3.1. Descriptive Statistics for Yield and Yield Components across Multiple Locations
The agronomic characteristics of five genotypes, coded V1 (Faraj), V2 (Itri), V3 (Karim), V4 (Luiza), and V5 (Nassira), were evaluated across five successive growing seasons (from 2016–2017 to 2020–2021) at the experimental station Sidi El Aïdi (SEA). This study defined “environment” as the varying growing seasons at the SEA location. A comprehensive two-factor analysis of variance (ANOVA) was employed to assess the agronomic traits of these genotypes over the different seasons. Post-hoc Tukey tests were conducted, identifying significant differences at the 95% confidence level, as detailed in
Table 3. This analysis provided an insightful depiction of how genotypes V1 through V5 responded to environmental variations across seasons.
The ANOVA findings highlighted the considerable impact of environmental conditions on key agronomic traits. Specifically, the analysis revealed a significant environmental effect on yield (p ≤ 0.001). This pattern was also reflected in other traits, such as thousand kernel weight (TKW), spikes per square meter (Spk/m2), and grains per square meter (G/m2), all showing similarly significant environmental influences. Notably, the environmental factors did not significantly impact biomass nor the number of grains per spike (G/S).
Regarding the genotypic impact, it was observed that the genotype had a significant exclusive effect on the TKW (p ≤ 0.05). Traits like yield, biomass, Spk/m2, G/S, and G/m2 did not exhibit significant differences attributable to genotypic variations. Furthermore, the interaction between genotype and environment (G * E) demonstrated statistical significance for the TKW (p ≤ 0.05) and Spk/m2 (p ≤ 0.01), indicating variability in the performance of different genotypes across the seasons for these traits.
In examining the annual yield performance, genotypes V5 (Nassira), V1 (Faraj), and V4 (Luiza) demonstrated notable variations across different growing seasons. These variations showcased a pronounced increase in yield of 80.7% from the 2016–2017 season to the 2017–2018 season, followed by a decrease of 59.6% in 2018–2019. This trend continued with a significant decline of 65% in 2019–2020 and an extraordinary rebound of 250% in the 2020–2021 season. It is essential to highlight that these yield changes were predominantly influenced by environmental factors rather than solely by genetic attributes of these genotypes.
Regarding analyzing other agronomic traits, the biomass evaluation revealed no significant differences related to genotype or environmental factors. Concurrently, thousand kernel weight (TKW) exhibited notable interactions between the genotype and environment. Among the genotypes, V5 and V4 distinguished themselves in biomass performance, and V5 and V3 in TKW across varying seasons. Despite these observations, it is essential to interpret any direct relationship between biomass and the TKW with caution, considering the complexity of the underlying interactions.
Additionally, this study observed variability in spikes per square meter (Spk/m2), which was attributed to both environmental factors and the genotype–environment (G * E) interaction. This variability and the observed patterns in grains per spike (G/S) and grains per square meter (G/m2) emphasize the genotypes’ capacity to adapt to changing environmental conditions. However, it is critical to acknowledge that the ANOVA results indicated no significant influence of the genotype, environment, or their interaction on G/S and G/m2. This finding indicates that the observed variations in these traits across different seasons at the SEA location cannot be solely attributed to genetic or environmental factors but are likely the result of a more intricate interplay between these elements.
Expanding on the insights gathered from the Sidi El Aidi (SEA) location, our analysis was extended to the experimental station Marchouch (MCH). The comprehensive study at MCH offers a deeper understanding of how environmental factors significantly impact agronomic traits across different growing seasons. The results from MCH, detailed in
Table 4, underscore the notable influence of both genotype and environmental conditions, and the interactions between these factors on vital agronomic traits.
Yield and biomass exhibited notable annual fluctuations. For instance, yield saw a significant increase of 28.1% between the 2016–2017 and 2017–2018 seasons, followed by a dramatic decline and a resurgence in subsequent years. Biomass trends showcased similar variations.
The thousand kernel weight (TKW) and spikes per square meter (Spk/m2) also demonstrated significant oscillations across the study period, reflecting environmental variability.
The grains per spike (G/S) showed dynamic fluctuations with significant genotype (Gen) * environment (Env) interaction effects, as indicated by the analysis of variance. These interactions underscore the complexity of agronomic traits influenced by both genetic and environmental factors.
What was particularly striking was the significant effect of genotype on grains per square meter (G/m2). A compelling example was observed during the 2020–2021 season, where the V1 and V5 genotypes showed remarkably different G/m2 values under the same environmental conditions. V1 recorded 14,693 G/m2, whereas V5 demonstrated a significantly higher 23,895 G/m2 value. This difference was statistically significant according to the ANOVA and was confirmed with the Tukey’s post-hoc test, highlighting the substantial impact of genetic factors on this trait.
