
Reduced Root Cortical Tissue with an Increased Root Xylem Investment Is Associated with High Wheat Yields in Central China

 , ,
, , 
Abstract
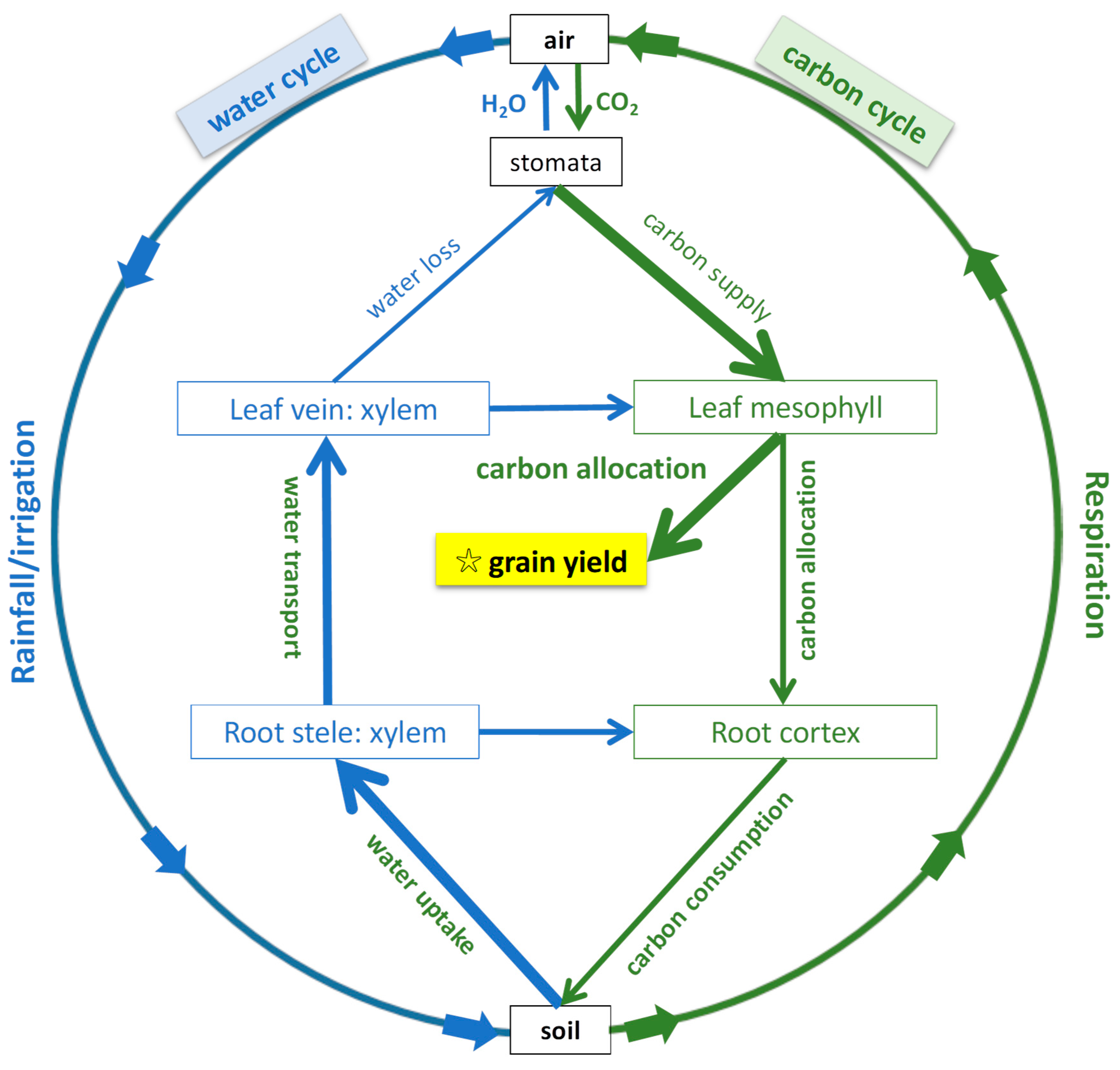
:1. Introduction
2. Materials and Methods
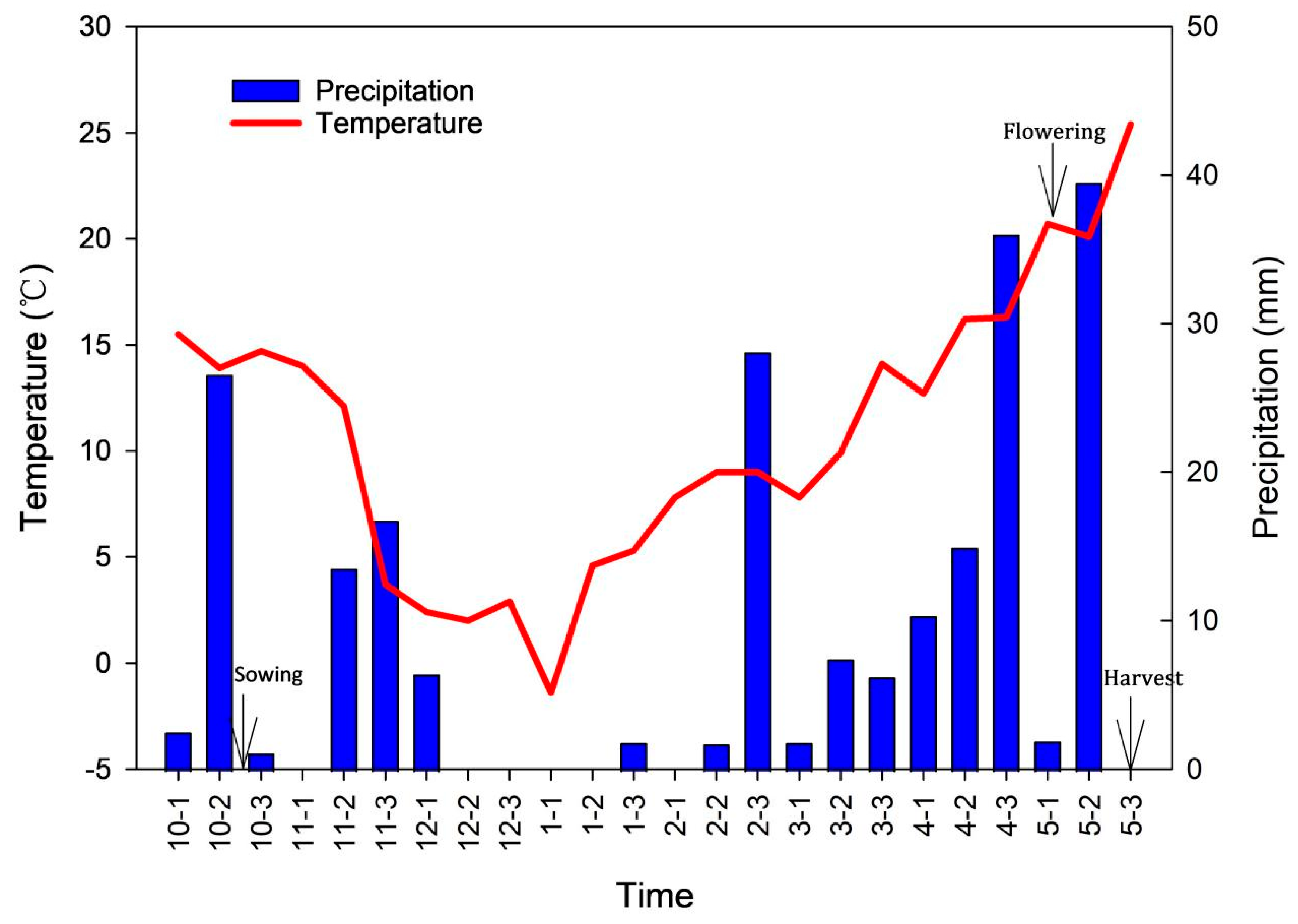
2.1. Plant Materials and Growth Conditions
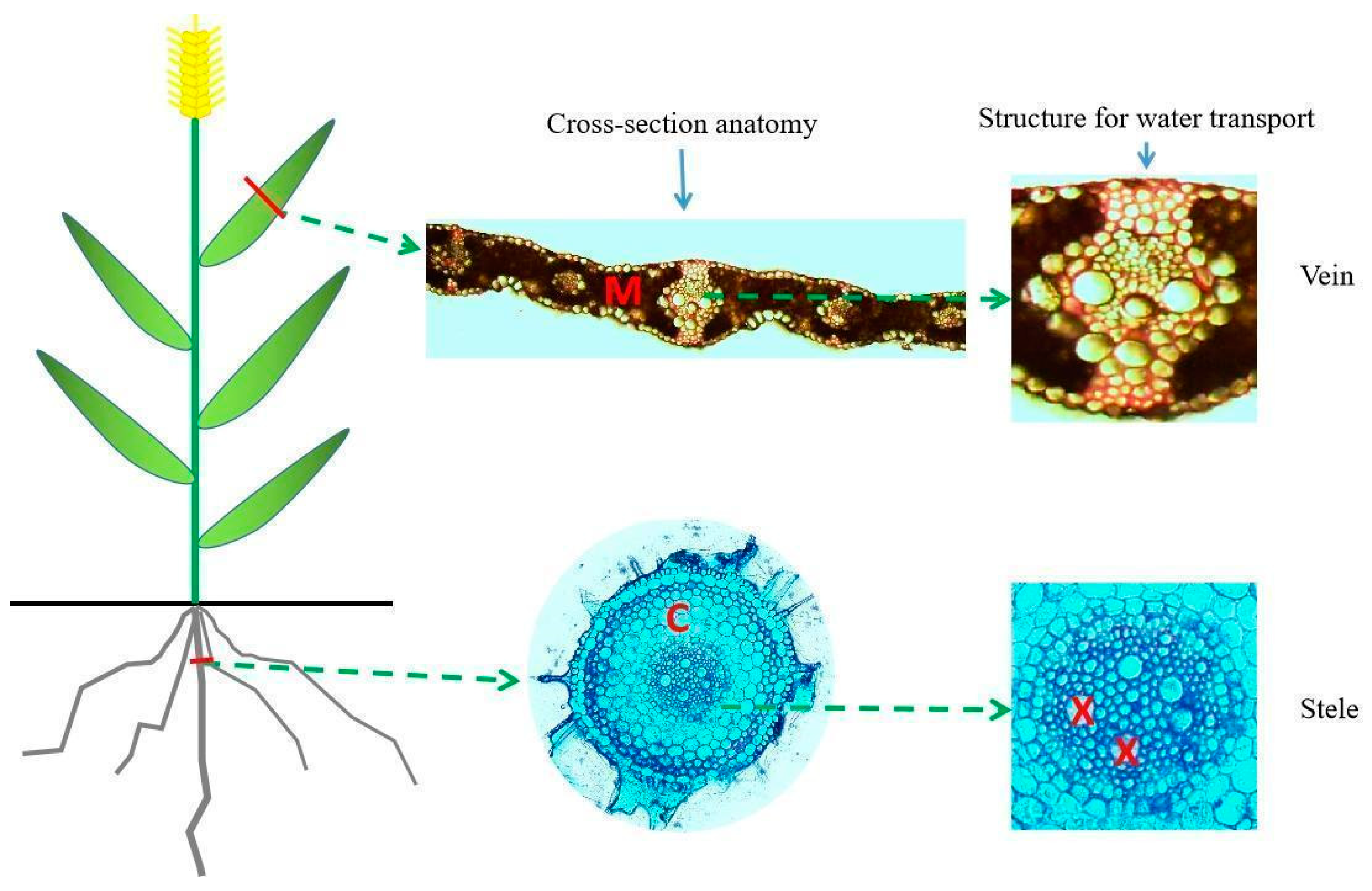
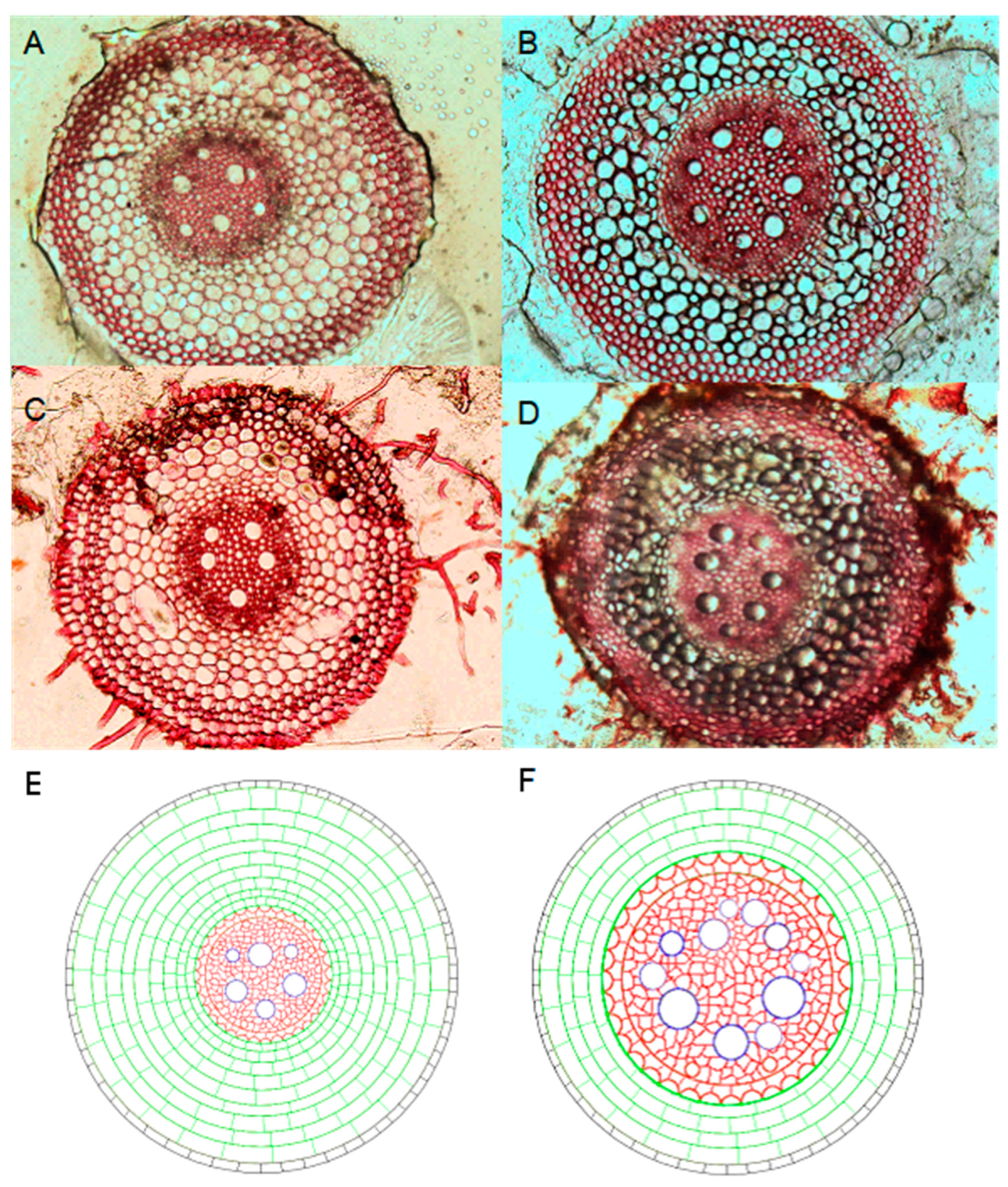
2.2. Measurements of the Leaf and Root Anatomical Traits
2.3. Statistical Analyses
3. Results
3.1. Yield and Yield Components
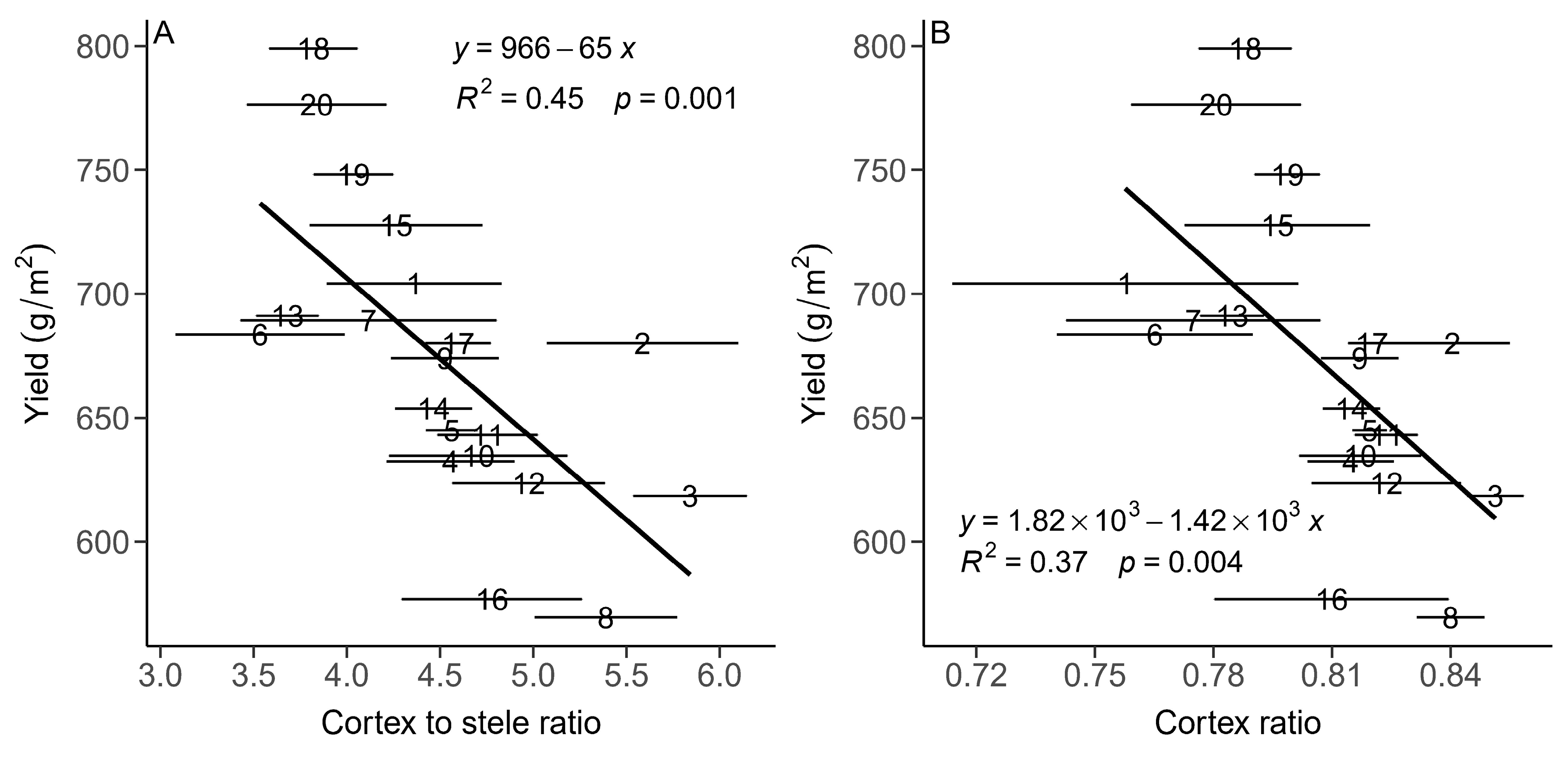
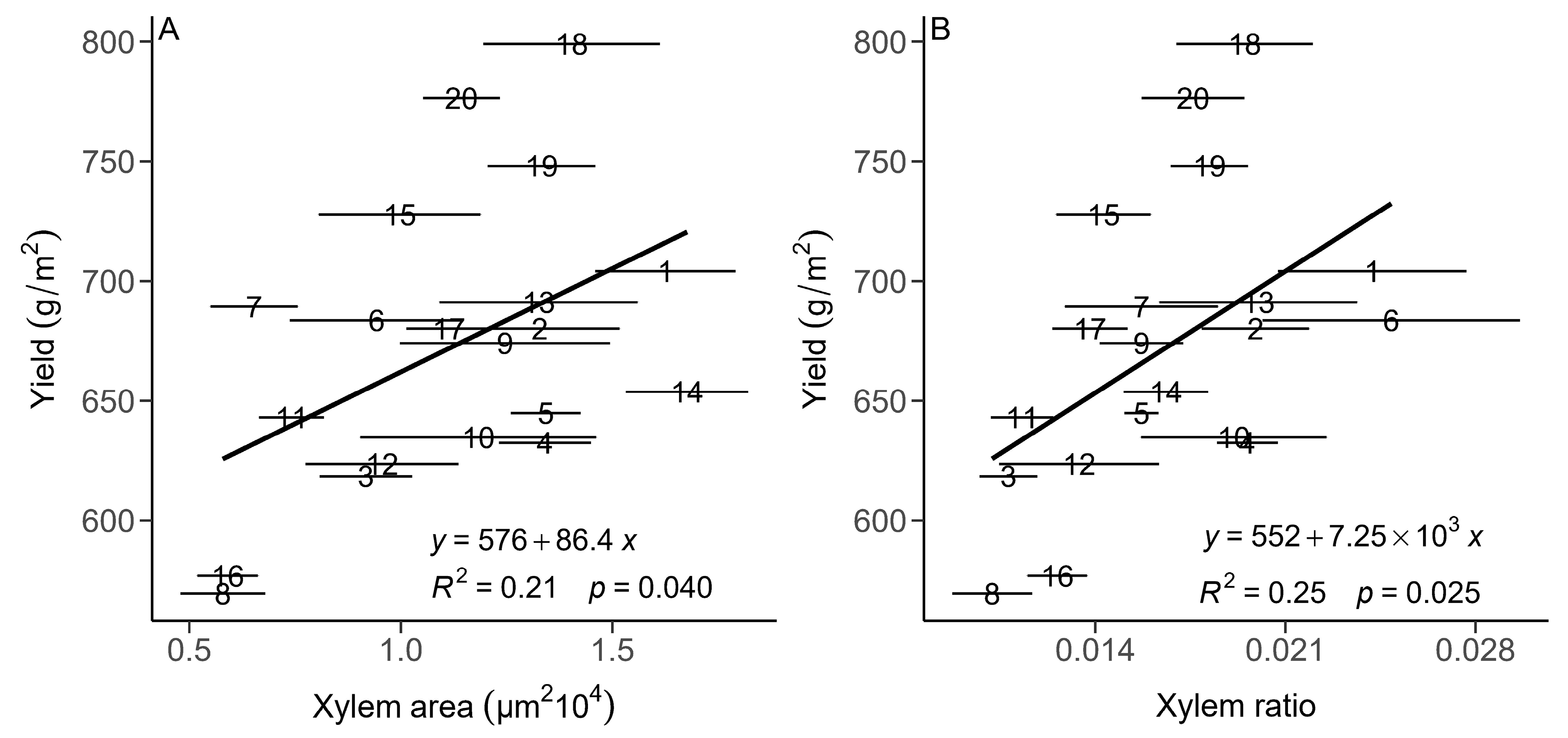
3.2. The Relationships between the Root and Leaf Anatomical Traits and Population Yield
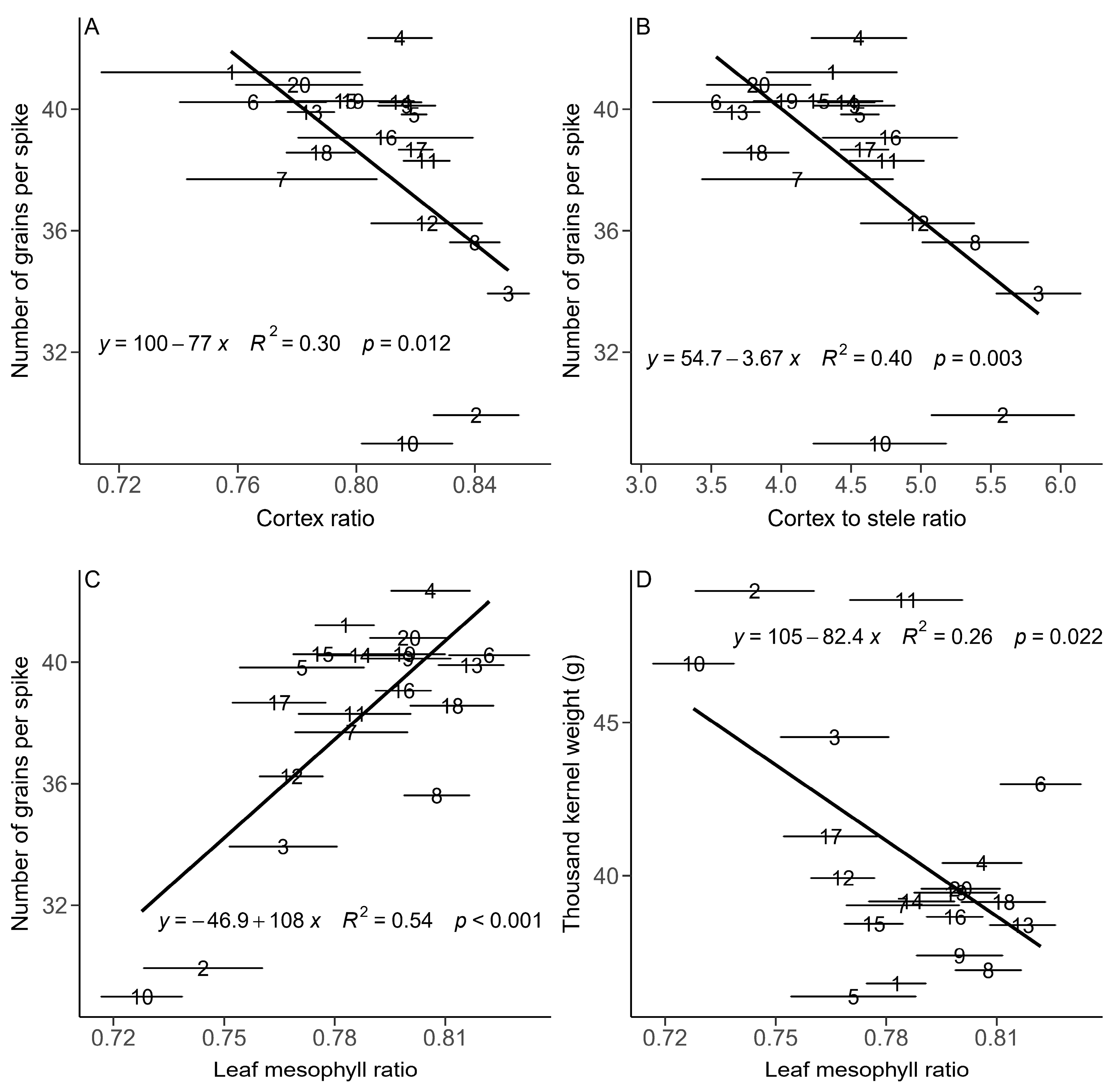
3.3. Relationships between the Root and Leaf Anatomical Traits and Yield Components
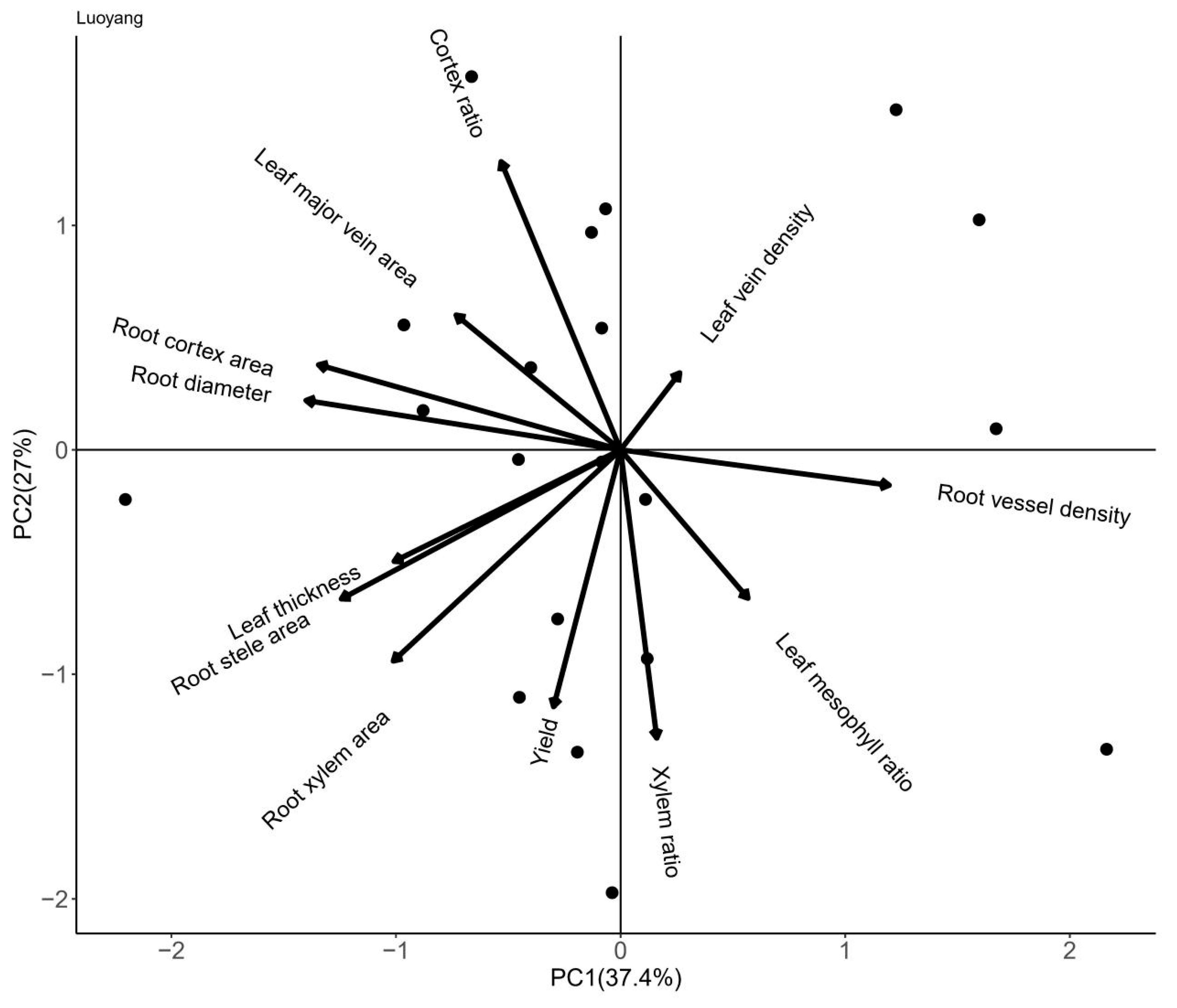
3.4. Results of Principal Component Analysis of Root and Leaf Anatomical Traits
4. Discussion
4.1. Root Anatomical Traits and Yield
4.2. Leaf Anatomical Traits and Yield
4.3. Root and Leaf Anatomical Traits and Yield Components
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weiner, J. Looking in the Wrong Direction for Higher-Yielding Crop Genotypes. Trends Plant Sci. 2019, 24, 927–933. [Google Scholar] [CrossRef]
- Milla, R.; Osborne, C.P.; Turcotte, M.M.; Violle, C. Plant domestication through an ecological lens. Trends Ecol. Evol. 2015, 30, 463–469. [Google Scholar] [CrossRef] [PubMed]
- York, L.M.; Galindo-Castaneda, T.; Schussler, J.R.; Lynch, J.P. Evolution of US maize (Zea mays L.) root architectural and anatomical phenes over the past 100 years corresponds to increased tolerance of nitrogen stress. J. Exp. Bot. 2015, 66, 2347–2358. [Google Scholar] [CrossRef] [PubMed]
- Lux, A.; Luxová, M.; Abe, J.; Morita, S. Root cortex: Structural and functional variability and responses to environmental stress. Root Res. 2004, 29, 117–131. [Google Scholar] [CrossRef]
