
WRKY22 Transcription Factor from Iris laevigata Regulates Flowering Time and Resistance to Salt and Drought

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. The Treatment of I. laevigata under Salt and Drought Stress
2.3. Gene Sequence Identification and Phylogenetic Analysis
2.4. Gene Cloning and Sequence Analysis
2.5. Subcellular Localization
2.6. Plant Transformation
2.7. Determination of Flowering Phenotypes and Gene Expression
2.8. The Treatment of Transgenic N. tabacum Salt and Drought Stress
2.9. Photosynthetic Parameters in N. tabacum under Salt and Drought Stress
2.10. Other Physiological Measurements under Salt and Drought Stress N. tabacum Plants
2.11. Reverse Transcription PCR (RT-PCR) and RT-qPCR
2.12. Statistical Analysis
3. Results
3.1. Identification of WRKY Genes in I. laevigata
3.2. Cloning of IlWRKY22 and Subcellular Localization
3.3. Expression of IlWRKY22 in I. laevigata
3.4. Overexpressing IlWRKY22 in Both A. thaliana and N. tabacum
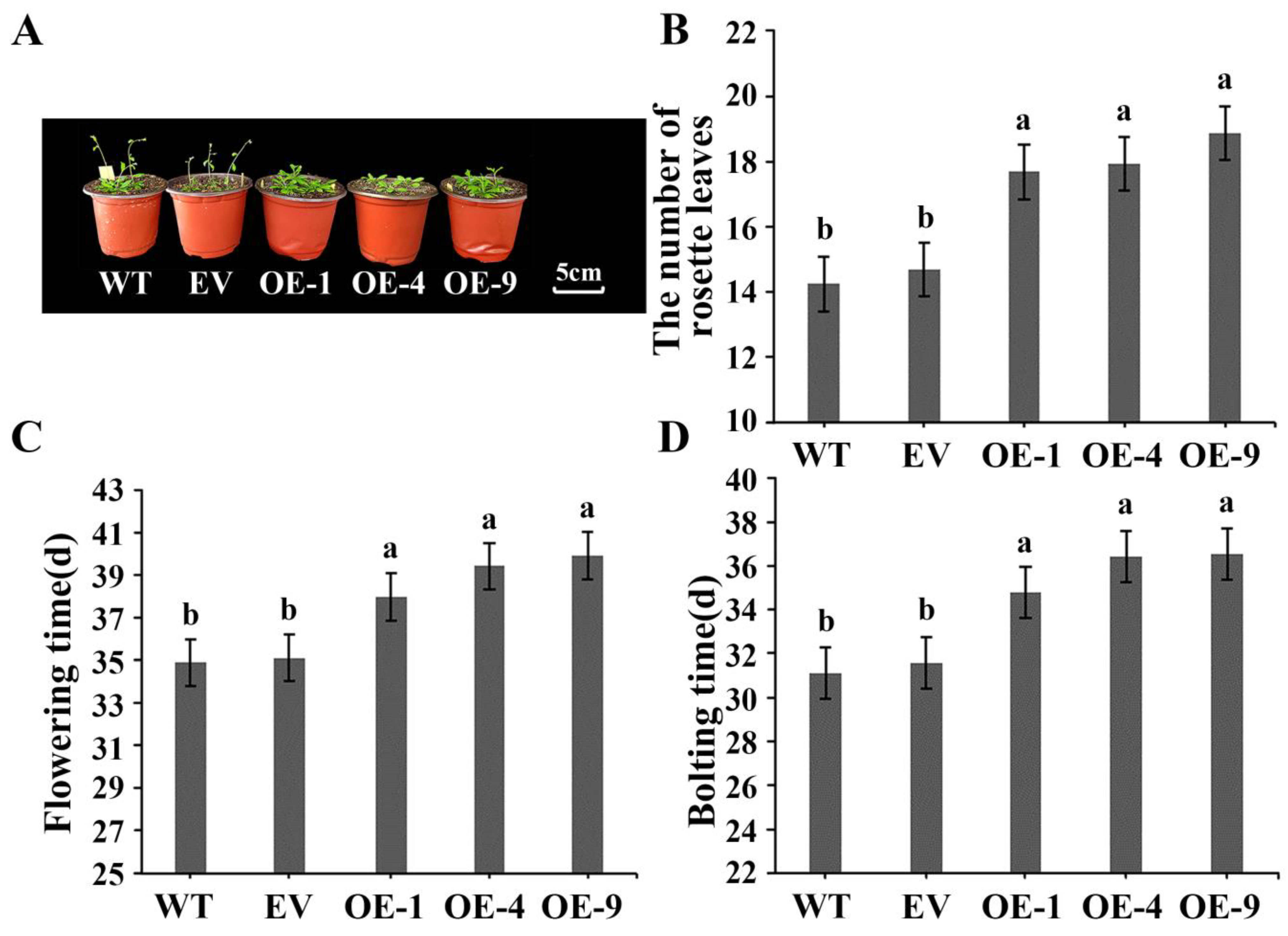
3.5. Overexpressing IlWRKY22 Delays Flowering in A. thaliana and N. tabacum
3.6. Overexpression of IlWRKY22 Modulates the Expression of Flowering Time Genes
3.7. Overexpressing IlWRKY22 in N. tabacum Enhances Resistance to Salt and Drought Stress
3.8. Impact of Overexpressing IlWRKY22 on Photosynthesis under Salt and Drought Stress
3.9. IlWRKY22 Promotes the Response of N. tabacum to Salt and Drought Stress
3.10. The IlWRKY22 on Expression Levels of Stress-Related Genes NtCAT, NtHAK1, NtPMA4, NtPOD, NtSOD, and NtSOS1 under Salt and Drought Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wellmer, F.; Riechmann, J.L. Gene networks controlling the initiation of flower development. Trends Genet. 2010, 26, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.; Bahadur, S.; Xia, W. An overview of floral regulatory genes in annual and perennial plants. Gene 2023, 885, 147699. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Chen, T.; Zeng, X. Genetic and epigenetic understanding of the seasonal timing of flowering. Plant Commun. 2020, 1, 100008. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, K.; Singh, A.; Berger, J.; Bishop-von, W.E.; Kahraman, A.; Aydogan, A.; Cook, D.; Nuzhdin, S.; Samsonova, M. Non-linear regression models for time to flowering in wild chickpea combine genetic and climatic factors. BMC Plant Biol. 2019, 19, 94. [Google Scholar] [CrossRef]
- Wang, F.; Li, S.; Kong, F.; Lin, X.; Lu, S. Altered regulation of flowering expands growth ranges and maximizes yields in major crops. Front. Plant Sci. 2023, 14, 1094411. [Google Scholar] [CrossRef]
- Shah, S.; Weinholdt, C.; Jedrusik, N.; Molina, C.; Zou, J.; Grosse, I.; Schiessl, S.; Jung, C.; Emrani, N. Whole-transcriptome analysis reveals genetic factors underlying flowering time regulation in rapeseed (Brassica napus L.). Plant Cell Environ. 2018, 41, 1935–1947. [Google Scholar] [CrossRef]
- Liu, Y.P.; Yang, J.; Yang, M.F. Pathways of flowering regulation in plants. Chin. J. Biol. 2015, 31, 1553–15666. [Google Scholar]