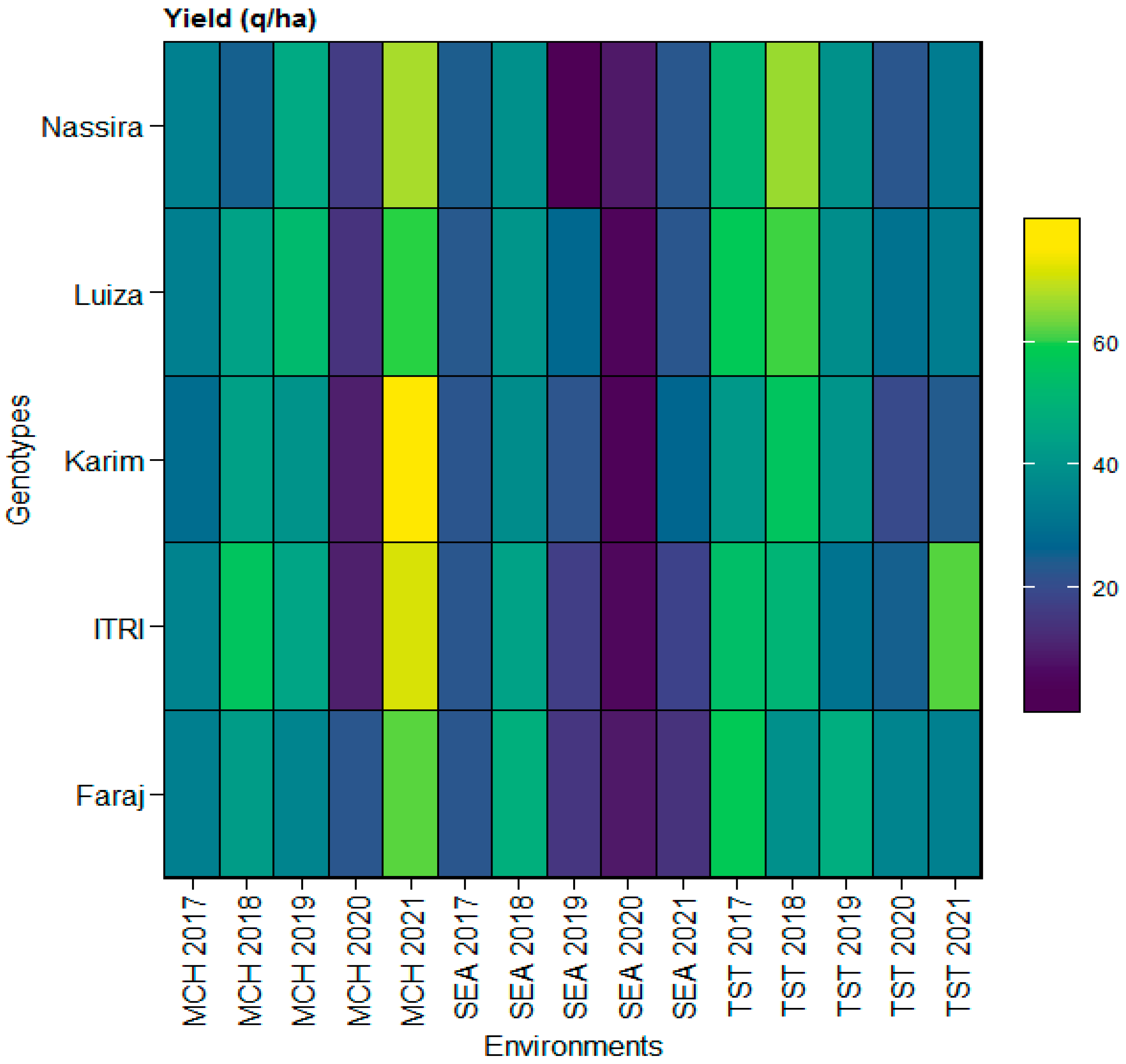
Incorporating the heatmap analysis, it became discernible that the yield variations were intricately linked to both genotype and environmental conditions at each location. The heatmap provided a comprehensive visual summary of the yield dynamics across different genotypes under varying environments from the SEA, MCH, and TST locations the years from 2017 to 2021. For instance, genotypes like Faraj (V1) and Itri (V2) demonstrated consistent yield performances under the SEA environments, corroborating the quantitative findings from SEA. Conversely, the heatmap also vividly captured the pronounced yield variations exhibited by genotypes like Luiza (V4) and Nassira (V5) in the TST and MCH environments, reinforcing the observed fluctuations in yield in these locations. This integrative approach combined quantitative analysis and visual representation, enabling a more nuanced understanding of the multifaceted interactions between genotype, environment, and yield. It also offered valuable insights into different genotypes’ adaptive capacities and yield potentials (V1–V5) under varied environmental conditions, as depicted in
Figure 1.
The two-way analysis of variance (ANOVA) conducted across different locations, including the TST station, revealed distinct patterns in the effects of genotype and environment on agronomic traits. The results for the TST location, as detailed in
Table 5, show significant genotypic effects on biomass, thousand kernel weight (TKW), spikes per square meter (Spk/m
2), and grains per spike (G/S), all at
p < 0.01. These findings at TST starkly contrast with other environments like SEA and MCH, where such genotypic effects were less pronounced.
At TST, the significant genotypic effects on biomass also highlighted a notable interaction between genotype and environment (genotype * environment). For instance, the V3 genotype demonstrated considerable variation in biomass values, increasing from 79.5 q/ha in 2017 to 152.5 q/ha in 2018. Similarly, the V2 genotype fluctuated from 107.5 q/ha in 2019 to 59.75 q/ha in 2020. These changes underscore the significant interaction effect, where different genotypes responded uniquely to environmental conditions over the years.
The environmental impact was notably significant on all traits except for the grains per spike (G/S) trait. For instance, the average biomass at TST showed considerable fluctuations over the years, reflecting environmental variability. In 2017, the average biomass was 105.3 q/ha, increasing to 140.83 q/ha in 2018, then decreasing to 120 q/ha in 2019, and further declining to 74.99 q/ha in 2020. However, a significant recovery was observed in 2021, with the average biomass rising to 146.18 q/ha. These variations align with the ANOVA results’ highly significant environmental differences (p < 0.001).
