The Response of the Functional Traits of Phragmites australis and Bolboschoenus planiculmis to Water and Saline–Alkaline Stresses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Relationships Between Key Functional Traits and Other Functional Traits in P. australis and B. planiculmis
2.2. Response of Functional Traits of P. australis and B. planiculmis to Saline–Alkaline Concentration
2.3. Water Level Induced Changes in the Functional Traits of P. australis and B. planiculmis in Response to Saline–Alkaline Concentration
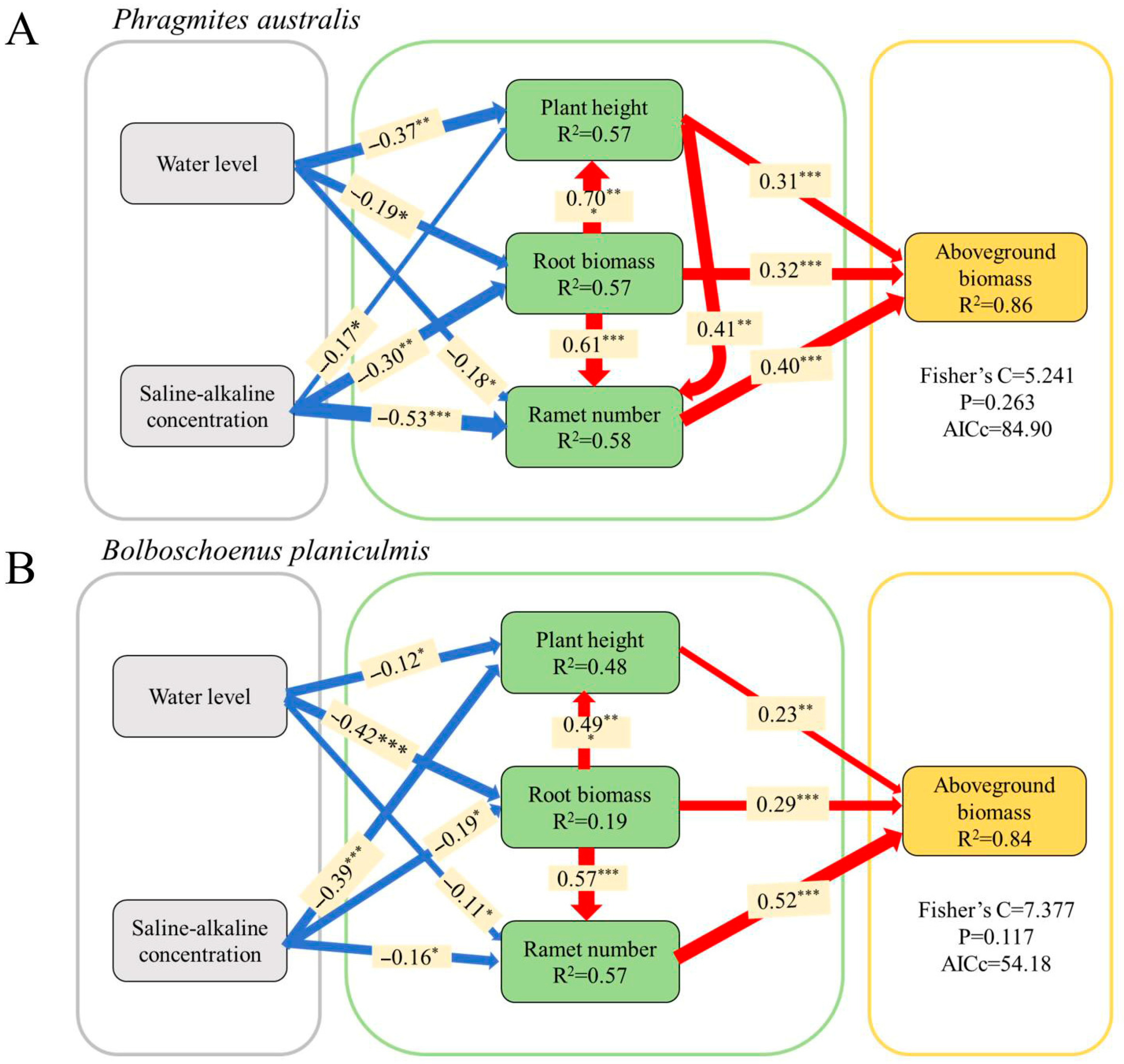
2.4. Response of Key Functional Traits of P. australis and B. planiculmis to Environmental Stresses and Their Ability to Predict Plant Production Performance
3. Discussion
3.1. Response of Functional Traits to Saline–Alkaline Stress
3.2. Response of Functional Traits to the Interactions Between Water Level and Saline–Alkaline Stress
3.3. Predictive Ability of Key Traits for Plant Performance
4. Materials and Methods
4.1. Study Area
4.2. Study Species
4.3. Sample Collection and Planting
4.4. Experimental Design
4.5. Functional Trait Measurements
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cañedo-Argüelles, M.; Bundschuh, M.; Gutiérrez-Cánovas, C.; Kefford, B.J.; Prat, N.; Trobajo, R. Effects of repeated salt pulses on ecosystem structure and functions in a stream mesocosm. Sci. Total Environ. 2014, 476, 634–642. [Google Scholar] [CrossRef]
- Garssen, A.G.; Baattrup-Pedersen, A.; Voesenek, L.; Verhoeven, J.T.A.; Soons, M.B. Riparian plant community responses to increased flooding: A meta-analysis. Glob. Change Biol. 2015, 21, 2881–2890. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, Z.; Bachofen, C.; Lou, Y.; Jiang, M.; Tang, X. The effect of saline-alkaline and water stresses on water use efficiency and standing biomass of Phragmites australis and Bolboschoenus planiculmis. Sci. Total Environ. 2018, 644, 207–216. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M.; Zak, M.; Carretero, E.M.; Araníbar, J. Plant functional traits, ecosystem structure and land-use history along a climatic gradient in central-western Argentina. J. Veg. Sci. 1999, 10, 651–660. [Google Scholar] [CrossRef]
- Ding, S.; Yu, X.; Zhang, J.; Yin, Z.; Zou, Y.; Wang, G. Bioconcentration and translocation of elements regulate plant responses to water-salt conditions in saline-alkaline wetlands. Environ. Exp. Bot. 2021, 183, 104360. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Anee, T.I.; Parvin, K.; Nahar, K.; Al Mahmud, J. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef]
- Elmore, A.J.; Manning, S.J.; Mustard, J.F.; Craine, J.M. Decline in alkali meadow vegetation cover in California: The effects of groundwater extraction and drought. J. Appl. Ecol. 2006, 43, 770–779. [Google Scholar] [CrossRef]
- Rosas, H.L.; Gonzalez, V.E.E.; Moreno-Casasola, P. Decreases in mangrove productivity and marsh die-off due to temporary increase in salinity, a case in Mexico. Hydrobiologia 2023, 850, 4497–4514. [Google Scholar] [CrossRef]
