Abstract
Isatis tinctoria L. (Brassicaceae), also known as woad or dyer’s woad, is an ancient plant with a rosy future ahead. Most of the knowledge about woad is related to indigo dye production and its medicinal applications, especially its leaves. The general interest in woad has decreased with the rise of petroleum-based products. However, nowadays this plant is attracting interest again with industries reintroducing natural dyes. To meet the market demand in a sustainable manner, recent studies have focused specifically on woad seeds, leading to a valorization of the whole woad plant. This review provides an overview of the botanical, phytochemical composition, and properties of woad seeds, primarily supporting their cosmetic and pharmaceutical potential. From a chemical point of view, woad seeds mainly contain fatty acids, amino acids, phytosterols and glucosinolates. These compounds have been investigated through their extraction and analytical methods, as well as their properties and industrial applications.
1. Introduction
Isatis tinctoria L. (Brassicaceae), also known as woad, is native to Southeastern Russia and is located throughout Europe, North Africa, Asia and North America [1,2,3]. The plant’s cultivation and use have undergone many changes over the centuries. Woad was cultivated in Europe for many centuries during the Middle Ages [4,5,6,7]. Isatis tinctoria L. was originally a source of blue dye [3,4,5,6,8,9,10,11,12,13], and to a lesser extent, a paint component and an active ingredient in medicines [9,12,13] from the 12th to the 17th century. Its fermented rosette leaves were a source of indigo and used for luxury dyeing [3,11,12,13]. Over time, the woad industry has encountered two crises and has been replaced by other indigo sources, which offer higher recovery yields, brighter colors, and lower costs [12,13,14]. Woad cultivation first competed qualitatively and quantitatively with imported East India indigo during the seventeenth century [4,5,12,14,15]. Quantitatively, tropical Indigofera tinctoria had a five times better yield extraction (0.2% against 1%), which increased again with synthetic methods, especially the Baeyer–Drewsen [2,16] (1882), the Steingruber [17] (2004) and then the Steingruber [17] (2004) methods [13]. At the end of the 19th century, plant-derived indigo was neither competitive nor well understood compared to the synthetic one [4,14,15,18]. Therefore, the cultivation of Isatis tinctoria L. for indigo production declined, giving way to synthetic indigo involving a more profitable process and a chemically indistinguishable pigment [4,9,12,14,18]. Thus, the cultivation and uses of woad have been forgotten and are only growing in wild places such as roadsides or random lands [3,11,13,19]. Moreover, woad became problematic because of its invasive characteristics, especially in the USA [19,20]. After a century, with a view to returning to naturalness, woad crops and other indigo-producing crops have been recently reintroduced by growers [4,9,13] for natural dye production [4,9,10,15,18]. A zero-waste approach has been recently adopted for an overall recovery of Isatis tinctoria L., including branches, flowers, leaves, roots, and to a lesser but not insignificant extent, seeds and oil [13,21,22]. Some research laboratories, notably in the Hauts-de-France region, are involved in this global study of woad. The aim of such research is to guarantee the long-term re-establishment of this plant in the region, through high value-added applications. The present review will begin with the phenology and botanical characteristics of the woad to better understand the life cycle of the plant and the specifications of its fruits. In the second part, the review will compare the compounds of Isatis tinctoria to those of a similar species, Isatis indigotica Fort. Indeed, it allows us to see the differences and the similarities between the two Isatis species, and therefore put forward hypothetical woad molecules. Then, the cosmetic and pharmaceutical properties of each molecule listed in the literature for woad seeds are studied, with a view to promoting their use in these fields. The extraction and analysis processes are also described, with particular attention paid to energy costs and extraction process efficiency. Next, concentrations of compounds in Isatis tinctoria seeds will be highlighted and compared with those in Isatis indigotica, for each molecular family. Finally, focus will be placed on the antioxidant activity to enhance the wellness potential of woad fruits and seeds.
2. Phenology and Botanical Description
2.1. International Names and Botanical Situation
Isatis tinctoria L. is the scientific botanical name of the plant, but it can be called by different names throughout the world, even in the same country. These vernacular names are listed in Table 1.

Table 1.
International names of Isatis tinctoria L.
To complete this overview, it is important to remember the botanical situation of the plant studied. Indeed, the species Isatis tinctoria L. belongs to the Isatis genus and to the Brassicaceae (Cruciferous) family [23]. In fact, plants with similar botanical situations, especially those of the same genus, may have similar chemical contents and/or similar activities and properties.
2.2. Botanical Description of I. tinctoria L. Fruit
Isatis tinctoria L. is a biennial plant that produces fruit every two years. The whole fruit is composed of a fruiting pedicel, a pod, a pericarp, and a seed (Figure 1).

Figure 1.
Botanical drawing of Isatis tinctoria L. [26].
The fruiting pedicel is 5–10 mm long [3,11]. This ramification between the stem and the fruit is slender and subclavate at apex [3].
The silicle is an indehiscent and winged pear-shaped fruit [12,23,27]. The protective silicle is composed of a woody pod which encases the pericarp and the seed [3,12,19]. The pendulous seed pod turns from light green to blackish-brown when mature [11,22,28]. The hairless or shortly hairy fruit is 10–20 mm long, 3–6 mm wide, with apical wings 3.5–5 mm wide [3,11,22,27,28]. The silicle shape is qualified as oblong, obovate, oblanceolate and elliptic with a cuneate base [3,22,27]. It has a distinct midvein and inconspicuous lateral veins [3,11]. The yellowish-brown seed is 2.3–3.5 mm long and 0.8–1 mm wide [3,11,12]. It is an uniovulated ovary, narrowly oblong, and encircled by a pericarp [23].
2.3. Phenology and Germination
The phenology and the germination of Isatis tinctoria L. are described in Table 2, in order to gain a better understanding of its life cycle.
Table 2.
Phenology and germination of Isatis tinctoria.

Figure 2.
Isatis tinctoria growth stages.
Figure 2.
Isatis tinctoria growth stages.

2.4. Reproduction
Fuller [12] described that different modes of reproduction are possible for woad. On the one hand, the rosette can reproduce asexually from the tap root. On the other hand, the stalk (or bolted) form can reproduce asexually, from the roots, or sexually, leading to seeds. Woad seed reproduction does not exactly follow the Brassicaceae pattern. Indeed, seeds germinate quickly and plenty of seedlings develop, as in typical Brassicaceae. However, the fruits are indehiscent and naturally encapsulated by a germination inhibitor. This inhibitor blocks the germination of competing species, notably varieties of grasses and forbs such as other Brassicaceae.
Woad is known for its rapid growth and prolific seed production [20]. Such easy proliferation of this invasive species can lead to problematic management [19,20]. According to Farah et al. [20], the main factor in the spread of woad seed does not appear to be wind, but is strongly linked to the fruit pedicel, which behaves like a hook and adheres to objects, suggesting animal dispersal.
3. Comparison of the Overall Chemical Composition of Isatis tinctoria and Isatis indigotica Seeds
Four main molecular families have been investigated in woad seeds throughout the literature. All molecules identified in the seeds are listed in Table 3 and classified according to their chemical classes.
Table 3.
Fatty acid, amino acid, phytosterol and glucosinolate composition of Isatis tinctoria and Isatis indigotica seeds.
Each molecular family will then be described in detail. Only organic compounds will be studied in this review, although the microelement composition of woad seeds has been described by Kizil et al. [29].
Moreover, Isatis tinctoria and Isatis indigotica were long considered to be the same species, but there is now tangible evidence to the contrary [36]. Thus, a quick study of the phytochemical description of Isatis indigotica seeds will be presented in this review in order to link to other hypothetical molecules found in Isatis tinctoria seeds. Indeed, similar plants like these two may have similar phytochemical composition and properties. Nevertheless, authors of this review will focus only on molecules identified and proven in Isatis tinctoria seeds, and not on theoretical ones.
4. Fatty Acids
Isatis tinctoria contains 10 fatty acids (Figure 3) described in the literature. Of these, 4 are saturated fatty acids (SFAs), four are monounsaturated fatty acids (MUFAs) and two are both polyunsaturated fatty acids (PUFAs) and essential fatty acids for humans. Isatis indigotica also contains these fatty acids, but not only.
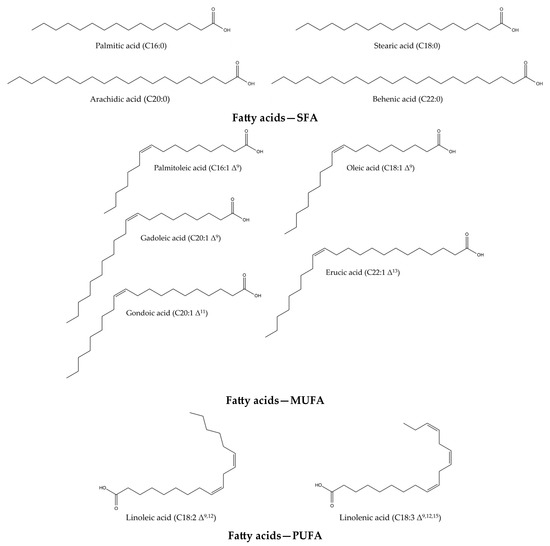
Figure 3.
Structures of the fatty acid compounds in Isatis tinctoria seeds.