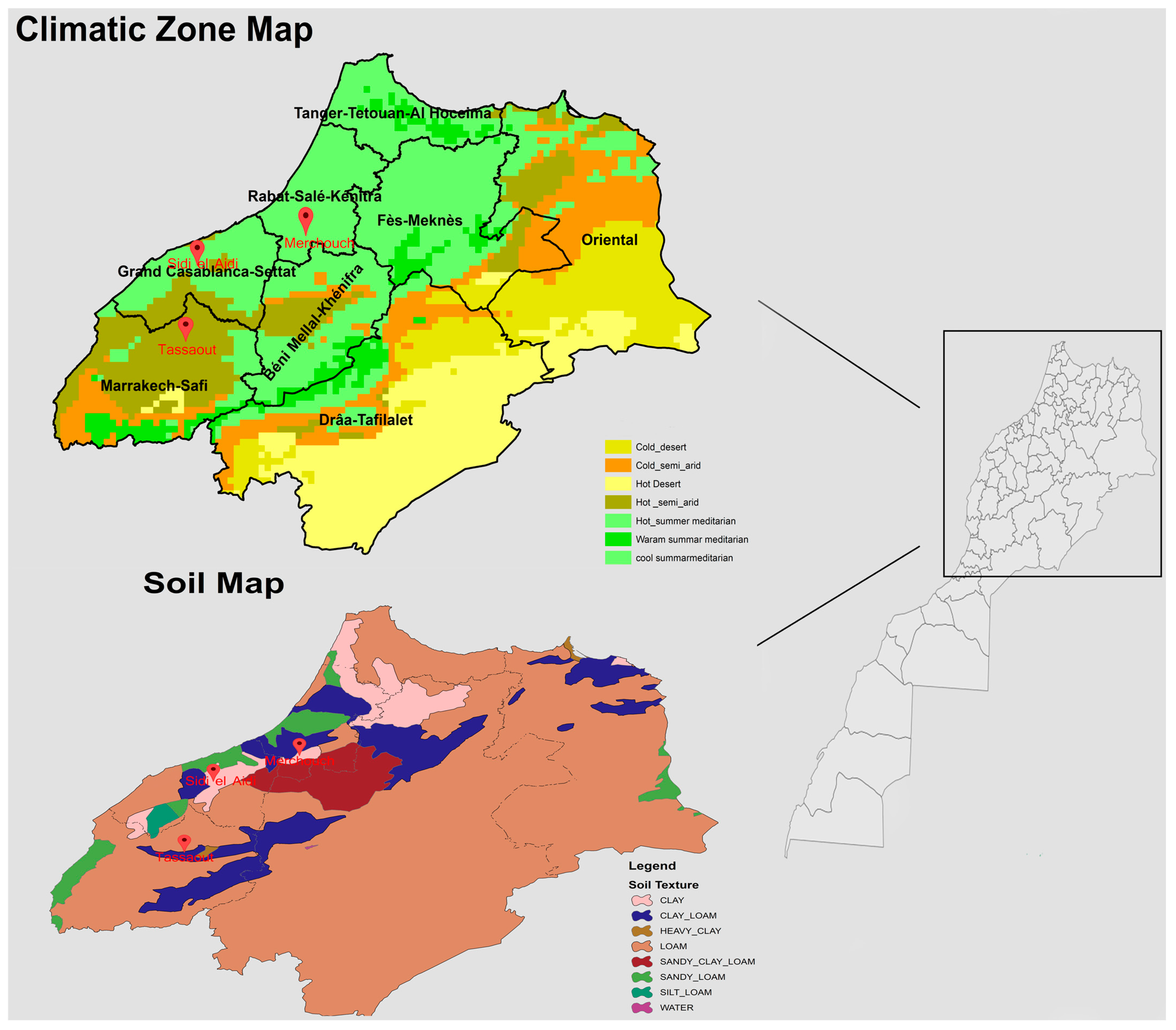
Further analyses, as detailed in
Table 1, elucidate the role of soil type in influencing wheat growth. The soil types at SEA (Vertisol), MCH (Cambisol), and TST (Alfisol) significantly impact wheat growth. Additionally, as indicated in
Figure 2, soil texture varies from clay at SEA to clay loam at MCH and TST. These variations in soil texture affect water retention, aeration, and nutrient availability, influencing wheat traits like biomass, TKW, Spk/m
2, and G/S.
The variability in precipitation from 2016 to 2021 also showed a substantial effect on wheat yields and other traits. For example, SEA experienced a precipitation low of 210 mm in 2018–2019 and a high of 505 mm in 2017–2018. These fluctuations in precipitation directly affect wheat yields and biomass, with increased precipitation typically leading to higher yields.
The distinct response of different wheat genotypes under varying environmental conditions, including soil type, texture, and precipitation, underscores the need for targeted breeding programs. Breeding programs should focus on genetic potential and environmental adaptability to optimize wheat production under specific agroecological zones.