- Liang, H.M.; Xia, Y.; Wang, T.M. Research Progress on Genetic Engineering of Plant Cold Resistance, Drought Resistance and Salt Tolerance. Acta Pratac. Sin. 2003, 3, 1–7. [Google Scholar]
- Sun, Y.; Zhou, J.; Guo, J. Advances in the knowledge of adaptive mechanisms mediating abiotic stress responses in Camellia sinensis. Front. Biosci. 2021, 26, 1714–1722. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Biswas, T.; Islam, M.A.; Haque, T. Exogenously applied moringa leaf extracts and mixed fertilizers in soil to improve growth and yield of tomato (Lycopersicon esculentum Mill.). Sustain. Food Agric. 2020, 1, 42–47. [Google Scholar] [CrossRef]
- Luo, X.; Li, C.; He, X.; Zhang, X.; Zhu, L. ABA signaling is negatively regulated by GbWRKY1 through JAZ1 and ABI1 to affect salt and drought tolerance. Plant Cell Rep. 2020, 39, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Xiao, L.Y.; Sun, G.W.; Liu, H.C.; Song, S.W.; Chen, R.Y. Functional analysis of transcription factor BCWRKY22 in low temperature-induced bolting and flowering of Brassica campestris. Mol. Breed. 2020, 18, 3862–3870. [Google Scholar]
- Li, J.; Han, G.; Sun, C.; Sui, N. Research advances of MYB transcription factors in plant stress resistance and breeding. Plant Signal. Behav. 2019, 14, 1613131. [Google Scholar] [CrossRef] [PubMed]
- Golldack, D.; Luking, I.; Yang, O. Plant tolerance to drought and salinity: Stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant Cell Rep. 2011, 30, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Fedoroff, N.V.; Battisti, D.S.; Beachy, R.N.; Cooper, P.J.; Fischhoff, D.A.; Hodges, C.N.; Knauf, V.C.; Lobell, D.; Mazur, B.J.; Molden, D.; et al. Radically rethinking agriculture for the 21st century. Science 2010, 327, 833–834. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Huang, G.; Zhu, Y. Sustainable plant disease control: Biotic information flow and behavior manipulation. Sci. China-Life Sci. 2019, 62, 1710–1713. [Google Scholar] [CrossRef]
- Ma, J.J. Cloning and Preliminary Function Analysis of Hawrky9/14/22/30/79 Genes in Helianthus annuus. Master’s Thesis, Harbin Normal University, Harbin, China, 2022; p. 001596. [Google Scholar]
- Chen, Y.P.; Lin, J.H.; Liu, Z.Q.; He, S.L.; She, W.Q.; Chen, G.X. Cloning and functional identification of promoter of PsWRKY22 gene in Prunus salicina. Mol. Breed. 2021, 19, 2837–2845. [Google Scholar]
- Khoso, M.A.; Hussain, A.; Ritonga, F.N.; Ali, Q.; Channa, M.M.; Alshegaihi, R.M.; Meng, Q.; Ali, M.; Zaman, W.; Brohi, R.D.; et al. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stresses in plants. Front. Plant Sci. 2022, 13, 1039329. [Google Scholar] [CrossRef]
- Phukan, U.J.; Jeena, G.S.; Shukla, R.K. WRKY transcription factors: Molecular regulation and stress responses in plants. Front. Plant Sci. 2016, 7, 760. [Google Scholar] [CrossRef]