4.1. Functions and Properties of Fatty Acids
PUFAs can be synthetized by humans, but not essential fatty acids like linoleic acid and linolenic acid. As mentioned Iba [37], essential fatty acids must be incorporated through a diversified diet, to reach the amounts essential for each individual organism. Gillingham et al. [38] have highlighted the health protective feature of MUFAs. Indeed, a diet enriched with MUFAs prevents or improves metabolic syndrome and the risk of cardiovascular disease, by favorably modulating blood lipids, blood pressure and insulin sensitivity. Thus, the quality and the quantity of dietary fats affect the risk of cardiovascular diseases [37,39]. MUFAs have slightly lower or equivalent effects to those of PUFAs in reducing low-density lipoprotein cholesterol and total cholesterol in plasma [38]. Nevertheless, MUFAs improve high-density lipoprotein cholesterol levels and may have hypotensive effects. Furthermore, preferential oxidation and metabolism of MUFAs have an impact on body composition and may reduce the risk of obesity [38]. In addition, when dietary saturated fatty acids are replaced with carbohydrates, MUFAs are efficient at maintaining high-density lipoprotein cholesterol levels, lowering triglyceride levels and enhancing insulin sensitivity, which could help people with metabolic syndrome or diabetes mellitus [38]. Furthermore, a deficiency in PUFAs can lead to a reduction in prostanoid production and a deterioration in the epidermal barrier. On the other hand, too high a level of PUFAs can also be problematic, inducing high cholesterol levels, high blood pressure, high platelet aggregation capacity, and hence cardiovascular diseases.
Some of the fatty acids listed below have interesting properties, notably in cosmetics. For instance, Spataro and Negri [40] have demonstrated emollient and moisturizing properties for use in soaps and body creams. Indeed, cutaneous applications of PUFAs have been shown to protect the skin barrier and fix skin lesions, in the manner of an emollient agent. The properties of SFAs, MUFAs, and PUFAs described in Table 4 focus mainly on cosmetic properties and, to a lesser extent, on pharmaceutical and food properties.
Table 4.
Industrial properties and applications of fatty acids contained in Isatis tinctoria seeds.
4.2. Oil Extraction and Analyses of Its Fatty Acid Contents
Studies providing fatty acid content mentioned different procedures for oil extraction, fatty acid preparation and analysis. These methods and, to a lesser extent, complementary ones will therefore be discussed.
For rigorous fatty acids analysis, some precautions must be taken for sample preparations. Firstly, the whole fruit must be dried to 0% moisture content. Then, the seeds are extracted from their silicle and pericarp, either manually or mechanically. Next, seeds are grinded using a blade mixer, to obtain different particle sizes [9,29,50].
Although several oil extraction techniques are described in the literature for fatty acids extraction, only Soxhlet [9,29,31,32,51] and Ultrasound-Assisted Extraction (UAE) [31] were chosen by the authors who provided the fatty acid contents reported in the present study.
The conventional Soxhlet method is an accessible technique but has significant disadvantages. Soxhlet extraction continuously recycles the extractive solvent but is also time-consuming (2 h–8 h). As a result, this method requires high to moderate energy consumption due to the constant and prolonged, but not excessive, heating of the solvent. Also, Soxhlet extraction is applicable at high temperatures and boosts process kinetics, but the technique has a low extraction efficiency [31,52].
The UAE technique uses high-frequency sound waves to promote cavitation and fatty acid extraction. This technique requires a moderate amount of energy, particularly for generating ultrasound over a prolonged period.
Many other extraction techniques could have been described, such as cold-pressed extraction [53,54,55], acid hydrolysis extraction (method 95.402 of the Association of Official Analytical Chemists international (Official methods of analysis, 16th edn. AOAC, Arlington), Accelerated Solvent Extraction (ASE) [52], or maceration coupled with UAE [50,52]. We note that Romdhane and Gourdon [50] used maceration coupled with ultrasound to extract fatty acids from woad seeds, but no concentrations of these fatty acids were mentioned in their publication.
Consequently, we can rank all these extraction methods in ascending order of energy costs: (1) cold-pressed extraction; (2) UAE; (3) maceration combined with ultrasounds; (4) solid-liquid extraction; (5) Soxhlet; (6) ASE. Indeed, each technique has specific advantages, but thermal and pressure-based methods (ASE, Soxhlet) tend to have higher energy costs than room-temperature methods such as maceration and solid-liquid extraction.
Once the oil has been extracted from the biomass, two sample preparation steps must be followed. The first one consists of evaporating the solvent from the extract. The objective of this step is to concentrate oil extracts by totally removing the solvent using a rotary evaporator or a nitrogen stream [9,29], then analyzing them by Gas Chromatography (GC) [9,50].
The second step, as performed by Romdhane and Gourdon [50], consists of converting fatty acids into fatty acid methyl esters (FAMEs) for gas chromatography analysis. The authors mentioning the fatty acid contents of woad seeds used TBME method [9] and transmethylation [29,32], but there are several methods for converting fatty acids into their corresponding methyl esters, such as the BF3 method, the BHT method [29], and many other transesterification [56,57] and esterification methods [58]. The choice of method depends mainly on the composition of the oil. In fact, some methods are generally more efficient, others are recommended for samples with low acidity, and still others for samples with high acidity. To conclude, there are various methods for converting fatty acids into volatile methyl esters, including some that are not mentioned in this review. The choice must be made by considering simultaneously the composition of the oil, the fatty acids of interest, the necessary reagents, and the availability of equipment.
To calculate the average oil content of seeds, samples can be characterized gravimetrically and by GC with different types of detectors, such as Flame Ionization Detector (FID), Mass Spectrometry (MS), and Thermal Conductivity Detector (TCD). The authors who provided the fatty acid contents utilized GC-FID [9,32], GC-MS [29,31] and GC-TCD [51] methods, enabling accurate quantitative determination. To choose the most suitable method, it is necessary to consider the availability of equipment, standards and/or mass spectrometry libraries. Moreover, GC-MS and GC-FID methods are the most widely described techniques in the literature for fatty acid analysis.
Besides the composition of the oil itself, oil can be characterized in many ways, as mentioned by Gambert et al. [59] For instance, color and clarity can be determined visually, the refraction index is identified with a refractometer, and density is calculated with a pycnometer. Viscosity is studied with a rotary viscometer, calculating the torsional force, the speed of the axis, and its characteristics. The moisture content is calculated after a period of heating using a thermo-balance. Numerous indexes are registered to characterize oils. (1) The acid index (NF EN 14104 for fatty acid methyl esters) refers to the amount of free fatty acids. The free acid content of fats increases over time, making it a good indicator of their state of deterioration. (2) The total acid number determines the sum of all acid compounds (ASTM D664-18e2 for petroleum products). (3) The saponification index (NF EN ISO 3657 for fats; ASTM D94-07(2017) for petroleum products) gives the molecular weight of all medium-chain fatty acids present in a sample. (4) The iodine index (NF EN ISO 3961 for fats; NF EN 14111 for fatty acid methyl esters; ASTM D5554-15(2021) for fats and oils) is used to assess the level of unsaturation in the oil. (5) The peroxide index (NF EN ISO 3960 for fats; ASTM E299-17a for organic solvents) is utilized to calculate the oxygen level in the oil.
4.3. Fatty Acid Composition in Isatis tinctoria Seed Oil
Table 5 compares the ratio of fatty acids from Isatis tinctoria and Isatis indigotica seeds according to crop and production site.
Table 5.
Fatty acid composition of Isatis tinctoria and Isatis indigotica seeds at harvest.
It appears that the contents of all SFAs, MUFAs, and PUFAs varies according to these different sites. The Turkish site quantified higher amounts of palmitic acid (SFA) than the French and US sites for Isatis tinctoria. Higher quantities of C20:1 fatty acid (MUFA) were found at the US site than at the other sites for Isatis tinctoria. The Turkish site had low quantities of erucic acid (MUFA) compared to the other two countries for Isatis tinctoria. Linoleic and linolenic acids (PUFAs), which are essential fatty acids, were found in low quantities at the Turkish site compared to those in France and the USA for Isatis tinctoria.
Roche et al. [9] and Mikolajczak et al. [51] showed that unsaturated fatty acids (MUFAs and PUFAs) are predominant in the composition of Isatis tinctoria seed oil. This trend is also confirmed by Li et al. [31] and Angelini et al. [32] for Isatis indigotica seed oil. Also, Kizil et al. [29] pointed out that MUFAs are the main class of fatty acid in woad oil. By cross-referencing data from different production sites, the main compounds in Isatis tinctoria and Isatis indigotica oils are oleic acid, erucic acid, and linolenic acid. The predominance of these three fatty acids is a common characteristic of the Brassicaceae oils [37]. These fatty acids can be useful in the cosmetics industry due to their emollient, surfactant, and fragrance properties. However, a cautionary note must be issued as the erucic acid content of the seeds greatly exceeds the amount permitted for food use. This amount must therefore also be controlled for cosmetic or pharmaceutical applications. Varietal selection limiting the erucic acid content within the plant could be a sustainable solution to the toxicity issue of this molecule, while complying with the regulations governing its potential applications.
The physico-chemical characteristics of the oil, based on the results of Dolya et al. [30], are summarized in Table 6.
Table 6.
Physico-chemical characteristics of Isatis tinctoria and Isatis indigotica oils.
According to Iba’s thesis [37], the fatty acid composition, particularly with a high concentration of unsaturated fatty acids, provides specific physical properties. Indeed, low viscosity is explained by the high amount of unsaturated fatty acids. The high density implies an oil with quite high saponification and iodine indexes. The refractive index is induced by the presence of unsaturated fatty acids with long carbons chains. However, as mentioned above, the fatty composition depends on the production site, the harvest period and the harvest year. Thus, the values presented in Table 6 are not necessarily constant for Isatis seed-extracted oil.
The refractive index is similar for both Isatis tinctoria oil and Isatis indigotica oil. Isatis tinctoria oil has a lower acid index, i.e., a lower free fatty acid content, than Isatis indigotica oil. The saponification index provides information on the average molecular weight of all bound and free fatty acids present in a sample. Isatis tinctoria oil has a higher saponification index, i.e., a lower molecular weight of all medium-chain fatty acids, than Isatis indigotica oil. Isatis tinctoria oil has a higher iodine index, i.e., more unsaturations or double bounds, than Isatis indigotica oil.