- Rogel, J.A.; Ariza, F.A.; Silla, R.O. Soil salinity and moisture gradients and plant zonation in Mediterranean salt marshes of Southeast Spain. Wetlands 2000, 20, 357–372. [Google Scholar] [CrossRef]
- An, Y.; Gao, Y.; Tong, S.; Liu, B. Morphological and physiological traits related to the response and adaption of Bolboschoenus planiculmis seedlings grown under salt-alkaline stress conditions. Front. Plant Sci. 2021, 12, 567782. [Google Scholar] [CrossRef]
- Lou, Y.; Pan, Y.; Gao, C.; Jiang, M.; Lu, X.; Xu, Y. Response of plant height, species richness and aboveground biomass to flooding gradient along vegetation zones in floodplain wetlands, northeast China. PLoS ONE 2016, 11, 0153972. [Google Scholar] [CrossRef]
- Voesenek, L.; Rijnders, J.; Peeters, A.J.M.; Van de Steeg, H.M.V.; De Kroon, H. Plant hormones regulate fast shoot elongation under water: From genes to communities. Ecology 2004, 85, 16–27. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, D.; Qi, Q.; Tong, S.; Wang, X.; An, Y. Flooding effects on population and growth characteristics of Bolboschoenus planiculmis in Momoge wetland, northeast China. Ecol. Indic. 2022, 137, 108730. [Google Scholar] [CrossRef]
- Sasidharan, R.; Hartman, S.; Liu, Z.; Martopawiro, S.; Sajeev, N.; van Veen, H. Signal Dynamics and interactions during flooding stress. Plant Physiol. 2018, 176, 1106–1117. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Voesenek, L. Flooding stress: Acclimations and genetic diversity. Annu. Rev. Plant Biol. 2008, 59, 313–339. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Voesenek, L. Life in the balance: A signaling network controlling survival of flooding. Curr. Opin. Plant Biol. 2010, 13, 489–494. [Google Scholar] [CrossRef]
- Ding, Z.; Liu, Y.; Lou, Y.; Jiang, M.; Li, H.; Lü, X. How soil ion stress and type influence the flooding adaptive strategies of Phragmites australis and Bolboschoenus planiculmis in temperate saline-alkaline wetlands? Sci. Total Environ. 2021, 771, 144654. [Google Scholar] [CrossRef]
- Shan, L.; Song, C.; Zhang, X.; Wang, X.; Luan, Z. Responses of above-ground biomass, plant diversity, and dominant species to habitat change in a freshwater wetland of northeast China. Russ. J. Ecol. 2020, 51, 57–63. [Google Scholar] [CrossRef]
- Dai, X.; Yu, Z.; Yang, G.; Wan, R. Role of flooding patterns in the biomass production of vegetation in a typical herbaceous wetland, poyang lake wetland, China. Front. Plant Sci. 2020, 11, 521358. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, M.; Tong, S.; Qi, Q.; Wang, X.; Lu, X. Growth and physiological responses of Carex schmidtii to water-level fluctuation. Hydrobiologia 2020, 847, 967–981. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, N.; Luo, L.; Li, X.; Ren, C.; Song, K. Shrinkage and fragmentation of marshes in the West Songnen Plain, China, from 1954 to 2008 and its possible causes. Int. J. Appl. Earth Obs. Geoinf. 2011, 13, 477–486. [Google Scholar] [CrossRef]
- Brix, H.; Cízková, H. Introduction: Phragmites-dominated wetlands, their functions and sustainable use. Aquat. Bot. 2001, 69, 87–88. [Google Scholar] [CrossRef]