The complex interplay between soil, the climate, and genotype is crucial in determining the agronomic traits of wheat. The significant variations observed across locations highlight the need for context-specific agronomic practices and breeding strategies. Understanding these dynamics can aid in developing wheat varieties that are resilient and high-yielding under varying environmental conditions. These findings from TST, in particular, underscore the importance of targeted breeding and management strategies that consider both genetic potential and environmental adaptability.
3.4. Comprehensive Stability Analysis
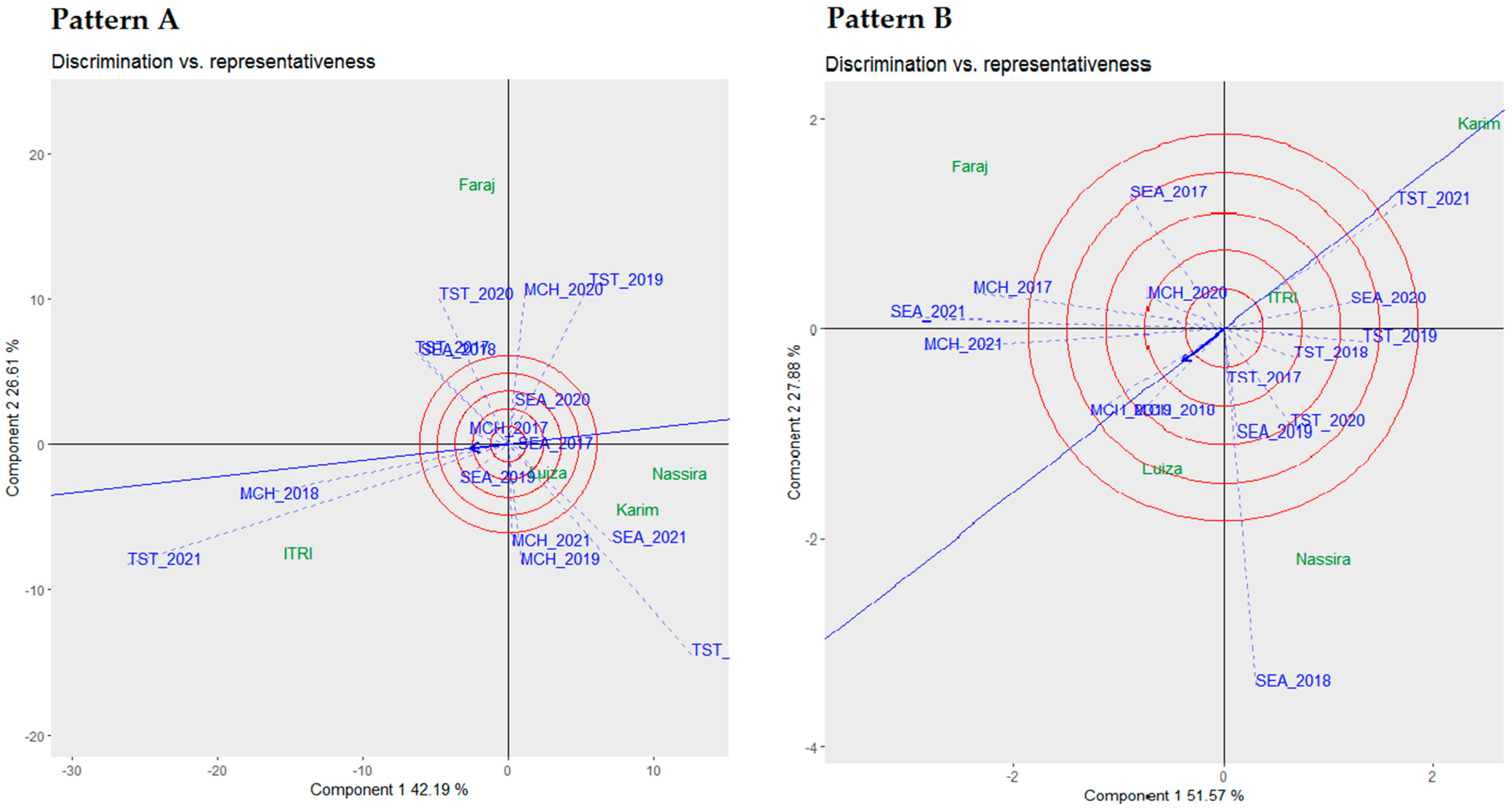
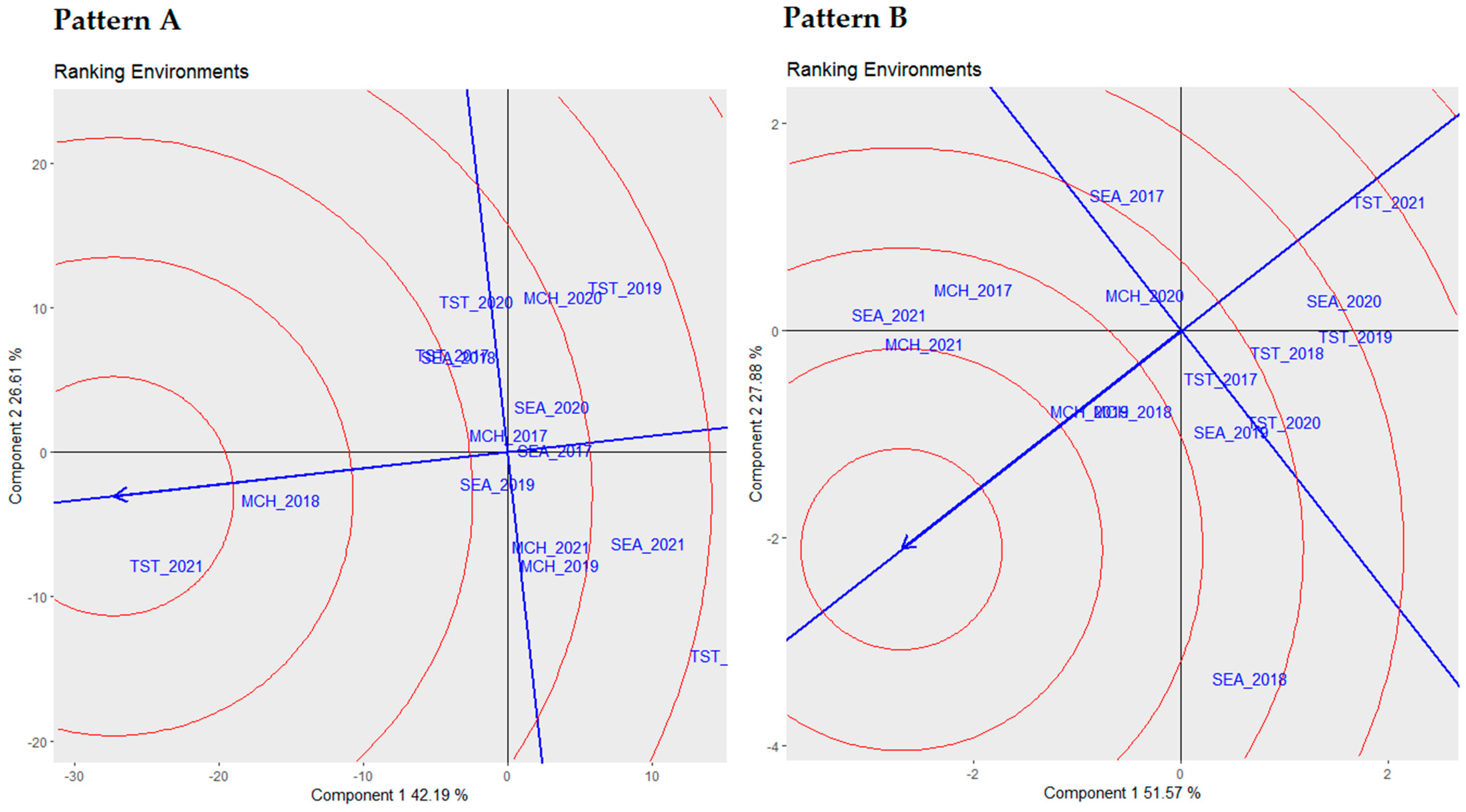
In our comprehensive analysis, as detailed in
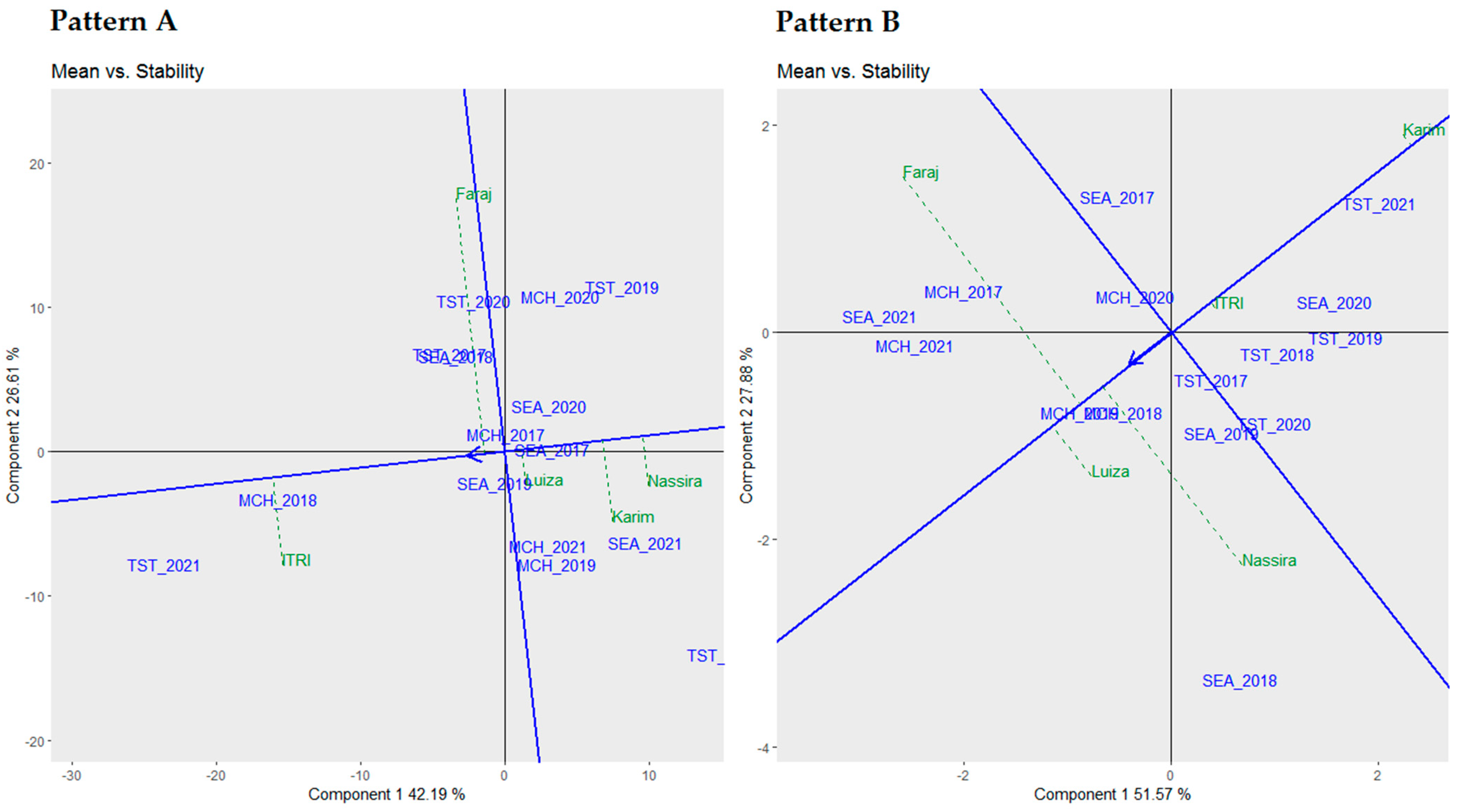
Table 6, we employed three robust statistical metrics, namely Francis cumulative values, Wricke’s ecovalence (W), and Shukla’s stability variance (σ2), to evaluate the stability of five distinct genotypes across multiple environments. The choice of multiple stability parameters was driven by the need to capture the multifaceted nature of genotype–environment interactions. Each metric offers unique insights, ranging from mean performance (Francis cumulative values) to interaction variances (Wricke’s ecovalence and Shukla’s stability variance). This provides a more nuanced and complete understanding of genotypic stability. The genotypes were ranked based on these stability metrics to provide an integrated view of their performance.
Contrary to our initial expectations, “V3” emerged as the most stable genotype according to Shukla’s stability variance (σ2) with a score of 400, leading to its first-place ranking (GR = 1) in this category. However, it displayed less stability in Francis cumulative values with a score of 34, placing it lower in that ranking.
“V1”, on the other hand, demonstrated exceptional stability under Wricke’s ecovalence (W) with a score of 3607,727, meriting the top rank in this specific metric. Interestingly, “Faraj” also showed strong performance in Shukla’s stability variance (σ2), ranked second with a score of 457, indicating its overall stability across varied conditions.
Meanwhile, “V4” exhibited noteworthy stability in Francis cumulative values, leading the category with a score of 49, suggesting a solid adaptability to different environments. In Shukla’s stability variance (σ2), “V4” maintained a solid performance with a score of 333. However, it ranked third in this metric, indicating specific stability traits that may be advantageous under certain environmental scenarios.
These findings highlight that while specific genotypes like “V3” and “V1” show strong stability across different metrics, each genotype possesses unique stability characteristics. This suggests the potential for specific genotypic advantages under certain environmental conditions, underscoring the need for a nuanced approach in selecting genotypes for diverse agricultural settings.