5. Amino Acids
Isatis tinctoria contains 18 amino acids described in the literature. Isatis indigotica also contains these amino acids, except for hydroxyproline. Only hydroxyproline is shown in Figure 4, as the other amino acids described are relatively common.
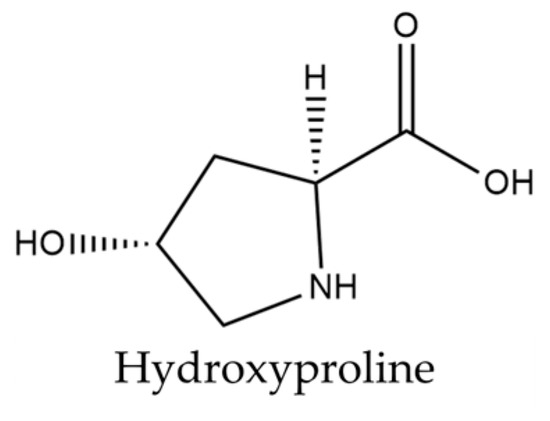
Figure 4.
Structure of hydroxyproline present in woad seeds.
5.1. Functions and Properties of Amino Acids
In general, amino acids play a crucial role in the structure, metabolism and physiology of cells, as they are the building blocks of peptides and proteins. In a study, Wu et al. [60] described that amino acids have significant biological, nutritional and health functions. In addition, amino acids are cell signaling molecules; they are key precursors for proteins, including hormones and low-molecular-weight nitrogenous compounds. Amino acids are also regulators of metabolic pathways and processes, which are essential for health, such as growth, development, reproduction, immunity, and homeostasis of living organisms. For instance, they contribute to the regulation of gene expression and protein phosphorylation cascades. Nevertheless, physiological concentrations of amino acids and their products (e.g., nitric oxide, polyamines, glutathione, taurine, thyroid hormones, and serotonin) are necessary for proper metabolic function. Too high levels of these substances can lead to neurological disorders, oxidative stress, and cardiovascular disease. An optimal balance of amino acids in dietary supplementation is essential for good health. Amino acids are valuable for improving health at different stages of life, for example in cases of fetal growth retardation, neonatal morbidity and mortality, intestinal dysfunction and weaning-related wasting syndrome, obesity, diabetes, cardiovascular disease, metabolic syndrome, infertility, and infection. They are also beneficial for enhancing the efficiency of metabolomic modifications to improve muscle protein synthesis, muscle growth, milk production, egg and meat quality, and athletic performance, while preventing excess fat deposition and reducing adiposity in living beings.
A total of 17 of the 20 basic amino acids are referenced for Isatis tinctoria. Hydroxyproline, formed from proline by hydroxylation, is the only derived molecule described in the literature for this plant. Although hydroxyproline is not specific to Isatis and is not the most abundant amino acid, this molecule remains interesting. Kumar Srivastava et al. [61] explained that its two isomeric forms (trans-4-hydroxy-L-proline and trans-3-hydroxy-L-proline) play a key role in the synthesis and thermodynamic stability of collagen fiber, suggesting a strong potential for hydroxyproline in cosmetic application. These two molecules are found in animal collagens, which are qualified as structural proteins.
VanEtten et al. [62] highlighted that hydroxyproline is found in the seed pod and pericarp of the woad fruit. It can therefore be assumed that structural proteins are present in these two parts of the plant. The protective property of the woad pod may be, to some extent, due to the presence of these proteins. The amino acids present in woad seeds have interesting cosmetic and pharmaceutical applications, which are summarized in Table 7.
Table 7.
Industrial properties and applications of the amino acids contained in Isatis tinctoria seeds.
5.2. Extraction and Analyses of Amino Acids
The research conducted by Miller et al. [33] focused on all amino acids in the seed pod of the woad, while VanEtten et al. [62] concentrated on hydroxyproline content in both the pod and the pericarp. With regard to sample preparation, seeds were grounded and then extracted with petroleum ether to remove the oil [33,62]. Next, the samples are acid-hydrolyzed before analysis [33,62,64].
Then, amino acids are isolated from the acid hydrolyzate of the seed pod using a procedure similar to that described by Levine [65]. The two imino acids of L-proline and L-hydroxyproline undergo acid hydrolysis by heating with nitrous acid (Figure 5).
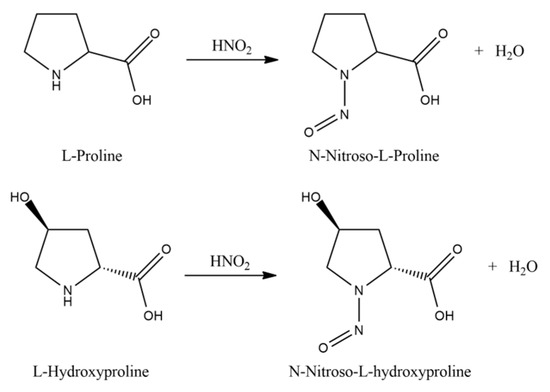
Figure 5.
Reaction of the acid hydrolysis of L-proline and L-hydroxyproline.
Low-Pressure Liquid Chromatography (LPLC) separates the two nitrosoamines thus obtained. Once crystallized, L-hydroxyproline is analyzed by X-ray fluorescence spectrometer (XRF) and its optical rotation is calculated. Furthermore, the sample elementary analysis gives C, 45.7%; H, 6.96%; N: 10.5%. The theoretical one gives C, 45.7%; H, 6.87%; N, 10.68%.
VanEtten et al. [62] highlighted the following two methods for quantitative amino acid analysis. The first one is the Ion Exchange Chromatography (IEC) method developed by Spackman, Stein and Moore [66] using an MS instrument and an UV detector at 440 nm. The second one is a Thin-Layer Chromatography (TLC) method, which refers to the method developed by Becker, Milner and Sagel [67].
5.3. Amino Acid Composition
Miller et al. [33] determined the overall amount of amino acids found in woad seeds harvested in the USA (Table 8).
Table 8.
Amino acid composition of Isatis tinctoria and Isatis indigotica seeds at harvest.
On a dry basis and after acid hydrolysis, I. tinctoria contains 12.5% c of crude protein in the whole seed and 14.3% c in the extracted meal. This species contains 12.6% of oil, also on a dry basis. In seed meals, 69.8% of the total nitrogen was accounted for as amino acids. Moreover, VanEtten et al. [62] calculated 150 mg of hydroxyproline per gram of nitrogen in Isatis tinctoria, confirmed by Miller et al. [33] who found 148 mg of this molecule per gram of nitrogen. The most abundant amnio acids in woad seeds are glutamic acid and, to a lesser extent, aspartic acid.
6. Phytosterols
Isatis tinctoria contains seven phytosterols described in the literature, including two found in trace amounts (Figure 6). Phytosterols are currently not described in Isatis indigotica.
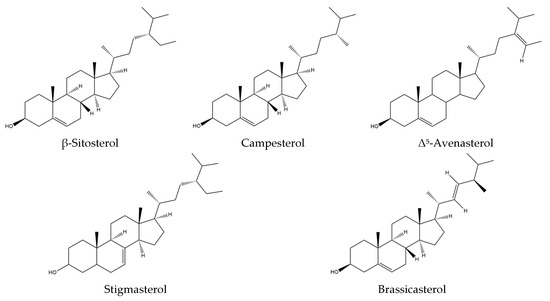
Figure 6.
Structures of the phytosterol compounds in Isatis tinctoria seeds.
6.1. Functions and Properties of Phytosterols
Phytosterols have applications in everyday life. Indeed, they add value to crop yields as well as human nutrition, pharmaceutical, and cosmetic applications [9]. Consuming phytosterols significantly decreases serum cholesterol levels [68,69,70,71,72,73,74,75], which lowers the risk of cardiovascular diseases [69,72,73,74,75]. Thanks to phytosterol-enriched diets, the toxic effects of these compounds enable tumoral cell apoptosis [68,72,74,75]. This molecular property helps to reduce the risk of common cancers such as lung, stomach, colon, breast, and prostate cancers [68,69,72,74,75]. In addition, Burg et al. [76] have shown that mixtures of phytosterols, mainly composed of stigmasterols, may be useful in preventing Alzheimer’s disease [68]. Other therapeutic properties have been reported, such as anti-inflammatory, anti-bacterial, anti-ulcerative, angiogenic, immunomodulatory, antidiabetic, anti-tumoral, antinociceptive, antituberculosis, and anti-atherosclerotic ones [68,69,73,74]. To summarize, Plaza et al. [77] emphasized that food and dietary supplements enriched with phytosterols offer many health benefits. In a less well-documented way, phytosterols also have cosmetic properties [41,68]. Table 9 summarizes all the properties mentioned above, focusing mainly on the cosmetic and pharmaceutical ones. However, some compounds have no described applications in cosmetics, probably due to a lack of tested biological activities.
Table 9.
Industrial properties and applications of phytosterols contained in Isatis tinctoria seeds.
6.2. Extraction and Analyses of Phytosterols
Phytosterols are contained in woad-extracted oil but cannot be separated from it as they are. Indeed, the samples need to undergo a saponification step (for phytosterol esters) and a derivatization (silylation) step. Finally, sterol trimethylsilyl ether derivatives can be analyzed by GC [9,83].
6.3. Phytosterol Composition
The authors want to point out that the phytosterols in woad seeds are not very well described in the literature, explaining why few references are available, and even fewer recent ones.
With regard to phytosterol composition, Roche et al. [9] studied phytosterol compounds according to their quantity encountered in woad seeds and their site of collection (Table 10).
Table 10.
Phytosterol composition of Isatis tinctoria seeds at harvest, collected by the CAPA Institute (Coopérative Agricole des Plaines d’Ariège), France (data from [9]).