- Cardoso, P.; Stoev, P.; Georgiev, T.; Senderov, V.; Penev, L. Species conservation profiles compliant with the iucn red list of threatened species. Biodiver. Data J. 2016, 4, 10356. [Google Scholar] [CrossRef]
- Cuda, J.; Skálová, H.; Meyerson, L.A.; Pysek, P. Regeneration of Phragmites australis from rhizome and culm fragments: An experimental test of environmental effects, population origin and invasion status. Preslia 2021, 93, 237–254. [Google Scholar] [CrossRef]
- Guo, C.; Ma, L.; Yuan, S.; Wang, R. Morphological, physiological and anatomical traits of plant functional types in temperate grasslands along a large-scale aridity gradient in northeastern China. Sci. Rep. 2017, 7, 40900. [Google Scholar] [CrossRef]
- Chai, Y.; Yue, M.; Wang, M.; Xu, J.; Liu, X.; Zhang, R. Plant functional traits suggest a change in novel ecological strategies for dominant species in the stages of forest succession. Oecologia 2016, 180, 771–783. [Google Scholar] [CrossRef]
- van der Plas, F.; Schröder-Georgi, T.; Weigelt, A.; Barry, K.; Meyer, S.; Alzate, A. Plant traits alone are poor predictors of ecosystem properties and long-term ecosystem functioning. Nat. Ecol. Evol. 2020, 4, 1602–1611. [Google Scholar] [CrossRef]
- Ostermann, T.S.; Kleyer, M.; Heuner, M.; Fuchs, E.; Temmerman, S.; Schoutens, K. Hydrodynamics affect plant traits in estuarine ecotones with impact on carbon sequestration potentials. Estuar. Coast. Shelf Sci. 2021, 259, 107464. [Google Scholar] [CrossRef]
- Li, X.; Wen, B.; Yang, F.; Hartley, A.; Li, X. Effects of alternate flooding-drought conditions on degenerated Phragmites australis salt marsh in Northeast China. Restor. Ecol. 2017, 25, 810–819. [Google Scholar] [CrossRef]
- Xu, X.; Liu, H.; Liu, Y.; Zhou, C.; Pan, L.; Fang, C. Human eutrophication drives biogeographic salt marsh productivity patterns in China. Ecol. Appl. 2020, 30, 2045. [Google Scholar] [CrossRef]
- Li, Q.; Wen, J.; Zhao, C.; Zhao, L.; Ke, D. The relationship between the main leaf traits and photosynthetic physiological characteristics of Phragmites australis under different habitats of a salt marsh in Qinwangchuan, China. Aob Plants 2022, 14, plac054. [Google Scholar] [CrossRef]
- Wang, C.; Lu, M.; Yang, B.; Yang, Q.; Zhang, X.; Hara, T. Effects of environmental gradients on the performances of four dominant plants in a Chinese saltmarsh: Implications for plant zonation. Ecol. Res. 2010, 25, 347–358. [Google Scholar] [CrossRef]
- Guo, X.; Yu, T.; Li, M.; Guo, W. The effects of salt and rainfall pattern on morphological and photosynthetic characteristics of Phragmites australis (Poaceae). J. Torrey Bot. Soc. 2018, 145, 212–224. [Google Scholar] [CrossRef]
- Song, H.; Jespersen, E.; Guo, X.; Du, N.; Xie, L.; Pei, L. Differences in relative air humidity affect responses to soil salinity in freshwater and salt marsh populations of the dominant grass species Phragmites australis. Hydrobiologia 2021, 848, 3353–3369. [Google Scholar] [CrossRef]
- Jespersen, E.; Kirk, G.H.; Brix, H.; Eller, F.; Sorrell, B.K. Shade and salinity responses of two dominant coastal wetland grasses: Implications for light competition at the transition zone. Ann. Bot. 2021, 128, 469–480. [Google Scholar] [CrossRef]