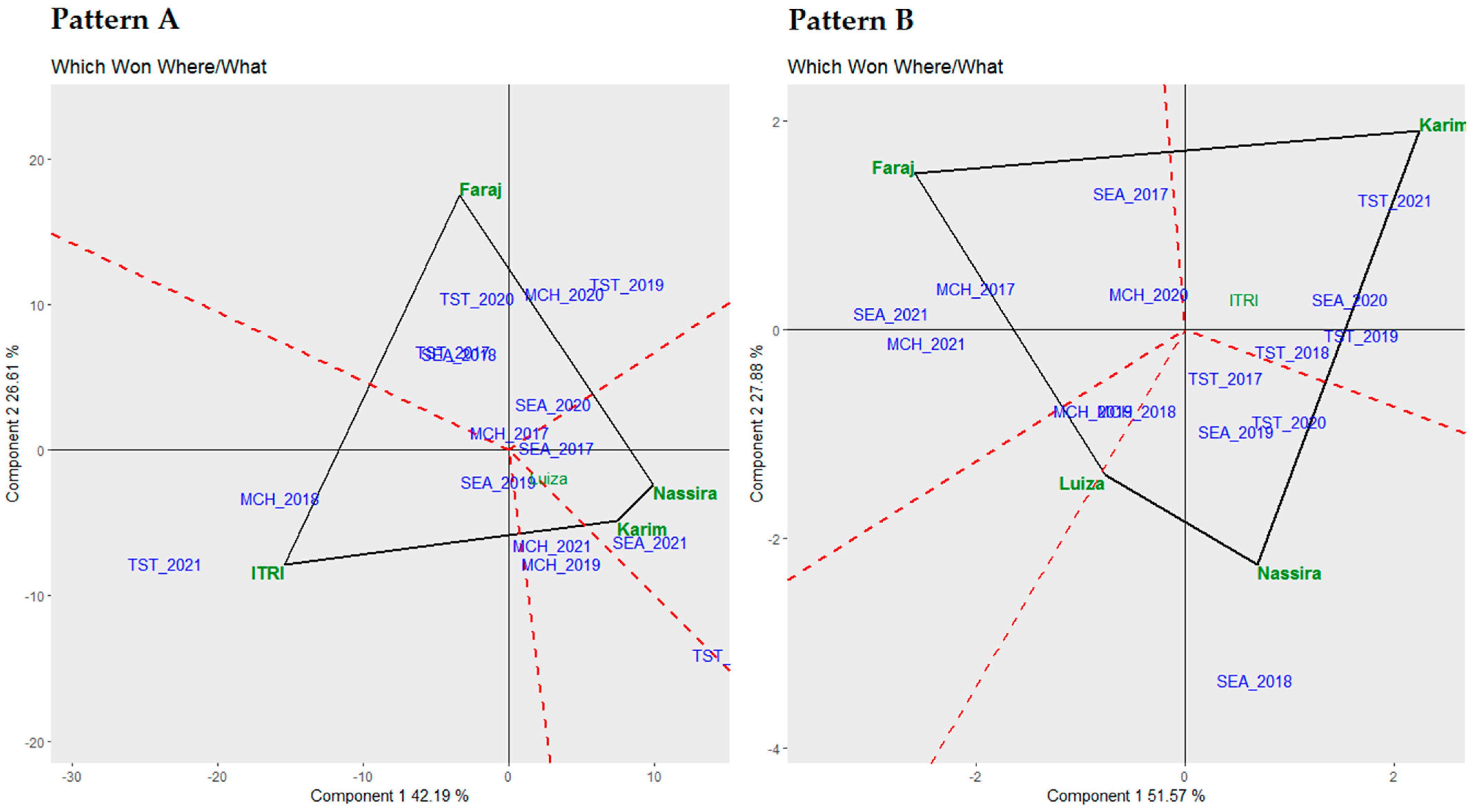
Upon establishing the stability of these genotypes, it was crucial to comprehend their mean performance in correlation with their stability, which was visually represented in the following biplots.
3.7. Descriptive Statistics for Quality Traits across Environments
At the SEA location, significant environmental effects (
p < 0.001) were observed on protein content, along with notable genotype–environment interaction effects (
p < 0.001). The protein content in variety V1 exhibited marked fluctuations, as shown in
Table 7. Specifically, the protein content ranged from 19.46% in 2017, decreased to 13.80% in 2020, and rose to 20.90% in 2021. Comprehensively detailed in
Table 7, these variations underscore the combined influence of environmental conditions and genotype–environment interactions on protein content.
A genotypic influence (p < 0.01) was observed regarding gluten content at SEA. It was also significantly influenced via both the environment (p < 0.001) and the genotype–environment interaction (p < 0.001), and the 2016–2017 growth season marked the peak of mean gluten content at 47.65%, where both Faraj and ITRI excelled, showcasing values of 49.63% and 49.9%, respectively. The lowest mean was documented in 2019–2020, settling at 36.0%, with Faraj recording the lowest value of 31.26%.
Baking strength at SEA revealed significant distinctions due to genotype (p < 0.01) and was profoundly shaped by the environment (p < 0.001), along with the genotype–environment interaction (p < 0.001). During the 2016–2017 growth span, the mean baking strength stood at 449.03 W, with Faraj ascending to the pinnacle spot with 494.29 W. A decline followed in subsequent years, culminating in the nadir in 2019–2020 at an average of 332 W. Nevertheless, the 2020–2021 season marked a revival, averaging at 418 W, with Faraj recording a notable strength of 461.83 W.
For the Marchouch (MCH) site, protein content displayed a paramount genotype influence (
p < 0.001) and was strongly influenced by environmental factors (
p < 0.001), as detailed in
Table 8. However, the interplay between genotype and environment was found to be insignificant (ns). During the 2016–2017 growth season, the mean protein content for all varieties was 20.16%, with Faraj attaining the peak value of 22%. A marked decrease occurred in 2017–2018, causing the mean protein content to drop to 15%. The 2020–2021 season recorded a mean protein content of 16.64%, with Faraj showing a significant resurgence to 18.5%.
Regarding gluten content at MCH, a dominant genotypic impact (p < 0.001) was identified, coupled with a pronounced environmental influence (p < 0.001). The interaction between genotype and environment was present, though lower in its magnitude (p < 0.05). The 2016–2017 growth span marked the apex in mean gluten content at 59.76%, with both Faraj and Luiza making a solid presence, sequentially registering values of 64.82% and 63%, respectively. A trough was hit in 2017–2018, pinning the mean gluten content at 36%, with Karim’s value dwindling to 32.43%.
Baking strength in MCH demonstrated an overarching genotypic impact (p < 0.001) and was decisively molded through environmental factors (p < 0.001). The interrelation between genotype and environment stood out emphatically (p < 0.001). During the 2016–2017 growth phase, the mean baking strength settled at 549.22 W, with Faraj elevating to a peak strength of 622.86 W. Subsequent growth periods witnessed a waning, bottoming out in 2017–2018 with a mean value of 316 W.