The main phytosterol found in woad seeds is β-sitosterol, which accounts for more than 50% of the total sterol content. The second main phytosterol is campesterol, representing 20% of the sterol content. The third most abundant one is Δ5-avenasterol, accounting for approximately 15% of the sterol content.
7. Glucosinolates
Isatis tinctoria contains 10 glucosinolates described in the literature (Figure 7). Isatis indigotica contains the same glucosinolates but in different amounts.
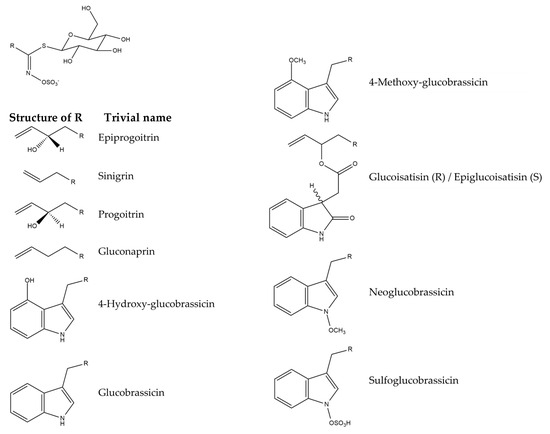
Figure 7.
Structures of the glucosinolate compounds in Isatis tinctoria seeds [35].
When plants are damaged, for example by chewing, they secrete an enzyme called myrosinase, which breaks down glucosinolates into bioactive compounds such as isothiocyanates, thiocyanates, and nitriles. These breakdown products are responsible for the antioxidant, anticarcinogenic, and antimicrobial properties of glucosinolates.
7.1. Functions and Properties of Glucosinolates
Glucosinolates play a role in plant defense mechanisms against herbivores, insects, and pathogens, particularly when these metabolites are hydrolyzed by myrosinase [84,85]. The health benefits of consuming plants containing glucosinolates from Brassicaceae are frequently emphasized [86,87,88]. These benefits are related to their antibacterial, antifungal, anticancer, antioxidant, and anti-inflammatory activities [86,87,88,89]. In addition, glucosinolate catabolites, such as isothiocyanates and nitriles, have therapeutic effects in carcinogenesis [89,90]. For instance, by providing chemoprotective effects against carcinogenesis in rat tissues, isothiocyanates offer potential therapeutic benefits for humans [89]. Furthermore, allyl isothiocyanates derived from the biocatalyzed hydrolysis of glucosinolates exhibit antimicrobial activities against specific bacteria [87,91]. From a cosmetic point of view, the antioxidant and anti-inflammatory properties of glucosinolates and their derivatives could help to improve tissue damage induced by oxidative stress and attenuate DNA damage caused by UVB rays, thereby limiting skin aging. Table 11 summarizes the pharmaceutical properties of glucosinolates, but no cosmetic properties are mentioned due to a lack of information.
Table 11.
Industrial properties and applications of glucosinolates contained in Isatis tinctoria seeds.
7.2. Sample Preparation, Extraction and Analyses of Glucosinolates
Few methods for extracting and analyzing glucosinolates are described in studies providing the glucosinolate content in woad seeds. These methods and, to a lesser extent, complementary ones will therefore be considered.
Mohn and Hamburger [35] and Angelini et al. [32], who studied the glucosinolate content in seeds of Isatis tinctoria and Isatis indigotica, respectively, did not provide any information regarding seed preparation and storage conditions. However, it has been noticed that freeze-drying of the raw material is recommended for long-term storage to maintain the recovery yield of glucosinolates [101].
Although several techniques for extraction glucosinolates are described in the literature, only the Soxhlet [32] and ASE [35] methods were chosen by the authors, who provided the glucosinolate contents mentioned in this study.
As mentioned above, Soxhlet extraction is a conventional method that recycles extractive solvent, but it is also time-consuming (2 h–8 h). This method therefore requires high to moderate energy consumption due to the steady and prolonged, but not excessive, heating of the solvent [102].
The unconventional ASE method is advantageous in terms of time, solvent consumption, energetic efficiency, and selectivity of glucosinolate extraction. However, this innovative process is quite expensive and/or not currently suitable for large-scale industrialization [101]. The ASE method uses high temperatures (35 to 200 °C) and high pressures to accelerate the extraction process from woad seeds and improve recovery yields [103]. This results in relatively high energy consumption due to the need to maintain heat and pressure. It is not advisable to extract glucosinolates above 50 °C, as they are thermosensitive and could therefore be degraded, as could their biological efficiency (notably their antioxidant and antimicrobial properties) [102]. ASE does not offer any significant advantage in terms of recovery yield compared to conventional maceration extraction and is not suitable for industrial-scale production. Glucosinolates recovered by ASE have lower biological activities compared to those obtained by other types of extraction, particularly with regard to their inhibition of bacterial growth [101].
It should be noted that many other extraction techniques could have been described, such as reflux set-up [104,105], UAE [101,106], or Supercritical Fluid Extraction (SFE) [107], in order to observe differences in efficiency. Also, glucosinolates must be efficiently separated from other water-soluble compounds in the sample, such as proteins or phenolic compounds.
On the other hand, in terms of energy costs, a ranking of all these extraction methods in ascending order can be suggested: (1) Soxhlet; (2) reflux set-up; (3) UAE; (4) ASE; (5) SFE. Thermal and pressure-based methods, such as ASE and SFE, have significantly higher energy costs than room-temperature methods such as maceration or those using ultrasound activation. Indeed, the choice of an extraction method depends on specific requirements in terms of extraction efficiency, the nature of the compounds to be extracted, and energy and economic costs.
The isolation of glucosinolates from aqueous plant extracts is a challenging process due to their hydrophilic nature, which Mohn and Hamburger [35] and Angelini et al. [32] did not put forward [101]. To overcome this problem, the remedies for purifying glucosinolates include ion-exchange resin-based methods and chromatographic techniques. The approach highlighted by Thies [108] and Wang et al. [106] emphasized ion-exchange resin-based methods, which aim to trap glucosinolates to purify them, thanks to their ionized nature. Ion-exchange resin-based methods, like batch adsorption, are simpler, faster, and more suited to industrial scales, but they do not easily allow the separation of the various glucosinolates in the extract.
On the other hand, chromatographic separation methods aim to isolate the integrality of glucosinolates from plant materials [101]. Preparative High-Performance Liquid Chromatography (prep-HPLC) and High-Speed Counter-Current Chromatography (HSCCC) can be used to isolate pure glucosinolates from Brassicaceae [109,110]. Chromatographic techniques are effective for isolating individual glucosinolates of high purity, but they require precise and expensive equipment, and also consume large amounts of solvents, energy, and time.
The choice of isolation method depends on the intended downstream applications for the isolated glucosinolates [101]. For instance, if the goal is to study the properties of an individual glucosinolate, a chromatographic technique may be preferred. However, if the aim is to obtain a large quantity of glucosinolates for general use, an ion-exchange resin-based method may be more practical. In summary, the isolation of glucosinolates requires a thorough study of their properties and the specific needs of the application. Technological and methodological advances continue to improve the efficiency and selectivity of glucosinolates isolation, making it a rapidly evolving field of research and development.
After being extracted from plant tissues and partially purified, glucosinolates can be identified and quantified, in particular by High-Performance Liquid Chromatography (HPLC). The authors, who quantified glucosinolates in the seeds of Isatis tinctoria and Isatis indigotica, used HPLC coupled with a Diode Array Detector (HPLC-DAD) [32] and HPLC-MS [35] techniques.
Many other analytical techniques could have been described for qualitative analysis, such as X-ray analysis [111,112], Fourier-transform infrared spectroscopy [113,114], Near-InfraRed Spectroscopy (NIRS), Nuclear Magnetic Resonance spectrometry (NMR) [115,116], ion-exchanger cartridge (desulfation process) [117,118,119], TLC, reversed-phase Ion-Pair Chromatography (IPC) for apolar glucosinolates, and normal-phase HydrophILic Interaction Chromatography (HILIC) for polar glucosinolates. To conclude, the efficiency of the entire extraction process depends on the accurate identification of glucosinolates. To meet the required criteria, a comprehensive characterization must include at least 1H NMR, mass spectrometry (MS), and infrared (IR) analyses. Further spectroscopic proofs, such as 13C NMR, MS-MS, and elemental analysis, clearly provide crucial structural information on the glucosinolate of interest.
Other methods less described in the literature can also be used to quantify glucosinolates and their breakdown products, as summarized by Śmiechowska et al. [120]. Indeed, some of these methods are based on measuring the glucose or sulfates released following the treatment of glucosinolates by myrosinase. Other methods, such as the glucose test, gravimetric methods, titrimetric methods, Enzyme-Linked ImmunoSorbent Assay (ELISA), GC (after derivatization), Supercritical Fluid Chromatography (SFC) (breakdown products), or Capillary Electrophoresis (CE) (both glucosinolate and their breakdown products) can also be used to quantify these molecules [120]. Furthermore, the amount of glucosinolates can be calculated by various HPLC dosages methods, such as (1) HPLC-UV methods for total glucosinolates [102,121] (at 229 nm and 420 nm) and desulfated glucosinolates [122,123,124] (at 230 and 229 nm); (2) LC-MS/MS methods for desulfated glucosinolates [125,126] and glucosinolates [127]; (3) HPLC-UV regression model for total glucosinolates (at 425 nm) [128].
Finally, HPLC-UV and LC-MS/MS methods are the most commonly used, despite the high costs associated with equipment acquisition and maintenance. Moreover, pure glucosinolate standards are required for accurate LC quantification, but these are not always readily available. Thus, these methods allow for accurate analysis but are not always the most practical ones. In addition, some of the methods mentioned above involve desulfation, which is controversial and requires time-consuming steps. Furthermore, spectrophotometric dosages, and especially the predictive formulas associated with them, offer both simplicity and cost savings.