- Schröder, M.; Kunz, U.; Stelzer, R.; Lehmann, H. On the evidence of a diffusion barrier in the outer cortex apoplast of cress-roots (Lepidium sativum), demonstrated by analytical electron microscopy. J. Plant Physiol. 2002, 159, 1197–1204. [Google Scholar] [CrossRef]
- Zhou, M.; Bai, W.; Li, Q.; Guo, Y.; Zhang, W. Root anatomical traits determined leaf-level physiology and responses to precipitation change of herbaceous species in a temperate steppe. New Phytol. 2021, 229, 1481–1491. [Google Scholar] [CrossRef] [PubMed]
- Chimungu, J.G.; Brown, K.M.; Lynch, J.P. Reduced Root Cortical Cell File Number Improves Drought Tolerance in Maize. Plant Physiol. 2014, 166, 1943–1955. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Castañeda, T.; Brown, K.M.; Lynch, J.P. Reduced root cortical burden improves growth and grain yield under low phosphorus availability in maize. Plant Cell Environ. 2018, 41, 1579–1592. [Google Scholar] [CrossRef]
- Kong, D.; Wang, J.; Valverde-Barrantes, O.J.; Kardol, P. A framework to assess the carbon supply–consumption balance in plant roots. New Phytol. 2021, 229, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Pedersen, O.; Nakazono, M.; Tsutsumi, N. Key root traits of Poaceae for adaptation to soil water gradients. New Phytol. 2021, 229, 3133–3140. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Brown, K.M.; Lynch, J.P. Root cortical aerenchyma improves the drought tolerance of maize (Zea mays L.). Plant Cell Environ. 2010, 33, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cao, J.-J.; Yang, Q.-P.; Wu, M.-Z.; Zhao, Y.; Kong, D.-L. The worldwide allometric relationship in anatomical structures for plant roots. Plant Divers. 2023, 45, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Fageria, N.K.; Virupax, C.B.; Ralph, C. Physiology of Crop Production; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Lynch, J.P. Root phenes that reduce the metabolic costs of soil exploration: Opportunities for 21st century agriculture. Plant Cell Environ. 2015, 38, 1775–1784. [Google Scholar] [CrossRef]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-H.; Weiner, J.; Yu, M.-X.; Li, F.-M. Evolutionary agroecology: Trends in root architecture during wheat breeding. Evol. Appl. 2019, 12, 733–743. [Google Scholar] [CrossRef]