- Agarwal, P.; Reddy, M.P.; Chikara, J. WRKY: Its structure, evolutionary relationship, DNA-binding selectivity, role in stress tolerance and development of plants. Mol. Biol. Rep. 2011, 38, 3883–3896. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Ma, S.; Ye, N.; Jiang, M.; Cao, J.; Zhang, J. WRKY transcription factors in plant responses to stresses. J. Integr. Plant Biol. 2017, 59, 86–101. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, C.; Wang, H.; Guo, Z. WRKY transcription factors: Evolution, binding, and action. Phytopathol. Res. 2019, 1, 13. [Google Scholar] [CrossRef]
- Bakshi, M.; Oelmuller, R. WRKY transcription factors: Jack of many trades in plants. Plant Signal. Behav. 2014, 9, e27700. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Li, F.; Shi, G.; Wang, L.; Wang, L.; Fan, L. Identification of MADS-Box transcription factors in Iris laevigata and functional assessment of IlSEP3 and IlSVP during flowering. Int. J. Mol. Sci. 2022, 23, 9950. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, S.; Shi, G.F.; Yan, L.; Lv, R.T.; Ma, Z.; Wang, L. Comparative analysis of R2R3-MYB transcription factors in the flower of Iris laevigata identifies a novel gene regulating tobacco cold tolerance. Plant Biol. 2022, 24, 1066–1075. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome editing and precision plant breeding in agriculture. Annu. Rev. Plant Biol. 2019, 70, 667–697. [Google Scholar] [CrossRef]
- Kourelis, J.; van der Hoorn, R.; Sueldo, D.J. Decoy engineering: The next step in resistance breeding. Trends Plant Sci. 2016, 21, 371–373. [Google Scholar] [CrossRef] [PubMed]
- Tribhuvan, K.U.; Mishra, T.; Kaur, S. Identification and characterization of novel drought-responsive lncRNAs in stone apple (Aegle marmelos L.) through whole-transcriptome analysis. Curr. Plant Biol. 2024, 38, 100336. [Google Scholar] [CrossRef]
- Zeng, J.; Wang, Y.; Wu, G. Comparative Transcriptome Analysis Reveals the Genes and Pathways Related to Wheat Root Hair Length. Int. J. Mol. Sci. 2024, 25, 2069. [Google Scholar] [CrossRef]
- Wang, Y.; Xun, H.; Wang, L. Identification of Key Genes Associated with 1,2,6-Tri-O-galloyl-beta-D-glucopyranose Accumulation in Camellia sinensis Based on Transcriptome Sequencing. Foods 2024, 13, 495. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, L. The WRKY transcription factor superfamily: Its origin in eukaryotes and expansion in plants. BMC Evol. Biol. 2005, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Ramos, R.N.; Zhang, N.; Lauff, D.B. Loss-of-function mutations in WRKY22 and WRKY25 impair stomatal-mediated immunity and PTI and ETI responses against Pseudomonas syringae pv. tomato. Plant Mol. Biol. 2023, 112, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.X.; Cao, H.X. Expression characteristics of WRKY transcription factor gene in oil palm under low temperature stress. J. South. Agric. 2018, 49, 1490–1497. [Google Scholar]
- Srikanth, A.; Schmid, M. Regulation of flowering time: All roads lead to Rome. Cell. Mol. Life Sci. 2011, 68, 2013–2037. [Google Scholar] [CrossRef] [PubMed]