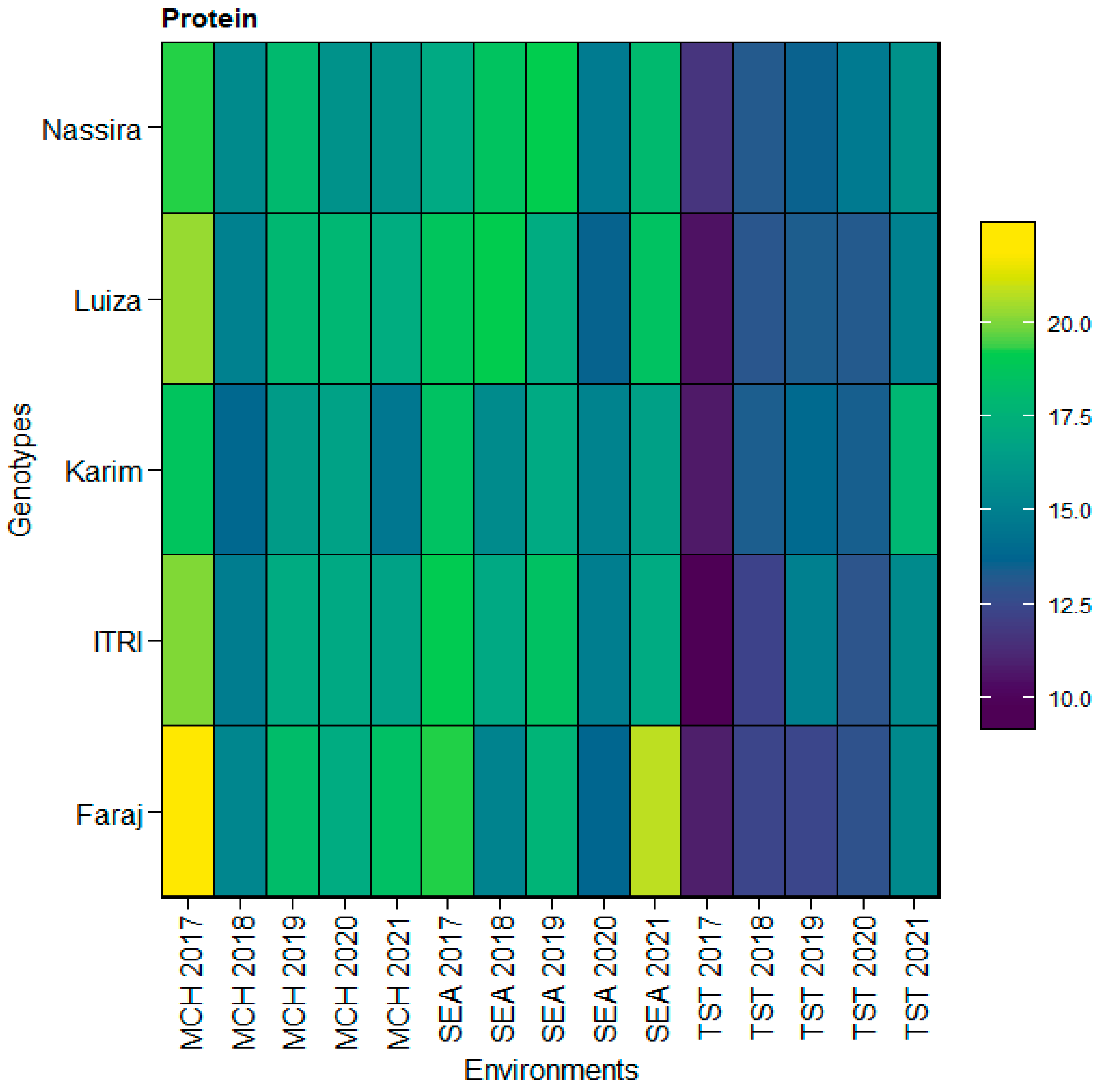
Upon integrating the insights from the heatmap analysis, we observed distinct patterns in protein content across the genotypes Faraj, Itri, Karim, Luiza, and Nassira in the environments SEA, MCH, and TST from 2017 to 2021. The heatmap illustrated the nuanced interactions between genotypes and environments, revealing areas of high protein content, especially in Faraj, across multiple environments. This corroborates the consistently high values observed in the SEA and MCH sites. Conversely, areas of lower protein content were also depicted, which aligned with the lower averages noted for Itri in the TST location. This visual representation enhanced our understanding of the intricate dynamics of protein content across different environments and genotypes, offering a holistic view of the variability and specific adaptations unique to each genotype, as depicted in
Figure 7.
At the Tassaout (TST) location, quality traits, including protein content, gluten content, and baking strength, exhibited varied influences from genotypic and environmental factors, as comprehensively detailed in
Table 9. The protein content displayed a non-significant effect from the genotype but was significantly influenced by environmental conditions (
p < 0.001). The average protein content across the growing years fluctuated, starting from 10.70% during the 2016–2017 season, increasing to 12.88% in 2017–2018, and reaching a peak of 15.99% in 2020–2021. These variations in protein content over the years at TST, including the specific changes for each season, are meticulously documented in
Table 9. This table clearly depicts how environmental factors across different growing seasons can significantly influence key quality traits like protein content in wheat.
The gluten content revealed a notable effect from genotype (p < 0.01) and was profoundly influenced by environmental conditions (p < 0.001). In contrast, the interaction between genotype and environment did not show significant effects (ns). The highest average gluten content was recorded in 2020–2021 at 32.14%, with Karim and Nassira showing elevated values of 34.85% and 33.29%, respectively. The lowest average gluten content was observed during the 2016–2017 growing season, at 23.84%, with Itri registering the lowest value of 21.29%.
Baking strength at TST displayed a highly significant genotypic effect (p < 0.001) and was also markedly influenced by the environment (p < 0.001). The genotype–environment interaction was non-significant (ns). During the 2016–2017 growing season, the average baking strength was 97.64 W, with Itri recording the lowest value of 53.47 W. A substantial increase was observed in the subsequent growing seasons, reaching its peak in 2017–2018 with an average of 226.65 W. Karim showed the highest value under this growing season, registering a baking strength of 265.46 W.
The data revealed a notable trend in protein content at the Tassaout (TST) location. TST, an irrigated site conventionally anticipated to be optimal for crop yield and quality conditions, has the lowest protein content among the three surveyed sites. In contrast, the SEA site, which had lower yields, showed a higher protein content. The data from SEA and TST illustrated an inverse relationship between yield and protein content. Such results suggest that while irrigation may enhance yield, particularly in traditionally arid regions like TST, it may simultaneously reduce grain protein concentration. This dynamic is crucial for breeding programs aiming to optimize wheat yield and protein content.