- Weiner, J.; Du, Y.; Zhang, C.; Qin, X.; Li, F. Evolutionary agroecology: Individual fitness and population yield in wheat (Triticum aestivum). Ecology 2017, 98, 2261–2266. [Google Scholar] [CrossRef] [PubMed]
- Hendel, E.; Bacher, H.; Oksenberg, A.; Walia, H.; Schwartz, N.; Peleg, Z. Deciphering the genetic basis of wheat seminal root anatomy uncovers ancestral axial conductance alleles. Plant Cell Environ. 2021, 44, 1921–1934. [Google Scholar] [CrossRef] [PubMed]
- Donald, C.M. Competitive plants, communal plants, and yields in wheat crops. In Wheat Science—Today and Tomorrow; Evans, L.T., Peacock, W.J., Eds.; Cambridge University Press: Cambridge, UK, 1981; pp. 223–247. [Google Scholar]
- Fahad, S.; Khan, F.A.; Pandupuspitasari, N.; Hussain, S.; Khan, I.A.; Saeed, M.; Saud, S.; Hassan, S.; Adnan, M.; Amanullah; et al. Suppressing photorespiration for the improvement in photosynthesis and crop yields: A review on the role of S-allantoin as a nitrogen source. J. Environ. Manag. 2019, 237, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Lovett Doust, J. Plant reproductive strategies and resource allocation. Trends Ecol. Evol. 1989, 4, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Du, Y.-L.; He, J.; Turner, N.C.; Wang, B.-R.; Zhang, C.; Cui, T.; Li, F.-M. Recently-released genotypes of naked oat (Avena nuda L.) out-yield early releases under water-limited conditions by greater reproductive allocation and desiccation tolerance. Field Crop. Res. 2017, 204, 169–179. [Google Scholar] [CrossRef]
- Wang, F.; Xie, R.; Ming, B.; Wang, K.; Hou, P.; Chen, J.; Liu, G.; Zhang, G.; Xue, J.; Li, S. Dry matter accumulation after silking and kernel weight are the key factors for increasing maize yield and water use efficiency. Agric. Water Manag. 2021, 254, 106938. [Google Scholar] [CrossRef]