- Collings, D.A. Subcellular localization of transiently expressed fluorescent fusion proteins. Methods Mol. Biol. 2013, 1069, 227–258. [Google Scholar] [PubMed]
- Wang, H.; Liu, Z.; Xie, J.; Li, J.; Zhang, J.; Yu, J.; Hu, L.; Zhang, G. The CaALAD gene from pepper (Capsicum annuum L.) confers chilling stress tolerance in transgenic Arabidopsis plants. Front. Plant Sci. 2022, 13, 884990. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, S.; Guan, C.; Kong, X.; Wang, Y.; Cui, Y.; Liu, B.; Zhou, Y.; Zhang, Y. Overexpressing the NAC transcription factor LpNAC13 from Lilium pumilum in tobacco negatively regulates the drought response and positively regulates the salt response. Plant Physiol. Biochem. 2020, 149, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, L.; Zhang, H.; Chen, L.; Yu, D. ERF1 delays flowering through direct inhibition of Flowering locus T expression in Arabidopsis. J. Integr. Plant Biol. 2021, 63, 1712–1723. [Google Scholar] [CrossRef]
- Fukazawa, J.; Ohashi, Y.; Takahashi, R.; Nakai, K.; Takahashi, Y. DELLA degradation by gibberellin promotes flowering via GAF1-TPR-dependent repression of floral repressors in Arabidopsis. Plant Cell 2021, 33, 2258–2272. [Google Scholar] [CrossRef]
- Sanagi, M.; Aoyama, S.; Kubo, A.; Lu, Y.; Sato, Y.; Ito, S.; Abe, M.; Mitsuda, N.; Ohme-Takagi, M.; Kiba, T.; et al. Low nitrogen conditions accelerate flowering by modulating the phosphorylation state of FLOWERING BHLH 4 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2022942118. [Google Scholar] [CrossRef] [PubMed]
- Tayengwa, R.; Sharma, K.P.; Pierce, C.F.; Werner, B.E.; Neff, M.M. Overexpression of AtAHL20 causes delayed flowering in Arabidopsis via repression of FT expression. BMC Plant Biol. 2020, 20, 559. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Y.; Liu, M.S.; Li, J.R.; Guan, C.M.; Zhang, X.S. The wheat TaGI1, involved in photoperiodic flowering, encodes an Arabidopsis GI ortholog. Plant Mol. Biol. 2005, 58, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Anwar, T.; Shehzadi, A.; Qureshi, H.; Shah, M.N.; Danish, S.; Salmen, S.H.; Ansari, M.J. Alleviation of cadmium and drought stress in wheat by improving growth and chlorophyll contents amended with GA3 enriched deashed biochar. Sci. Rep. 2023, 13, 18503. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Zhu, G.; Zhou, G.; Liu, J.; Younas, M.U.; Zhu, Y. Melatonin role in plant growth and physiology under abiotic stress. Int. J. Mol. Sci. 2023, 24, 8759. [Google Scholar] [CrossRef] [PubMed]
- Waadt, R.; Seller, C.A.; Hsu, P.K. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef]
- Gao, Y.-F.; Liu, J.-K.; Yang, F.-M.; Zhang, G.-Y.; Wang, D.; Zhang, L.; Ou, Y.-B.; Yao, Y.-A. The WRKY transcription factor WRKY8 promotes resistance to pathogen infection and mediates drought and salt stress tolerance in Solanum lycopersicum. Physiol. Plant. 2020, 168, 98–117. [Google Scholar] [CrossRef] [PubMed]