7.3. Glucosinolate Composition
Glucosinolates are sulfur-containing glucose derivatives found in many plant families, especially in the Brassicaceae family, which includes Isatis tinctoria and Isatis indigotica. Table 12 classifies glucosinolate compounds according to their quantity and collection site.
Table 12.
Glucosinolates composition of Isatis tinctoria and Isatis indigotica seeds at harvest.
The main glucosinolates in Isatis tinctoria seeds are aliphatic ones, and especially epiprogoitrin [32,35]. Isatis indigotica also has more aliphatic glucosinolates than indolic ones. Its main compound is either progoitrin or epiprogoitrin, depending on the collection site. On the other hand, Isatis tinctoria contains more glucosinolates overall than Isatis indigotica.
8. Antioxidant Potential
The antioxidant potential of Isatis tinctoria has been studied with regards to its oilseed cakes. The raw materials underwent controlled loss-on-drying of less than 10% and were then ground. The hexane extracts were obtained directly from the raw material and all other extracts were defatted with hexane before polar extractions.
Peschel et al. [129] conducted several antioxidant tests on numerous oilseed cakes and plant antioxidants. These samples were compared in Table 13 to a synthetic antioxidant standard (butylated hydroxytoluene or BHT) and three commercial samples of well-established plant antioxidants (grape seed extract, rosemary superfluid extract and green tea extract).
Table 13.
Yield and activity of Isatis tinctoria oilseed cakes in comparison to BHT, grape seed extract, rosemary SF and green tea extract (data from [129]).
The different assays were undertaken as follows. The Total Phenolic Content (TPC) was studied with the Folin–Ciocalteu reagent and gallic acid as the standard phenolic molecule. The equivalent in µg gallic acid (GAE) of a sample is determined by spectrophotometry (no wavelength described). The free radical scavenging activity (DPPH assay) uses the 1,1-diphenyl-2-picryl-hydrazil (DPPH) reagent and a spectrophotometric determination at 517 nm. The activity is calculated as a percentage of inhibition compared to the blank control. The superoxide anion scavenging activity (NTZ assay) is determined with the nitroblue tetrazolium (NTZ) reagent. The percentage of inhibition is also calculated spectrophotometrically at 560 nm. The Rancimat assay evaluates the inhibition of the oxidation rate of stressed linoleic acid by colorimetric detection, after the oxidation and heating of the samples.
For Isatis tinctoria, the most effective extraction in terms of TPC was achieved with ethanol 75%, with 106.10 mg/GAE g.
The radical scavenging activity was investigated with the DPPH assay for free radical scavenging activity and the NTZ assay for superoxide anion scavenging activity. The decolorizing process during the DPPH assay was monitored, and woad extracts with propylene glycol appear to slightly reduce free radicals at a concentration of 10 µg/mL. According to the NTZ assay results, the activities of the polar woad extracts are significantly higher than the commercial rosemary extract (4.30%).
The Rancimat® assay is an accelerated aging test that highlights the oxidation stability of fats and oils. The focus was therefore on the inhibition of linolenic acid oxidation, revealing an inhibition of 30.07%. Also, this assay indicates that the woad extract with propylene glycol has better antioxidant activity than the rosemary standard.
Finally, woad seed extracts, and particularly the propylene glycol extracts, demonstrate interesting antioxidant activity. The toxicity of these extracts could be tested for high-value-added applications, such as in the cosmetics or pharmaceutical industries.
9. Conclusions
The review presented Isatis tinctoria, especially its fruits and seeds, from a phenological and botanical perspective. This biennial species produces fruits composed of three parts: the pod, the seed, and the pericarp. The present study mainly focused on seeds and only briefly examined these three fruit sections due to a lack of information. A complementary study based on the different parts of the fruit could be interesting to better understand the distribution of molecules.
Since Isatis tinctoria species and Isatis indigotica species had long been described as the same, comparing their phytochemical composition was relevant. Although the two species were ultimately dissociated, they exhibit obvious phytochemical similarities. Indeed, most of the fatty acids, amino acids, and glucosinolates described in Isatis tinctoria have also been described in Isatis indigotica. However, Isatis indigotica contains additional molecules described in these first two molecular families. Further study of the fruits and seeds of woad could determine whether these additional molecules are also present in Isatis tinctoria seeds, or whether they are specific to Isatis indigotica seeds. For the time being, these few molecules remain only hypothetical in terms of the composition of woad seeds. On the other hand, four molecular families are described in woad seeds: fatty acids, amino acids, phytosterols and glucosinolates. Also, as some phenolic compounds have been described in Isatis indigotica seeds and most plants contain these metabolites, the authors suggest that further studies on woad seeds could focus on this molecular family among others that have not yet been documented. This line of research could be an opportunity to broaden knowledge about this little-described fruit plant.
In addition, numerous cosmetic properties and, to a lesser extent, pharmaceutical and food properties have been summarized. Once their non-toxicity has been verified, woad seeds could be used in these fields of application. However, as the hulling of woad fruit is a very meticulous and time-consuming process, its use would be better suited to high value-added applications.
Furthermore, extraction and analysis methods have been studied for each molecular family. Although these processes are readily available for the study of these compounds, the authors wanted to focus on the energy consumption and efficiency of these techniques, in order to better align with green chemistry.
In addition, the compound content of woad seeds was examined by cross-referencing available sources. This study highlighted the main compounds of Isatis tinctoria and compared their concentrations with those of another Isatis species. Thanks to this comparison, it is possible to focus on one species or another, depending on the compound or family of compounds targeted.
Finally, the antioxidant activity of woad oilseed cakes has been demonstrated. Once the toxicity issues have been resolved, these antioxidant activities could potentially be exploited in the cosmetics and pharmaceutical industries. Also, a complementary study on the antioxidant activity of whole fruit, seed, oil and pericarp could be relevant. This supplemental study could also link antioxidant activity to single molecules or to a symbiotic group of molecules.
Author Contributions
Conceptualization, N.J. (Nicolas Joly), N.J. (Nathalie Jullian), É.C. and P.M.; investigation, J.D. and R.V.; resources, N.J. (Nicolas Joly), N.J. (Nathalie Jullian), V.L., R.V. and P.M.; writing—original draft preparation, J.D.; writing—review and editing, N.J. (Nicolas Joly), N.J. (Nathalie Jullian), É.C. and P.M.; visualization, J.D.; supervision, N.J. (Nicolas Joly) and P.M.; project administration, N.J. (Nicolas Joly) and P.M.; funding acquisition, N.J. (Nicolas Joly), N.J. (Nathalie Jullian) and P.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Hauts-de-France Region, Unité Transformations & Agroressource and Université d’Artois.
Acknowledgments
The authors would like to thank the Hauts-de-France Region for its financial support in the GRAWAI project realization, in addition to the industrial partners of the project: SAS Couleurs Végétales de France and La Savonnerie des Hauts-de-France. The GRAWAI project is supported by Bioeconomy For Change, EuraMaterials, Cosmetic Valley competitiveness clusters and SATT Nord.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Young, J.A.; Evans, R.A. Germination of Dyers Woad. Weed Sci. 1971, 19, 76–78. [Google Scholar] [CrossRef]
- Prasad, R. Indigo—The Crop that Created History and then Itself Became History. Indian. J. Hist. Sci. 2018, 53, 296–301. [Google Scholar] [CrossRef]
- eFlora. Available online: https://www.efloras.org/florataxon.aspx?flora_id=2&taxon_id=200009571 (accessed on 11 March 2024).
- Sales, E.; Kanhonou, R.; Baixauli, C.; Giner, A.; Cooke, D.; Gilbert, K.; Arrillaga, I.; Segura, J.; Ros, R. Sowing date, transplanting, plant density and nitrogen fertilization affect indigo production from Isatis species in a Mediterranean region of Spain. Ind. Crops Prod. 2006, 23, 29–39. [Google Scholar] [CrossRef]
- Kokubun, T.; Edmonds, J.; John, P. Indoxyl Derivatives In Woad In Relation To Medieval Indigo Production. Phytochemistry 1998, 49, 79–87. [Google Scholar] [CrossRef]
- Angelini, L.G.; Tozzi, S.; Nassi o Di Nasso, N. Differences in leaf yield and indigo precursors production in woad (Isatis tinctoria L.) and Chinese woad (Isatis indigotica Fort.) genotypes. Field Crops Res. 2007, 101, 285–295. [Google Scholar] [CrossRef]
- Giot, C.; Clausse, E.; Carbonnier, Y.; Vauquelin, R.; Joly, N.; Martin, P.; Schneider, P. Isatis tinctoria & Indigo; A Spatiotemporal Culture in France and Elsewhere-Historical Review. Am. J. Plant Sci. 2025, 16, 287–300. [Google Scholar] [CrossRef]
- Clark, R.J.H.; Cooksey, C.J.; Daniels, M.A.M.; Withnall, R. Indigo, woad, and Tyrian Purple: Important vat dyes from antiquity to the present. Endeavour 1993, 17, 191–199. [Google Scholar] [CrossRef]
- Roche, J.; Mouloungui, Z.; Cerny, M.; Merah, O. Fatty acid and phytosterol accumulation during seed development in three oilseed species. Int. J. Food Sci. Technol. 2016, 51, 1820–1826. [Google Scholar] [CrossRef]
- Spataro, G.; Taviani, P.; Negri, V. Genetic Variation and Population Structure in a Eurasian Collection of Isatis tinctoria L. Genet. Resour. Crop Evol. 2007, 54, 573–584. [Google Scholar] [CrossRef]
- eFlora. Available online: https://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=200009571 (accessed on 11 March 2024).