- Yang, C.; Chong, J.; Li, C.; Kim, C.; Shi, D.; Wang, D. Osmotic adjustment and ion balance traits of an alkali resistant halophyte Kochia sieversiana during adaptation to salt and alkali conditions. Plant Soil 2007, 294, 263–276. [Google Scholar] [CrossRef]
- Sheng, W.; Liu, L.; Wu, Y.; Yin, M.; Yu, Q.; Guo, X. Exploring salt tolerance and indicator traits across four temperate lineages of the common wetland plant, Phragmites australis. Sci. Total Environ. 2024, 912, 169100. [Google Scholar] [CrossRef]
- Grewell, B.J.; Gallego-Tévar, B.; Gillard, M.B.; Futrell, C.J.; Reicholf, R.; Castillo, J.M. Salinity and inundation effects on Iris pseudacorus: Implications for tidal wetland invasion with sea level rise. Plant Soil 2021, 466, 275–291. [Google Scholar] [CrossRef]
- Sun, X.; Chen, Y.; Zhuo, N.; Cui, Y.; Luo, F.; Zhang, M. Effects of salinity and concomitant species on growth of Phragmites australis populations at different levels of genetic diversity. Sci. Total Environ. 2021, 780, 146516. [Google Scholar] [CrossRef]
- Song, H.; Guo, X.; Yang, J.; Liu, L.; Li, M.; Wang, J. Phenotypic plasticity variations in Phragmites australis under different plant-plant interactions influenced by salinity. J. Plant Ecol. 2024, 17, 1752–9921. [Google Scholar] [CrossRef]
- Assaeed, A.M.; Dar, B.A.; Al-Doss, A.A.; Al-Rowaily, S.L.; Malik, J.A.; Abd-ElGawad, A.M. Phenotypic plasticity strategy of Aeluropus lagopoides grass in response to heterogenous saline habitats. Biology 2023, 12, 553. [Google Scholar] [CrossRef]
- Zhang, D.; Tong, S.; Qi, Q.; Zhang, M.; An, Y.; Wang, X. Effects of drought and re-flooding on growth and photosynthesis of Carex schmidtii Meinsh: Implication for tussock restoration. Ecol. Indic. 2019, 103, 134–144. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, R.; Zhu, H.; Cheng, X.; Shutes, B.; Yan, B. Seed germination and early seedling growth of six wetland plant species in saline-alkaline environment. Int. J. Phytoremediation 2020, 22, 1185–1194. [Google Scholar] [CrossRef]
- Reich, P.B.; Tjoelker, M.G.; Pregitzer, K.S.; Wright, I.J.; Oleksyn, J.; Machado, J.L. Scaling of respiration to nitrogen in leaves, stems and roots of higher land plants. Ecol. Lett. 2008, 11, 793–801. [Google Scholar] [CrossRef]
- De Battisti, D.; Fowler, M.S.; Jenkins, S.R.; Skov, M.W.; Rossi, M.; Bouma, T. Intraspecific root trait variability along environmental gradients affects salt marsh resistance to lateral erosion. Front. Ecol. Evol. 2019, 7, 150. [Google Scholar] [CrossRef]
- Luo, F.; Matsubara, S.; Chen, Y.; Wei, G.; Dong, B.; Zhang, M. Consecutive submergence and de-submergence both impede growth of a riparian plant during water level fluctuations with different frequencies. Environ. Exp. Bot. 2018, 155, 641–649. [Google Scholar] [CrossRef]
- An, Y.; Gao, Y.; Zhang, Y.; Tong, S.; Liu, X. Early establishment of Suaeda salsa population as affected by soil moisture and salinity: Implications for pioneer species introduction in saline-sodic wetlands in Songnen Plain, China. Ecol. Indic. 2019, 107, 105654. [Google Scholar] [CrossRef]