- Ampofo, J.O.; Ngadi, M. Stimulation of the phenylpropanoid pathway and antioxidant capacities by biotic and abiotic elicitation strategies in common bean (Phaseolus vulgaris) sprouts. Process Biochem. 2021, 100, 98–106. [Google Scholar] [CrossRef]
- Chen, H.; Li, X.; Li, F.; Li, D.; Dong, Y.; Fan, Y. Bioinformatics analysis of wrky family genes in Erianthus fulvus Ness. Genes 2022, 13, 2102. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z. Identification of WRKY Transcription Factor Family in Xanthoceras sorbifolia and Analysis of Response Patterns to Abiotic Stresses. Master’s Thesis, Northeast Forestry University, Harbin, China, 2020; p. 000390. [Google Scholar]
- Liu, Y.X. Identification and Expression Analysis of WRKY Gene Family in Tomato. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2020; p. 000900. [Google Scholar]
- Zhou, X.; Jiang, Y.; Yu, D. WRKY22 transcription factor mediates dark-induced leaf senescence in Arabidopsis. Mol. Cells 2011, 31, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, X.; Huang, X.; Su, W.; Hao, Y.; Liu, H.; Chen, R.; Song, S. BcSOC1 Promotes Bolting and Stem Elongation in Flowering Chinese Cabbage. Int. J. Mol. Sci. 2022, 23, 3459. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, W.; Xu, Z.; Chen, M.; Yu, D. Functions of WRKYs in plant growth and development. Trends Plant Sci. 2023, 28, 630–645. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.; Du, M.; Long, J.; Xie, Y.; Zhang, J.; Xu, L.; He, Y.; Li, Q.; Chen, S.; Zou, X. Transcription factor WRKY22 regulates canker susceptibility in sweet orange (Citrus sinensis Osbeck) by enhancing cell enlargement and CsLOB1 expression. Hortic. Res. 2021, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Li, T.; Cao, X.; Zhang, D.; Teng, N. Lily WRKY factor LlWRKY22 promotes thermotolerance through autoactivation and activation of Lldreb2b. Hortic. Res. 2022, 9, uhac186. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S. Identification of WRKY gene family in chestnut and analysis of its expression under drought stress. North China J. Agron. 2024, 39, 72–82. [Google Scholar]
- Balti, I.; Benny, J.; Perrone, A.; Caruso, T.; Abdallah, D.; Salhi-Hannachi, A.; Martinelli, F. Identification of conserved genes linked to responses to abiotic stresses in leaves among different plant species. Funct. Plant Biol. 2020, 48, 54–71. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, S.; Sun, X.; Khan, N.U.; Zhong, Q.; Zhang, Z.; Zhang, H.; Ming, F.; Li, Z.; Li, J. Variations in OsSPL10 confer drought tolerance by directly regulating OsNAC2 expression and ROS production in rice. J. Integr. Plant Biol. 2023, 65, 918–933. [Google Scholar] [CrossRef] [PubMed]
- Anuradha, M.; Sivaraju, K.; Krishnamurthy, V. Effect of waterlogging on physiological characteristics, yield and quality of flue-cured tobacco. Indian J. Plant Physiol. 2013, 18, 67–70. [Google Scholar] [CrossRef]
