | 15 | 36 | 316 | |
| MCH 2019 | V1 | 18.27 bcdef | 49.6 defg | 466.3 cde |
| V2 | 17.21 cdefgh | 46.75 fg | 403.97 efg | |
| V3 | 16.41 defghi | 44.13 fghi | 399.31 fg | |
| V4 | 17.98 bcdefg | 50.51 def | 450.77 def | |
| V5 | 18.1 bcdefg | 48.21 efg | 447.4 def | |
| Mean | 18 | 48 | 434 | |
| MCH 2020 | V1 | 17.25 cdefgh | 46.85 fg | 440.29 def |
| V2 | 17.05 cdefgh | 46.32 fgh | 400.17 fg | |
| V3 | 16.75 cdefgh | 45.05 fgh | 407.7 efg | |
| V4 | 17.9 bcdefg | 50.27 def | 448.53 def | |
| V5 | 15.95 efghi | 42.48 ghij | 394.62 fg | |
| Mean | 16.98 | 46.19 | 418.26 | |
| MCH 2021 | V1 | 18.5 bcde | 50.2 def | 472.02 cd |
| V2 | 16.75 cdefgh | 45.5 fgh | 393.25 fgh | |
| V3 | 14.6 hi | 39.27 hijk | 355.43 ghi | |
| V4 | 17.3 cdefgh | 48.62 efg | 433.97 def | |
| V5 | 16.05 defghi | 42.75 ghij | 397.29 fg | |
| Mean | 16.64 | 45.27 | 410.39 | |
| Analysis of variance | ||||
| Gen (G) | *** | *** | *** | |
| Env (E) | *** | *** | *** | |
| Gen (G) * Env (E) | ns | * | *** | |
| Env (E): (Combinations of Location * Season) | Gen (G) | Protein (%) | Gluten (%) | Baking Strength (W) |
|---|---|---|---|---|
| TST 2017 | V1 | 10.85 cde | 24.09 cd | 108.25 efgh |
| V2 | 9.8 e | 21.29 d | 53.47 h | |
| V3 | 10.7 de | 23.86 cd | 89.79 fgh | |
| V4 | 10.45 de | 23.52 cd | 80.97 gh | |
| V5 | 11.7 bcde | 26.44 abcd | 155.73 bcdefg | |
| Mean | 10.7 | 23.84 | 97.64 | |
| TST 2018 | V1 | 12.37 bcde | 27.24 abcd | 215.95 abc |
| V2 | 12.22 bcde | 27.4 abcd | 185.05 abcde | |
| V3 | 13.46 abcde | 30.73 abc | 265.46 a | |
| V4 | 13.1 bcde | 30.42 abc | 222.97 abc | |
| V5 | 13.26 abcde | 30.67 abc | 243.83 ab | |
| Mean | 12.88 | 29.29 | 226.65 | |
| TST 2019 | V1 | 12.41 bcde | 26.32 abcd | 166.04 bcdefg |
| V2 | 15.02 abcd | 31.58 abc | 153.49 bcdefg | |
| V3 | 14.02 abcde | 29.81 abcd | 194.02 abcde | |
| V4 | 13.38 abcde | 29.62 abcd | 166.57 bcdefg | |
| V5 | 13.62 abcde | 30.14 abcd | 210.85 abcd | |
| Mean | 13.69 | 29.49 | 178.19 | |
| TST 2020 | V1 | 12.95 bcde | 25.8 bcd | 162.24 bcdefg |
| V2 | 13.05 bcde | 25.5 bcd | 123.38 defgh | |
| V3 | 13.5 abcde | 26.27 abcd | 170.84 bcdefg | |
| V4 | 13.25 abcde | 27.77 abcd | 155.41 bcdefg | |
| V5 | 14.75 abcd | 30.96 abc | 216.52 abc | |
| Mean | 13.5 | 27.26 | 165.68 | |
| TST 2021 | V1 | 15.5 abc | 30.79 abc | 194.84 abcde |
| V2 | 15.55 ab | 30.16 abcd | 148.55 cdefg | |
| V3 | 17.9 a | 34.85 a | 226.81 abc | |
| V4 | 15.1 abcd | 31.6 abc | 178.55 abcdef | |
| V5 | 15.9 ab | 33.29 ab | 232.98 abc | |
| Mean | 15.99 | 32.14 | 196.34 | |
| Analysis of variance | ||||
| Gen (G) | ns | ** | *** | |
| Env (E) | *** | *** | *** | |
| Gen (G) * Env (E) | ns | ns | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hnizil, O.; Baidani, A.; Khlila, I.; Taghouti, M.; Nsarellah, N.; Amamou, A. Dissecting Genotype by Environment Interactions in Moroccan Wheat: An Advanced Biplot and Heatmap Analysis Unveiling Agronomic, Quality Traits, and Genotypic Stability for Tailored Breeding Strategies. Plants 2024, 13, 1068. https://doi.org/10.3390/plants13081068
Hnizil O, Baidani A, Khlila I, Taghouti M, Nsarellah N, Amamou A. Dissecting Genotype by Environment Interactions in Moroccan Wheat: An Advanced Biplot and Heatmap Analysis Unveiling Agronomic, Quality Traits, and Genotypic Stability for Tailored Breeding Strategies. Plants. 2024; 13(8):1068. https://doi.org/10.3390/plants13081068
Chicago/Turabian StyleHnizil, Oussama, Aziz Baidani, Ilham Khlila, Mouna Taghouti, Nasserelhaq Nsarellah, and Ali Amamou. 2024. "Dissecting Genotype by Environment Interactions in Moroccan Wheat: An Advanced Biplot and Heatmap Analysis Unveiling Agronomic, Quality Traits, and Genotypic Stability for Tailored Breeding Strategies" Plants 13, no. 8: 1068. https://doi.org/10.3390/plants13081068
APA StyleHnizil, O., Baidani, A., Khlila, I., Taghouti, M., Nsarellah, N., & Amamou, A. (2024). Dissecting Genotype by Environment Interactions in Moroccan Wheat: An Advanced Biplot and Heatmap Analysis Unveiling Agronomic, Quality Traits, and Genotypic Stability for Tailored Breeding Strategies. Plants, 13(8), 1068. https://doi.org/10.3390/plants13081068