- Pennings, S.C.; Grant, M.B.; Bertness, M.D. Plant zonation in low-latitude salt marshes: Disentangling the roles of flooding, salinity and competition. J. Ecol. 2005, 93, 159–167. [Google Scholar] [CrossRef]
- Ziegler, V.H.; Ploschuk, E.; Weibel, A.; Insausti, P. Short-term responses to flooding stress of three Prunus rootstocks. Sci. Hortic. 2017, 224, 135–141. [Google Scholar] [CrossRef]
- De Battisti, D.; Fowler, M.S.; Jenkins, S.R.; Skov, M.W.; Bouma, T.J.; Neyland, P.J. Multiple trait dimensions mediate stress gradient effects on plant biomass allocation, with implications for coastal ecosystem services. J. Ecol. 2020, 108, 1227–1240. [Google Scholar] [CrossRef]
- Huang, H.; Cao, Y.; Xin, K.; Liang, R.; Chen, Y.; Qi, J. Morphological and physiological changes in Artemisia selengensis under drought and after rehydration recovery. Front. Plant Sci. 2022, 13, 851942. [Google Scholar] [CrossRef]
- Yang, J.; Zheng, W.; Tian, Y.; Wu, Y.; Zhou, D. Effects of various mixed salt-alkaline stresses on growth, photosynthesis, and photosynthetic pigment concentrations of Medicago ruthenica seedlings. Photosynthetica 2011, 49, 275–284. [Google Scholar] [CrossRef]
- Askari, E.; Ehsanzadeh, P. Effectiveness of exogenous salicylic acid on root and shoot growth attributes, productivity, and water use efficiency of water-deprived fennel genotypes. Hortic. Environ. Biotechnol. 2015, 56, 687–696. [Google Scholar] [CrossRef]
- Pan, Y.; Xie, Y.; Deng, Z.; Tang, Y.; Pan, D. High water level impedes the adaptation of Polygonum hydropiper to deep burial: Responses of biomass allocation and root morphology. Sci. Rep. 2014, 4, 5612. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, M.; Ding, Z.; Liu, B.; Jiang, M.; Lü, X. Light-acquisition traits link aboveground biomass and environment in inner saline-alkaline herbaceous marshes. Sci. Total Environ. 2023, 857, 159660. [Google Scholar]
- Minden, V.; Kleyer, M. Ecosystem multifunctionality of coastal marshes is determined by key plant traits. J. Veg. Sci. 2015, 26, 651–662. [Google Scholar] [CrossRef]
- Freschet, G.T.; Roumet, C.; Comas, L.H.; Weemstra, M.; Bengough, A.G.; Rewald, B. Root traits as drivers of plant and ecosystem functioning: Current understanding, pitfalls and future research needs. New Phytol. 2021, 232, 1123–1158. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhu, J.; Liang, J.; Li, M.; Huang, S.; Li, H. Urban dominant trees followed the optimal partitioning theory and increased root biomass allocation and nutrient uptake under elevated nitrogen deposition. Forests 2024, 15, 100. [Google Scholar] [CrossRef]
- Cardona-Olarte, P.P. Responses of Neotropical Mangrove Seedlings to Environmental Regulators. Doctoral Dissertation, University of Louisiana at Lafayette, Lafayette, LA, USA, 2004. [Google Scholar]
- Pang, B.; Xie, T.; Ning, Z.; Cui, B.; Zhang, H.; Wang, X. Invasion patterns of Spartina alterniflora: Response of clones and seedlings to flooding and salinity—A case study in the Yellow River Delta, China. Sci. Total Environ. 2023, 877, 162803. [Google Scholar] [CrossRef]