Expanding on the previous insights, we then focused on examining the intricate relationships between agronomic and quality traits within the framework of genotype–environment interactions.
3.8. Correlations among Agronomic and Quality Traits in the Context of Genotype–Environment Interactions
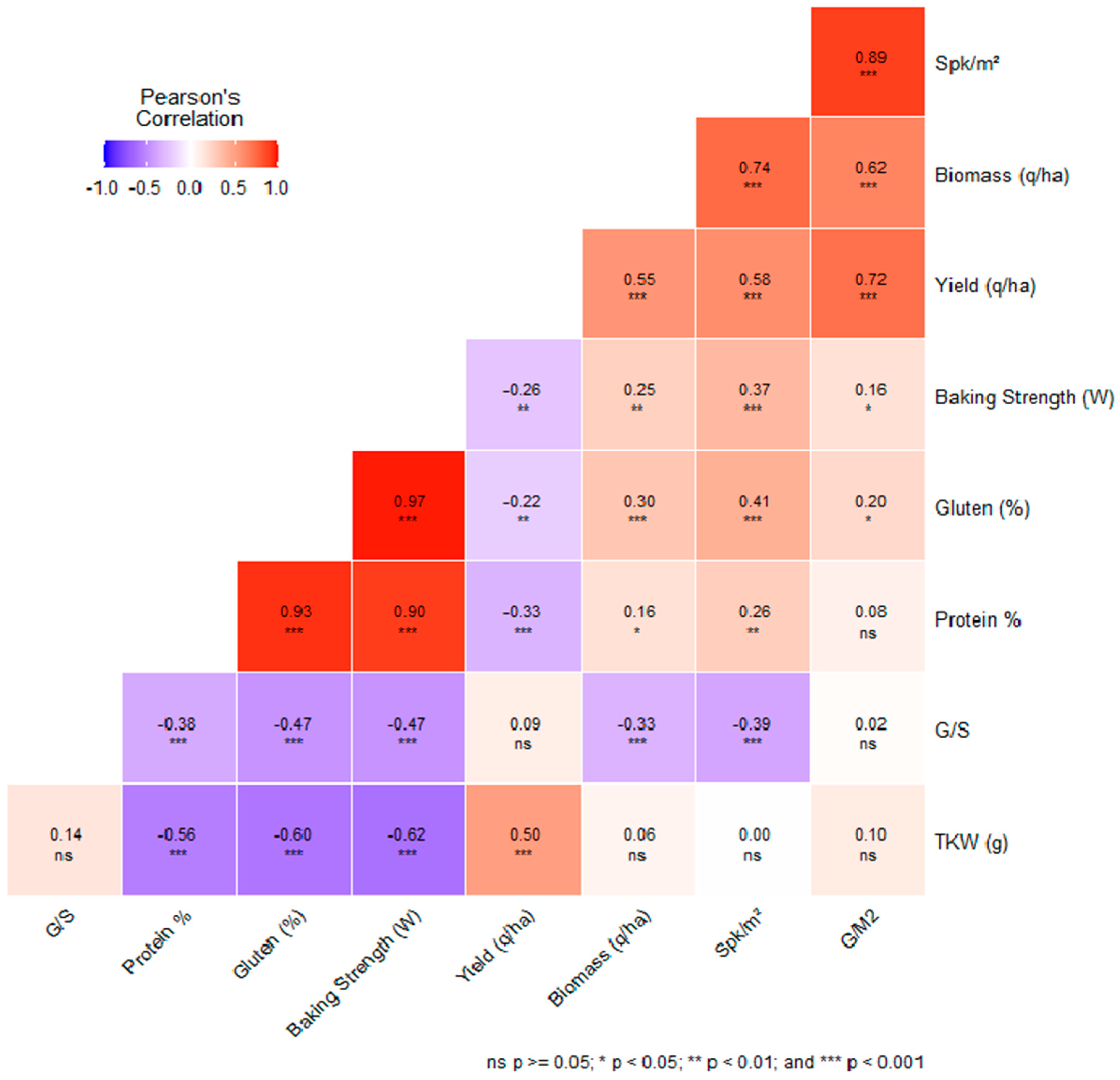
In the context of genotype–environment interactions, agronomic traits, such as yield, biomass, thousand kernel weight (TKW), spikes per square meter (Spk/m
2), grain-to-straw ratio (G/S), and grain yield per square meter (G/m
2), were evaluated alongside quality traits, such as protein percentage, gluten percentage, and baking strength. Pearson correlation coefficients and their respective
p-values were calculated to gauge the relationships between these agronomic and quality traits (
Figure 8).
For instance, a moderate positive correlation was observed between yield and biomass (r = 0.55, p < 0.05). This suggests that genotypes with increased biomass are generally associated with higher yields. Moreover, there was a negative correlation between yield and protein content (r = −0.33, p < 0.001), indicating that augmenting yield could potentially reduce protein concentration. The relationship between the density of spikes per square meter (Spk/m2) and biomass showed a significant positive trend (r = 0.74, p < 0.001), emphasizing that a rise in spike density is linked to greater biomass levels.
In addition, a prominent positive correlation was identified between grain yield per square meter (G/m2) and the aggregate yield (r = 0.72, p < 0.001). This relationship highlights that an increase in grain yield per square meter significantly boosts the total yield. As a result, G/m2 can be considered a reliable indicator for predicting yield outcomes.
In examining quality traits, a robust positive correlation was identified between protein and gluten contents (r = 0.93, p < 0.001), signifying a robust relationship between these two elements. This finding posits that a selection favoring higher protein content could concurrently augment gluten levels, a critical aspect for end-use quality.
Lastly, a robust positive correlation was noted between gluten content and baking strength (r = 0.97, p < 0.001). This observation underscores that gluten content significantly impacts baking strength, accentuating the significance of gluten quality in determining a genotype’s suitability for specific end applications.
The interactions and correlations presented here provide invaluable insights. The subsequent section aimed to contextualize these findings within the broader scientific discourse, offering a detailed understanding of the genotype–environment interaction effects on agronomic and quality traits.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}