- Elshoky, H.A.; Yotsova, E.; Farghali, M.A.; Farroh, K.Y.; El-Sayed, K.; Elzorkany, H.E.; Rashkov, G.; Dobrikova, A.; Borisova, P.; Stefanov, M.; et al. Impact of foliar spray of zinc oxide nanoparticles on the photosynthesis of Pisum sativum L. under salt stress. Plant Physiol. Biochem. 2021, 167, 607–618. [Google Scholar] [CrossRef]
- Razi, K.; Muneer, S. Drought stress-induced physiological mechanisms, signaling pathways and molecular response of chloroplasts in common vegetable crops. Crit. Rev. Biotechnol. 2021, 41, 669–691. [Google Scholar] [CrossRef] [PubMed]
- Zahra, N.; Al, H.M.; Hafeez, M.B.; Rehman, A.; Wahid, A.; Siddique, K.; Farooq, M. Regulation of photosynthesis under salt stress and associated tolerance mechanisms. Plant Physiol. Biochem. 2022, 178, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Skopelitis, D.S.; Paranychianakis, N.V.; Paschalidis, K.A.; Pliakonis, E.D.; Delis, I.D.; Yakoumakis, D.I.; Kouvarakis, A.; Papadakis, A.K.; Stephanou, E.G.; Roubelakis-Angelakis, K.A. Abiotic stress generates ROS that signal expression of anionic glutamate dehydrogenases to form glutamate for proline synthesis in tobacco and grapevine. Plant Cell 2006, 18, 2767–2781. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Riadh, K.; Gopi, R.; Manivannan, P.; Inès, J.; Al-Juburi, H.J.; Chang-Xing, Z.; Hong-Bo, S.; Panneerselvam, R. Antioxidant defense responses: Physiological plasticity in higher plants under abiotic constraints. Acta Physiol. Plant 2009, 31, 427–436. [Google Scholar] [CrossRef]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Moriau, L.; Bogaerts, P.; Jonniaux, J.L.; Boutry, M. Identification and characterization of a second plasma membrane H (+)-ATPase gene subfamily in Nicotiana plumbaginifolia. Plant Mol. Biol. 1993, 21, 955–963. [Google Scholar] [CrossRef]
- Qin, L.J.; Song, L.L.; Zhao, D.; Zhao, D.G. Overexpression of tobacco high-affinity potassium High affinity kalium transporter1 gene (NtHAK1) increased tobacco salt stress ability. J. Agric. Biotechnol. 2015, 23, 1576–1587. [Google Scholar]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, L.; Niu, Z.; Shi, G.; Song, Z.; Yang, Q.; Zhou, S.; Wang, L. WRKY22 Transcription Factor from Iris laevigata Regulates Flowering Time and Resistance to Salt and Drought. Plants 2024, 13, 1191. https://doi.org/10.3390/plants13091191
Fan L, Niu Z, Shi G, Song Z, Yang Q, Zhou S, Wang L. WRKY22 Transcription Factor from Iris laevigata Regulates Flowering Time and Resistance to Salt and Drought. Plants. 2024; 13(9):1191. https://doi.org/10.3390/plants13091191
Chicago/Turabian StyleFan, Lijuan, Zhaoqian Niu, Gongfa Shi, Ziyi Song, Qianqian Yang, Sheng Zhou, and Ling Wang. 2024. "WRKY22 Transcription Factor from Iris laevigata Regulates Flowering Time and Resistance to Salt and Drought" Plants 13, no. 9: 1191. https://doi.org/10.3390/plants13091191
APA StyleFan, L., Niu, Z., Shi, G., Song, Z., Yang, Q., Zhou, S., & Wang, L. (2024). WRKY22 Transcription Factor from Iris laevigata Regulates Flowering Time and Resistance to Salt and Drought. Plants, 13(9), 1191. https://doi.org/10.3390/plants13091191