- Zhang, X.-F.; Luo, C.-L.; Ren, H.-X.; Mburu, D.; Wang, B.-Z.; Kavagi, L.; Wesly, K.; Nyende, A.B.; Xiong, Y.-C. Water productivity and its allometric mechanism in mulching cultivated maize (Zea mays L.) in semiarid Kenya. Agric. Water Manag. 2021, 246, 106647. [Google Scholar] [CrossRef]
- He, J.; Du, Y.-L.; Wang, T.; Turner, N.C.; Yang, R.-P.; Jin, Y.; Xi, Y.; Zhang, C.; Cui, T.; Fang, X.-W.; et al. Conserved water use improves the yield performance of soybean (Glycine max (L.) Merr.) under drought. Agric. Water Manag. 2017, 179, 236–245. [Google Scholar] [CrossRef]
- Zhou, C.; Zhang, R.; Ning, X.; Zheng, Z. Spatial-Temporal Characteristics in Grain Production and Its Influencing Factors in the Huang-Huai-Hai Plain from 1995 to 2018. Int. J. Environ. Res. Public Health 2020, 17, 9193. [Google Scholar] [CrossRef] [PubMed]
- Wei, N.; Wang, N.; Zheng, Y.; Liu, H. Contribution of climate change and urbanization to the variation of extreme precipitation in the urban agglomerations over the Loess Plateau. Hydrol. Process. 2022, 36, e14489. [Google Scholar] [CrossRef]
- Li, H.; Zhang, D. Morphological characteristics and growth redundancy of spring wheat root system in semi-arid regions. Chin. J. Appl. Ecol. 1999, 10, 26–30. [Google Scholar]
- Passioura, J.B. Roots and drought resistance. Agric. Water Manag. 1983, 7, 265–280. [Google Scholar] [CrossRef]
- Richards, R.A.; Passioura, J.B. Seminal root morphology and water-use of wheat. 1. Environmental effects. Crop Sci. 1981, 21, 249–252. [Google Scholar] [CrossRef]
- Richards, R.A.; Passioura, J.B. Seminal root morphology and water use of wheat. II. Genetic variation. Crop Sci. 1981, 21, 253–255. [Google Scholar] [CrossRef]
- Richards, R.; Passioura, J. A breeding program to reduce the diameter of the major xylem vessel in the seminal roots of wheat and its effect on grain yield in rain-fed environments. Aust. J. Agric. Res. 1989, 40, 943–950. [Google Scholar] [CrossRef]