- Fuller, A.T. An Autecological Study of Dyers Woad (Isatis tinctoria L.) on Utah Rangeland. Master’s Thesis, Utah State University, Logan, UT, USA, 1985; pp. 1–61. [Google Scholar] [CrossRef]
- Vauquelin, R.; Juillard-Condat, L.; Joly, N.; Jullian, N.; Choque, E.; Martin, P. Study of Woad (Isatis tinctoria L.)-Extracted Indoxyl Precursors Conversion into Dyes: Influence of the Oxidative Media on Indigo Recovery Yields and Indigotin/Indirubin Ratio Measured by HPLC-DAD Method. Molecules 2024, 29, 4804. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, E.F. The Woad Plant and its Dye. Nature 1931, 127, 658–659. [Google Scholar] [CrossRef]
- Gilbert, K.G.; Cooke, D.T. Dyes from plants: Past usage, present understanding and potential. Plant Growth Regul. 2001, 34, 57–69. [Google Scholar] [CrossRef]
- Baeyer, A.; Drewsen, V. Darstellung von Indigblau aus Orthonitrobenzaldehyd. Berichte Dtsch. Chem. Ges. 1882, 15, 2856–2864. [Google Scholar] [CrossRef]
- Steingruber, E. Indigo and Indigo Colorants. Ullmanns Encycl. Ind. Chem. 2004, 19, 55–63. [Google Scholar] [CrossRef]
- Bechtold, T.; Turcanu, A.; Geissler, S.; Ganglberger, E. Process balance and product quality in the production of natural indigo from Polygonum tinctorium Ait. applying low-technology methods. Bioresour. Technol. 2002, 81, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, J.J. Dyer’s Woad. UC IPM Integr. Pest Manag. Home Gard. Landsc. Prof. 2017, 74175, 1–5. Available online: https://www.academia.edu/44904675/Dyers_Woad (accessed on 4 March 2024).
- Farah, K.O.; Tanaka, A.F.; West, N.E. Autecology and Population Biology of Dyers Woad (Isatis tinctoria). Weed Sci. 1988, 36, 186–193. [Google Scholar] [CrossRef]
- Wakeel, A.; Jan, S.A.; Ullah, I.; Shinwari, Z.K.; Xu, M. Solvent polarity mediates phytochemical yield and antioxidant capacity of Isatis tinctoria. PeerJ 2019, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Speranza, J.; Miceli, N.; Taviano, M.F.; Ragusa, S.; Kwiecień, I.; Szopa, A.; Ekiert, H. Isatis tinctoria L. (Woad): A Review of its Botany, Ethnobotanical Uses, Phytochemistry, Biological Activities, and Biotechnological Studies. Plants 2020, 9, 298. [Google Scholar] [CrossRef] [PubMed]
- Bontaz-Carion, J.; Montaut, S.; Goetz, P. Isatis tinctoria L. Brassicaceae: Le pastel des teinturiers. Phytothérapie 2012, 10, 238–244. [Google Scholar] [CrossRef]
- Inventaire National du Patrimoine Naturel. Available online: https://inpn.mnhn.fr/espece/cd_nom/103817 (accessed on 2 July 2024).
- Info Flora. Available online: https://www.infoflora.ch/fr/flore/isatis-tinctoria.html (accessed on 2 July 2024).
- Biolib. Available online: http://www.biolib.de/sturm/flora/index.html (accessed on 6 January 2025).
- Plantes et Botanique. Available online: https://www.plantes-botanique.org/espece_isatis_tinctoria (accessed on 11 March 2024).
- Forest Service of the USDA. Available online: https://www.fs.usda.gov/database/feis/plants/forb/isatin/all.html#Botanical%20description (accessed on 10 July 2025).
- Kizil, S.; Turk, M.; Çakmak, Ö.; Özgüven, M.; Khawar, K.M. Microelement Contents and Fatty Acid Compositions of some Isatis Species Seeds. Not. Bot. Horti Agrobot. Cluj-Napoca 2009, 37, 175–178. Available online: https://www.notulaebotanicae.ro/index.php/nbha/article/view/3115 (accessed on 10 July 2025).
- Dolya, V.S.; Koreshchuk, K.E.; Fursa, N.S.; Golodner, D.N.; Kaminskii, N.A. Oils of three representatives of the family Cruciferae. Chem. Nat. Compd. 1972, 8, 377–378. [Google Scholar] [CrossRef]
- Li, T.; Qu, X.-Y.; Zhang, Q.-A.; Whang, Z.-Z. Ultrasound-assisted extraction and profile characteristics of seed oil from Isatis indigotica Fort. Ind. Crops Prod. 2012, 35, 98–104. [Google Scholar] [CrossRef]
- Angelini, L.G.; Tavarini, S.; Antichi, D.; Bagatta, M.; Matteo, R.; Lazzeri, L. Fatty acid and glucosinolate patterns of seed from Isatis indigotica Fortune as bioproducts for green chemistry. Ind. Crops Prod. 2015, 75, 51–58. [Google Scholar] [CrossRef]
- Miller, R.W.; VanEtten, C.H.; McGrew, C.; Wolff, I.A.; Jones, Q. Seed Meal Amino Acids, Amino Acid Composition of Seed Meals from Forty-One Species of Cruciferae. J. Agric. Food Chem. 1962, 10, 426–430. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, L.; Guan, L.; Zhang, L. Metabolome and transcriptome reveal the biosynthesis of flavonoids and amino acids in Isatis indigotica fruit during development. Physiol. Plant 2024, 176, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mohn, T.; Hamburger, M. Glucosinolate Pattern in Isatis tinctoria and I. indigotica Seeds. Planta Med. 2008, 74, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Zhang, M.; Guo, Q.; Wei, M.; Shi, H.; Wang, T.; Han, Z.; Liu, H.; Liu, C.; Huang, J. Classification of Isatis indigotica Fortune and Isatis tinctoria Linnaeus via comparative analysis of chloroplast genomes. BMC Genom. 2023, 24, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Iba, N. Les Esters D’acides Gras Polyinsaturés d’Isatis tinctoria (pastel): Mise au Point d’une Technique de Séparation—Generalisation à Différentes Huiles Insaturées. Ph.D. Thesis, Institut National Polytechnique de Toulouse, Toulouse, France, 1990. [Google Scholar]
- Gillingham, L.G.; Harris-Janz, S.; Jones, P.J.H. Dietary Monounsaturated Fatty Acids Are Protective Against Metabolic Syndrome and Cardiovascular Disease Risk Factors. Lipids 2011, 46, 209–228. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, B.M.; Vessby, B.; Uusitupa, M.; Berglund, L.; Pedersen, E.; Riccardi, G.; Rivellesse, A.A.; Tapsell, L.; Hermansen, K. Effects of dietary saturated, monounsaturated, and n−3 fatty acids on blood pressure in healthy subjects. Am. J. Clin. Nutr. 2006, 83, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Spataro, G.; Negri, V. Adaptability and variation in Isatis tinctoria L.: A new crop for Europe. Euphytica 2008, 163, 89–102. [Google Scholar] [CrossRef]
- Cosmile Europe. Available online: https://cosmileeurope.eu/fr/inci/ (accessed on 24 May 2024).
- Cosmetic Ingredient Review Safety. Available online: https://cir-reports.cir-safety.org/view-attachment/?id=3a3ac6dd-8c74-ec11-8943-0022482f06a6 (accessed on 28 August 2024).
- Publications Office of the European Union. Available online: https://op.europa.eu/en/publication-detail/-/publication/e332d361-c3e3-4bc0-9fd9-334ebdfbab12/language-en (accessed on 10 June 2024).
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to oleic acid intended to replace saturated fatty acids (SFAs) in foods or diets and maintenance of normal blood LDL-cholesterol concentrations (ID 673, 728, 729, 1302, 4334) and maintenance of normal (fasting) blood concentrations of triglycerides (ID 673, 4334) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2011, 9, 1–17. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to linoleic acid and maintenance of normal blood cholesterol concentrations (ID 489) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2009, 7, 1–12. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to linoleic acid and “molecule precursors regulating cell functions (prostaglandins, leucotrienes)” (ID 488, 4670), maintenance of normal blood LDL-cholesterol concentrations (ID 2899) and protection of the skin from UV-induced damage (ID 3659) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2011, 9, 1–15. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Opinion on the substantiation of health claims related to alpha linolenic acid and maintenance of normal blood cholesterol concentrations (ID 493) and maintenance of normal blood pressure (ID 625) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2009, 7, 1–17. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on health claims already evaluated (ID 215, 568, 674, 712, 1398, 1633, 1974, 4191, 4192, 4193, 4236, 4335, 4698, 4704) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2011, 9, 1–22. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to the replacement of mixtures of saturated fatty acids (SFAs) as present in foods or diets with mixtures of monounsaturated fatty acids (MUFAs) and/or mixtures of polyunsaturated fatty acids (PUFAs), and maintenance of normal blood LDL cholesterol concentrations (ID 621, 1190, 1203, 2906, 2910, 3065) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2011, 9, 1–18. [Google Scholar] [CrossRef]
- Romdhane, M.; Gourdon, C. Investigation in solid–liquid extraction: Influence of ultrasound. Chem. Eng. J. 2002, 87, 11–19. [Google Scholar] [CrossRef]
- Mikolajczak, K.L.; Miwa, T.K.; Earle, F.R.; Wolff, I.A.; Jones, Q. Search for new industrial oils. V. Oils of cruciferae. J. Americ Oil Chem. Soc. 1961, 38, 678–681. [Google Scholar] [CrossRef]
- Jha, A.K.; Sit, N. Extraction of bioactive compounds from plant materials using combination of various novel methods: A review. Trends Food Sci. Technol. 2022, 119, 579–591. [Google Scholar] [CrossRef]
- Ashraf, R.; Ghufran, S.; Akram, S.; Mushtaq, M.; Sultana, B. Cold pressed coriander (Coriandrum sativum L.) seed oil. In Cold Pressed Oils; Ramadan, M.F., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 345–356. [Google Scholar] [CrossRef]
- Prescha, A.; Grajzer, M.; Dedyk, M.; Grajeta, H. The Antioxidant Activity and Oxidative Stability of Cold-Pressed Oils. J. Am. Oil Chem. Soc. 2014, 91, 1291–1301. [Google Scholar] [CrossRef] [PubMed]
- Özcan, M.M.; Al-Juhaimi, F.Y.; Ahmed, I.A.M.; Osman, M.A.; Gassem, M.A. Effect of soxhlet and cold press extractions on the physico-chemical characteristics of roasted and non-roasted chia seed oils. J. Food Meas. Charact. 2019, 13, 648–655. [Google Scholar] [CrossRef]
- International Olive Council COI/T.20/Doc. No 33—Determination of Fatty Acid Methyl Esters by Gas Chromatography. 2017. chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/. Available online: https://www.internationaloliveoil.org/wp-content/uploads/2019/11/COI-T.20-Doc.-No-33-Rev.-1-2017.pdf (accessed on 14 April 2024).