- Stuefer, J.F.; Hutchings, M.J. Environmental heterogeneity and clonal growth—A study of the capacity for reciprocal translocation in Glechoma-hederacea L. Oecologia 1994, 100, 302–308. [Google Scholar] [CrossRef]
- Benot, M.L.; Bonis, A.; Mony, C. Do spatial patterns of clonal fragments and architectural responses to defoliation depend on the structural blue-print? An experimental test with two rhizomatous Cyperaceae. Evol. Ecol. 2010, 24, 1475–1487. [Google Scholar] [CrossRef]
- Herben, T.; Nováková, Z.; Klimesová, J. Clonal growth and plant species abundance. Ann. Bot. 2014, 114, 377–388. [Google Scholar] [CrossRef]
- Minden, V.; Kleyer, M. Testing the effect-response framework: Key response and effect traits determining above-ground biomass of salt marshes. J. Veg. Sci. 2011, 22, 387–401. [Google Scholar] [CrossRef]
- Shen, X.; Jiang, M.; Lu, X.; Liu, X.; Liu, B.; Zhang, J. Aboveground biomass and its spatial distribution pattern of herbaceous marsh vegetation in China. Sci. China-Earth Sci. 2021, 64, 1115–1125. [Google Scholar] [CrossRef]
- Tang, H.; Liu, Y.; Lou, Y.; Yu, D.; Zhou, M.; Lu, X. Nitrogen availability affects the responses of marsh grass and sedge plants (Phragmites australis and Bolboschoenus planiculmis) to flooding time. Sci. Total Environ. 2024, 908, 168008. [Google Scholar] [CrossRef]
- Wang, Z.; Song, K.; Zhang, B.; Liu, D.; Ren, C.; Luo, L. Shrinkage and fragmentation of grasslands in the West Songnen Plain, China. Agric. Ecosyst. Environ. 2009, 129, 315–324. [Google Scholar] [CrossRef]
- Li, X.; Yang, F.; Liu, X. Evaluation of rice-reed-fish model in salinized wetland of west Songnen Plain, China. Chin. J. Eco-Agric. 2007, 15, 174–177. [Google Scholar]
- Hao, M. The Ecological Restoration Research on Momoge Scirpus planiculmis Wetland. Master’s Dissertation, University of Chinese Academy of Sciences, Changchun, China, 2016. [Google Scholar]
- Zhang, L.; Zhang, G.; Li, H.; Sun, G. Eco-Physiological Responses of Scirpus planiculmis to Different Water-Salt Conditions in Momoge Wetland. Pol. J. Environ. Stud. 2014, 23, 1813–1820. [Google Scholar]
- Lefcheck, J.S. Piecewisesem: Piecewise structural equation modelling in R for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Lou, Y.; Tang, Z. The Response of the Functional Traits of Phragmites australis and Bolboschoenus planiculmis to Water and Saline–Alkaline Stresses. Plants 2025, 14, 2112. https://doi.org/10.3390/plants14142112
Yang L, Lou Y, Tang Z. The Response of the Functional Traits of Phragmites australis and Bolboschoenus planiculmis to Water and Saline–Alkaline Stresses. Plants. 2025; 14(14):2112. https://doi.org/10.3390/plants14142112
Chicago/Turabian StyleYang, Lili, Yanjing Lou, and Zhanhui Tang. 2025. "The Response of the Functional Traits of Phragmites australis and Bolboschoenus planiculmis to Water and Saline–Alkaline Stresses" Plants 14, no. 14: 2112. https://doi.org/10.3390/plants14142112
APA StyleYang, L., Lou, Y., & Tang, Z. (2025). The Response of the Functional Traits of Phragmites australis and Bolboschoenus planiculmis to Water and Saline–Alkaline Stresses. Plants, 14(14), 2112. https://doi.org/10.3390/plants14142112