- Voss-Fels, K.P.; Snowdon, R.J.; Hickey, L.T. Designer roots for future crops. Trends Plant Sci. 2018, 23, 957–960. [Google Scholar] [CrossRef]
- Cossani, C.M.; Sadras, V.O. Symmetric response to competition in binary mixtures of cultivars associates with genetic gain in wheat yield. Evol. Appl. 2021, 14, 2064–2078. [Google Scholar] [CrossRef] [PubMed]
- Donald, C.M. The breeding of crop ideotypes. Euphytica 1968, 17, 385–403. [Google Scholar] [CrossRef]
- Brunel-Saldias, N.; Ferrio, J.P.; Elazab, A.; Orellana, M.; del Pozo, A. Root Architecture and Functional Traits of Spring Wheat Under Contrasting Water Regimes. Front. Plant Sci. 2020, 11, 581140. [Google Scholar] [CrossRef]
- Sadras, V.; Lawson, C. Nitrogen and water-use efficiency of Australian wheat varieties released between 1958 and 2007. Eur. J. Agron. 2013, 46, 34–41. [Google Scholar] [CrossRef]
- Prince, S.J.; Murphy, M.; Mutava, R.N.; Durnell, L.A.; Valliyodan, B.; Shannon, J.G.; Nguyen, H.T. Root xylem plasticity to improve water use and yield in water-stressed soybean. J. Exp. Bot. 2017, 68, 2027–2036. [Google Scholar] [CrossRef]
- Fonta, J.E.; Giri, J.; Vejchasarn, P.; Lynch, J.P.; Brown, K.M. Spatiotemporal responses of rice root architecture and anatomy to drought. Plant Soil 2022, 479, 443–464. [Google Scholar] [CrossRef]
- Cabello, J.V.; Chan, R.L. Arabidopsis and sunflower plants with increased xylem area show enhanced seed yield. Plant J. 2019, 99, 717–732. [Google Scholar] [CrossRef]
- Chen, D.G.; Zhou, X.Q.; Chen, K.; Chen, P.L.; Guo, J.; Liu, C.G.; Chen, Y.D. Fine-mapping and candidate gene analysis of a major locus controlling leaf thickness in rice (Oryza sativa L.). Mol. Breed. 2022, 42, 6. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wan, B.; Zhang, X. Plant ideotype at heading for super high-yielding rice in double cropping system in south China. Rice Sci. 2005, 12, 92–100. [Google Scholar]