- Demirbas, A. Comparison of transesterification methods for production of biodiesel from vegetable oils and fats. Energy Convers. Manag. 2008, 49, 125–130. [Google Scholar] [CrossRef]
- Milinsk, M.C.; Matsushita, M.; Visentainer, J.V.; de Oliveira, C.C.; de Souza, N.E. Comparative analysis of eight esterification methods in the quantitative determination of vegetable oil fatty acid methyl esters (FAME). J. Braz. Chem. Soc. 2008, 19, 1475–1483. [Google Scholar] [CrossRef]
- Gambert, A.-G.; Niţu, S.; Tămaș, A.; Fanani, M.L.; Dupré, J.; Delepine, C.; Chaveriat, L.; Martin, P.; Rusnac, L. Influence of the Extraction Process on the Characteristics of Romanian Mountain Walnut Oil. Am. J. Plant Sci. 2024, 15, 940–967. [Google Scholar] [CrossRef]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kumar Srivastava, A.; Khare, P.; Kumar Nagar, H.; Raghuwanshi, N.; Srivastava, R. Hydroxyproline: A Potential Biochemical Marker and Its Role in the Pathogenesis of Different Diseases. Curr. Protein Pept. Sci. 2016, 17, 596–602. [Google Scholar] [CrossRef] [PubMed]
- VanEtten, C.H.; Miller, R.W.; Earle, F.R.; Wolffe, I.A.; Jones, Q. Plant Protein Constituents, Hydroxyproline Content of Seed Meals and Distribution of the Amino Acid in Kernel, Seed Coat, and Pericarp. J. Agric. Food Chem. 1961, 9, 433–435. [Google Scholar] [CrossRef]
- Hara, R.; Kino, K. Enzymatic reactions and microorganisms producing the various isomers of hydroxyproline. Appl. Microbiol. Biotechnol. 2020, 104, 4771–4779. [Google Scholar] [CrossRef] [PubMed]
- VanEtten, C.H.; Miller, R.W.; Wolff, I.A. Nutrients in Seed Meals, Amino Acid Composition of Twenty-Seven Selected Seed Meals. J. Agric. Food Chem. 1961, 9, 79–82. [Google Scholar] [CrossRef]
- Levine, M. A New Method for Isolation of Hydroxy-l-proline and l-Proline from Gelatin. J. Biol. Chem. 1959, 234, 1731–1732. [Google Scholar] [CrossRef] [PubMed]
- Spackman, D.H.; Stein, W.H. Moore Stanford. Automatic Recording Apparatus for Use in Chromatography of Amino Acids. Anal. Chem. 1958, 30, 1190–1206. [Google Scholar] [CrossRef]
- Becker, H.C.; Milner, R.T.; Nagel, R.H. A Method for the Determination of Nonprotein Nitrogen in Soybean Meal. Cereal Chem. 1940, 17, 447–457. Available online: https://www.cabidigitallibrary.org/doi/full/10.5555/19401402307 (accessed on 14 April 2024).
- Ms, U.; Ferdosh, S.; Haque Akanda, M.J.; Ghafoor, K.; Rukshana, A.H.; Ali, M.E.; Kamaruzzaman, B.Y.; Fauzi, M.B.; Hadijah, S.; Shaarani, S.; et al. Techniques for the extraction of phytosterols and their benefits in human health: A review. Sep. Sci. Technol. 2018, 53, 2206–2223. [Google Scholar] [CrossRef]
- Garcia-Llatas, G.; Alegría, A.; Barberá, R.; Cilla, A. Current methodologies for phytosterol analysis in foods. Microchem. J. 2021, 168, 1–16. [Google Scholar] [CrossRef]
- Wong, N.D. Epidemiological studies of CHD and the evolution of preventive cardiology. Nat. Rev. Cardiol. 2014, 11, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Piironen, V.; Lindsay, D.G.; Miettinen, T.A.; Toivo, J.; Lampi, A.-M. Plant sterols: Biosynthesis, biological function and their importance to human nutrition. J. Sci. Food Agric. 2000, 80, 939–966. [Google Scholar] [CrossRef]
- Bouic, P.J. The role of phytosterols and phytosterolins in immune modulation: A review of the past 10 years. Curr. Opin. Clin. Nutr. Metab. Care 2001, 4, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Barkizatova, G.; Turgumbayeva, A.; Zhakipbekov, K.; Bekesheva, K.; Arystanov, Z.; Arystanova, T.; Kayupova, F.; Zhumalina, K.; Toxanbayeva, Z.; Ibragimova, A.; et al. Exploring the Pharmacological Potential of Lithospermum officinale L.: A Review of Phytochemicals and Ethnomedicinal Uses. Molecules 2024, 29, 1856. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.B.; Fink, C.S. Phytosterols as Anticancer Dietary Components: Evidence and Mechanism of Action. J. Nutr. 2000, 130, 2127–2130. [Google Scholar] [CrossRef] [PubMed]
- Woyengo, T.A.; Ramprasath, V.R.; Jones, P.J.H. Anticancer effects of phytosterols. Eur. J. Clin. Nutr. 2009, 63, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Burg, V.K.; Grimm, H.S.; Rothhaar, T.L.; Grösgen, S.; Hundsdörfer, B.; Haupenthal, V.J.; Zimmer, V.C.; Mett, J.; Weingärtner, O.; Laufs, U.; et al. Plant sterols the better cholesterol in Alzheimer’s disease? A mechanistical study. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 16072–16087. [Google Scholar] [CrossRef] [PubMed]
- Plaza, M.; Cifuentes, A.; Ibanez, E. In the search of new functional food ingredients from algae. Trends Food Sci. Technol. 2008, 19, 31–39. [Google Scholar] [CrossRef]
- Bin Sayeed, M.; Karim, S.; Sharmin, T.; Morshed, M. Critical Analysis on Characterization, Systemic Effect, and Therapeutic Potential of Beta-Sitosterol: A Plant-Derived Orphan Phytosterol. Medicines 2016, 3, 29. [Google Scholar] [CrossRef] [PubMed]
- Cosmetic Ingredient Review Safety. Available online: https://cir-reports.cir-safety.org/view-attachment/?id=56e8bd9b-8c74-ec11-8943-0022482f06a6 (accessed on 29 August 2024).