- Sadras, V.; Lawson, C.; Montoro, A. Photosynthetic traits in Australian wheat varieties released between 1958 and 2007. Field Crop. Res. 2012, 134, 19–29. [Google Scholar] [CrossRef]
- Gaju, O.; Reynolds, M.P.; Sparkes, D.L.; Foulkes, M.J. Relationships between large-spike phenotype, grain number, and yield potential in spring wheat. Crop Sci. 2009, 49, 961–973. [Google Scholar] [CrossRef]
- Philipp, N.; Weichert, H.; Bohra, U.; Weschke, W.; Schulthess, A.W.; Weber, H. Grain number and grain yield distribution along the spike remain stable despite breeding for high yield in winter wheat. PLoS ONE 2018, 13, e0205452. [Google Scholar] [CrossRef] [PubMed]
- Shearman, V.J.; Sylvester-Bradley, R.; Scott, R.K.; Foulkes, M.J. Physiological Processes Associated with Wheat Yield Progress in the UK. Crop. Sci. 2005, 45, 175–185. [Google Scholar] [CrossRef]
- Quintero, A.; Molero, G.; Reynolds, M.P.; Calderini, D.F. Trade-off between grain weight and grain number in wheat depends on GxE interaction: A case study of an elite CIMMYT panel (CIMCOG). Eur. J. Agron. 2018, 92, 17–29. [Google Scholar] [CrossRef]
- Zhai, H.; Feng, Z.; Du, X.; Song, Y.; Liu, X.; Qi, Z.; Song, L.; Li, J.; Li, L.; Peng, H.; et al. A novel allele of TaGW2-A1 is located in a finely mapped QTL that increases grain weight but decreases grain number in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2018, 131, 539–553. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Cultivar | Year of Release | Days to Flower | Days to Mature | Source |
|---|---|---|---|---|---|
| 1 | Yumai49 | 2000 | 183 | 219 | Xiangyun Agricultural Technology Station |
| 2 | Zhengmai9023 | 2002 | 184 | 217 | Henan Academy of Agricultural Science |
| 3 | Luohan2 | 2003 | 184 | 222 | Luoyang Agricultural Science Research Institute |
| 4 | Zhoumai18 | 2005 | 184 | 221 | Zhoukou Academy of Agricultural Science |
| 5 | BainongAK58 | 2005 | 180 | 217 | Henan Institute of Science and Technology |
| 6 | Jimai22 | 2007 | 184 | 220 | Shandong Academy of Agricultural Science |
| 7 | Luohan12 | 2008 | 184 | 220 | Luoyang Agricultural Science Research Institute |
| 8 | Yunhan20410 | 2008 | 185 | 219 | Shanxi Academy of Agricultural Science |
| 9 | Luomai9 | 2008 | 185 | 220 | Luohe Academy of Agricultural Science |
| 10 | Shannong20 | 2010 | 183 | 219 | Shandong Agricultural University |
| 11 | Zhongmai175 | 2011 | 182 | 219 | Chinese Academy of agricultural science |
| 12 | Zhoumai26 | 2012 | 182 | 220 | Zhoukou Agricultural University |
| 13 | Henong7106 | 2012 | 183 | 222 | Hebei Agricultural University |
| 14 | Zhengmai101 | 2013 | 183 | 220 | Henan Academy of Agricultural Science |
| 15 | Bainong207 | 2013 | 184 | 221 | Henan Bainong Seed Industry Co. Ltd. |
| 16 | Yumai158 | 2014 | 185 | 221 | Luohe Academy of Agricultural Science |
| 17 | Zhengmai379 | 2016 | 184 | 221 | Henan Academy of Agricultural Science |
| 18 | Lunxuan99 | 2016 | 182 | 219 | Chinese Academy of agricultural science |
| 19 | Zhoumai32 | 2018 | 183 | 219 | Zhoukou Academy of Agricultural Science |
| 20 | Tongmai6 | 2019 | 183 | 221 | Center of Tongchuan Agricultural Technology Development |
| Anatomical Traits | Units | Descriptions |
|---|---|---|
| Root anatomical traits | ||
| Root diameter | μm | The whole root’s cross-sectional diameter |
| Cortex area | μm2 | The area between the exodermis and endodermis |
| Xylem vessel area | μm2 | Also named the xylem area, total vessel area |
| Xylem ratio | % | Root xylem vessel area/total root cross-sectional area ratio |
| Cortex-to-stele ratio | Root cortex area/total stele area ratio | |
| Cortex ratio | Root cortex area/total cross-sectional area ratio | |
| Stele ratio | Root stele area/total cross-sectional area ratio | |
| Root vessel density | Root vessel number/root stele area ratio | |
| Leaf anatomical traits | ||
| Vein density | mm−1 | Total number of veins per mm, also known as “vein length per leaf area” |
| Major vein area | μm2 | Includes leaf vascular bundle sheath, mechanical tissue, xylem and phloem |
| Leaf thickness | μm | The average leaf thickness at three positions |
| Leaf vein area ratio | The total leaf vein area to the total leaf cross-sectional area | |
| Leaf mesophyll area ratio | The total leaf mesophyll area to the total leaf cross-sectional area |
| Yield and Anatomical Traits | PC1 | PC2 |
|---|---|---|
| Yield | −0.42 | 0.67 |
| Root anatomical traits | ||
| Root diameter | 0.75 | 0.58 |
| Cortex area | 0.80 | 0.48 |
| Stele area | 0.25 | 0.90 |
| Xylem area | 0.01 | 0.92 |
| Root vessel density | −0.62 | −0.51 |
| Cortex ratio | 0.86 | −0.33 |
| Xylem ratio | −0.69 | 0.51 |
| Leaf anatomical traits | ||
| Leaf vein density | 0.04 | −0.28 |
| Major vein area | 0.63 | 0.08 |
| Leaf thickness | 0.22 | 0.71 |
| Leaf mesophyll area ratio | −0.58 | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, P.; Zhu, Y.-H.; Weiner, J.; Sun, Z.; Li, H.; Feng, T.; Li, F.-M. Reduced Root Cortical Tissue with an Increased Root Xylem Investment Is Associated with High Wheat Yields in Central China. Plants 2024, 13, 1075. https://doi.org/10.3390/plants13081075
Du P, Zhu Y-H, Weiner J, Sun Z, Li H, Feng T, Li F-M. Reduced Root Cortical Tissue with an Increased Root Xylem Investment Is Associated with High Wheat Yields in Central China. Plants. 2024; 13(8):1075. https://doi.org/10.3390/plants13081075
Chicago/Turabian StyleDu, Pengzhen, Yong-He Zhu, Jacob Weiner, Zhengli Sun, Huiquan Li, Tao Feng, and Feng-Min Li. 2024. "Reduced Root Cortical Tissue with an Increased Root Xylem Investment Is Associated with High Wheat Yields in Central China" Plants 13, no. 8: 1075. https://doi.org/10.3390/plants13081075
APA StyleDu, P., Zhu, Y.-H., Weiner, J., Sun, Z., Li, H., Feng, T., & Li, F.-M. (2024). Reduced Root Cortical Tissue with an Increased Root Xylem Investment Is Associated with High Wheat Yields in Central China. Plants, 13(8), 1075. https://doi.org/10.3390/plants13081075