- Nazir, S.; Chaudhary, W.; Mubashar, A.; Anjum, I.; Hameed, S.; Azhar, S. Campesterol: A Natural Phytochemical with Anti Inflammatory Properties as Potential Therapeutic Agent for Rheumatoid Arthritis: A Systematic Review: Campesterol: A Natural Phytochemical. Pak. J. Health Sci. 2023, 4, 1–9. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Zhu, L.; Wang, X.; Meng, F.; Xia, L.; Zhang, H. Advances in Stigmasterol on its anti-tumor effect and mechanism of action. Front. Oncol. 2022, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.T.S. Brassicasterol with Dual Anti-Infective Properties against HSV-1 and Mycobacterium tuberculosis, and Cardiovascular Protective Effect: Nonclinical In Vitro and In Silico Assessments. Biomedicines 2020, 8, 132. [Google Scholar] [CrossRef] [PubMed]
- Roche, J.; Alignan, M.; Bouniols, A.; Cerny, M.; Mouloungui, Z.; Vear, F.; Merah, O. Sterol content in sunflower seeds (Helianthus annuus L.) as affected by genotypes and environmental conditions. Food Chem. 2010, 121, 990–995. [Google Scholar] [CrossRef]
- Martinez-Ballesta, M.; Moreno-Fernández, D.A.; Castejon, D.; Ochando, C.; Morandini, P.A.; Carvajal, M. The impact of the absence of aliphatic glucosinolates on water transport under salt stress in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mumm, R.; Burow, M.; Bukovinszkine’Kiss, G.; Kazantzidou, E.; Wittstock, U.; Dicke, M.; Gershenzon, J. Formation of Simple Nitriles upon Glucosinolate Hydrolysis Affects Direct and Indirect Defense Against the Specialist Herbivore, Pieris rapae. J. Chem. Ecol. 2008, 34, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, A.; Dwivedi, A.; du Plessis, J. Sinigrin and Its Therapeutic Benefits. Molecules 2016, 21, 416. [Google Scholar] [CrossRef] [PubMed]
- Michaud, D.S.; Spiegelman, D.; Clinton, S.K.; Rimm, E.B.; Willett, W.C.; Giovannucci, E.L. Fruit and Vegetable Intake and Incidence of Bladder Cancer in a Male Prospective Cohort. J. Natl. Cancer Inst. 1999, 91, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Munday, C.M. Induction of Phase II Detoxification Enzymes in Rats by Plant-Derived Isothiocyanates: Comparison of Allyl Isothiocyanate with Sulforaphane and Related Compounds. J. Agric. Food Chem. 2004, 52, 1867–1871. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W. Chemoprevention by Inducers of Carcinogen Detoxication Enzymes. Environ. Health Perspect. 1997, 105, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Dias, C.; Aires, A.; Saavedra, M. Antimicrobial Activity of Isothiocyanates from Cruciferous Plants against Methicillin-Resistant Staphylococcus aureus (MRSA). Int. J. Mol. Sci. 2014, 15, 19552–19561. [Google Scholar] [CrossRef] [PubMed]
- Nie, L.; Wu, Y.; Dai, Z.; Ma, S. Antiviral activity of Isatidis Radix derived glucosinolate isomers and their breakdown products against influenza A in vitro/ovo and mechanism of action. J. Ethnopharmacol. 2020, 251, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tarini, G.; Melo, A.S.; Fontana, L.F.; Da Silva, E.; Bolanho, B.C.; Moreno, B.P.; Sarragiotto, M.H.; Dias-Arieira, C.R. Aqueous extracts of Crambe abyssinica seed cake: Chemical composition and potential for nematode control. Ind. Crops Prod. 2020, 156, 1–9. [Google Scholar] [CrossRef]
- Ahmad, I.; Fatima, I.; Afza, N.; Malik, A.; Lodhi, M.A.; Choudhary, M.I. Urease and Serine Protease inhibitory alkaloids from Isatis tinctoria. J. Enzyme Inhib. Med. Chem. 2008, 23, 918–921. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Park, H.; Moon, B.; Kim, S. Analysis of glucosinolates and their breakdown products from Mul-kimchis using UPLC-MS/MS. J. Food Compos. Anal. 2024, 125, 1–8. [Google Scholar] [CrossRef]
- McDanell, R.; McLean, A.E.M.; Hanley, A.B.; Heaney, R.K.; Fenwick, G.R. Chemical and biological properties of indole glucosinolates (glucobrassicins): A review. Food Chem. Toxicol. 1988, 26, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.E. Indoles Derived from Glucobrassicin: Cancer Chemoprevention by Indole-3-Carbinol and 3,3′-Diindolylmethane. Front. Nutr. 2021, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Valan Arasu, M.; Lee, M.-K.; Chun, J.-H.; Seo, J.M.; Lee, S.-W.; Al-Dhabi, N.A.; Kim, S.-J. Quantification of glucosinolates, anthocyanins, free amino acids, and vitamin C in inbred lines of cabbage (Brassica oleracea L.). Food Chem. 2014, 145, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Nartea, A.; Fanesi, B.; Pacetti, D.; Lenti, L.; Fiorini, D.; Lucci, P.; Frega, N.G.; Falcone, P.M. Cauliflower by-products as functional ingredient in bakery foods: Fortification of pizza with glucosinolates, carotenoids and phytosterols. Curr. Res. Food Sci. 2023, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dunn, S.E.; LeBlanc, G.A. Hypocholesterolemic properties of plant indoles. Biochem. Pharmacol. 1994, 47, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.P.T.; Stewart, J.; Lopez, M.; Ioannou, I.; Allais, F. Glucosinolates: Natural Occurrence, Biosynthesis, Accessibility, Isolation, Structures, and Biological Activities. Molecules 2020, 25, 4537. [Google Scholar] [CrossRef] [PubMed]
- Mohn, T.; Cutting, B.; Ernst, B.; Hamburger, M. Extraction and analysis of intact glucosinolates--a validated pressurized liquid extraction/liquid chromatography-mass spectrometry protocol for Isatis tinctoria, and qualitative analysis of other cruciferous plants. J. Chromatogr. A 2007, 1166, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Richter, B.E.; Jones, B.A.; Ezzell, J.L.; Porter, N.L.; Avdalovic, N.; Pohl, C. Accelerated Solvent Extraction: A Technique for Sample Preparation. Anal. Chem. 1996, 68, 1033–1039. [Google Scholar] [CrossRef]
- Chen, R.; Wang, X.-J.; Zhang, Y.-Y.; Xing, Y.; Yang, L.; Ni, H.; Li, H.-H. Simultaneous extraction and separation of oil, proteins, and glucosinolates from Moringa oleifera seeds. Food Chem. 2019, 300, 125–162. [Google Scholar] [CrossRef] [PubMed]
- Förster, N.; Ulrichs, C.; Schreiner, M.; Müller, C.T.; Mewis, I. Development of a reliable extraction and quantification method for glucosinolates in Moringa oleifera. Food Chem. 2015, 166, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liang, H.; Yuan, Q. Separation of sinigrin from Indian mustard (Brassica juncea L.) seed using macroporous ion-exchange resin. Korean J. Chem. Eng. 2012, 29, 396–403. [Google Scholar] [CrossRef]
- Sut, S.; Boschiero, I.; Solana, M.; Malagoli, M.; Bertucco, A.; Dall’Acqua, S. Supercritical CO2 Extraction of Eruca sativa Using Cosolvents: Phytochemical Composition by LC-MS Analysis. Molecules 2018, 23, 3240. [Google Scholar] [CrossRef] [PubMed]
- Thies, W. Isolation of Sinigrin and Glucotropaeolin from Cruciferous Seeds. Lipid Fett. 1988, 90, 311–314. [Google Scholar] [CrossRef]
- Rochfort, S.; Caridi, D.; Stinton, M.; Trenerry, V.C.; Jones, R. The isolation and purification of glucoraphanin from broccoli seeds by solid phase extraction and preparative high performance liquid chromatography. J. Chromatogr. A 2006, 1120, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y. High-speed countercurrent chromatography. Nature 1987, 326, 419–420. [Google Scholar] [CrossRef] [PubMed]
- Jaki, B.; Sticher, O.; Veit, M.; Fröhlich, R.; Pauli, G.F. Evaluation of Glucoiberin Reference Material from Iberis amara by Spectroscopic Fingerprinting. J. Nat. Prod. 2002, 65, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Marsh, R.E.; Waser, J. Refinement of the crystal structure of sinigrin. Acta Crystallogr. B 1970, 26, 1030–1037. [Google Scholar] [CrossRef]
- Chidewe, C.; Castillo, U.F.; Sem, D.S. Structural Analysis and Antimicrobial Activity of Chromatographically Separated Fractions of Leaves of Sesamum angustifolium (Oliv.) Engl. J. Biol. Act. Prod. Nat. 2017, 7, 463–474. [Google Scholar] [CrossRef]
- Montaut, S.; Bleeker, R.S.; Jacques, C. Phytochemical constituents of Cardamine diphylla. Can. J. Chem. 2010, 88, 50–55. [Google Scholar] [CrossRef]
- Blažević, I.; Đulović, A.; Čikeš Čulić, V.; Popović, M.; Guillot, X.; Burčul, F.; Rollin, P. Microwave-Assisted versus Conventional Isolation of Glucosinolate Degradation Products from Lunaria annua L. and Their Cytotoxic Activity. Biomolecules 2020, 10, 215. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.; Allart-Simon, I.; De Nicola, G.R.; Iori, R.; Renault, J.-H.; Rollin, P.; Nuzillard, J.-M. Advanced NMR-Based Structural Investigation of Glucosinolates and Desulfoglucosinolates. J. Nat. Prod. 2018, 81, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Grosser, K.; Van Dam, N.M. A Straightforward Method for Glucosinolate Extraction and Analysis with High-pressure Liquid Chromatography (HPLC). J. Vis. Exp. 2017, 121, 1–9. [Google Scholar] [CrossRef]
- Doheny-Adams, T.; Redeker, K.; Kittipol, V.; Bancroft, I.; Hartley, S.E. Development of an efficient glucosinolate extraction method. Plant Methods 2017, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Olsen, C.E.; Huang, X.-C.; Hansen, C.I.C.; Cipollini, D.; Ørgaard, M.; Matthes, A.; Geu-Flores, F.; Koch, M.A.; Agerbirk, N. Glucosinolate diversity within a phylogenetic framework of the tribe Cardamineae (Brassicaceae) unraveled with HPLC-MS/MS and NMR-based analytical distinction of 70 desulfoglucosinolates. Phytochemistry 2016, 132, 33–56. [Google Scholar] [CrossRef] [PubMed]
- Śmiechowska, A.; Bartoszek, A.; Namieśnik, J. Determination of Glucosinolates and Their Decomposition Products—Indoles and Isothiocyanates in Cruciferous Vegetables. Crit. Rev. Anal. Chem. 2010, 40, 202–216. [Google Scholar] [CrossRef]
- Gallaher, C.M.; Gallaher, D.D.; Peterson, S. Development and Validation of a Spectrophotometric Method for Quantification of Total Glucosinolates in Cruciferous Vegetables. J. Agric. Food Chem. 2012, 60, 1358–1362. [Google Scholar] [CrossRef] [PubMed]
- Kopsell, D.A.; Barickman, T.C.; Sams, C.E.; McElroy, J.S. Influence of Nitrogen and Sulfur on Biomass Production and Carotenoid and Glucosinolate Concentrations in Watercress (Nasturtium officinale R. Br.). J. Agric. Food Chem. 2007, 55, 10628–10634. [Google Scholar] [CrossRef] [PubMed]
- Cieślik, E.; Leszczyńska, T.; Filipiak-Florkiewicz, A.; Sikora, E.; Pisulewski, P.M. Effects of some technological processes on glucosinolate contents in cruciferous vegetables. Food Chem. 2007, 105, 976–981. [Google Scholar] [CrossRef]
- Rungapamestry, V.; Duncan, A.J.; Fuller, Z.; Ratcliffe, B. Changes in Glucosinolate Concentrations, Myrosinase Activity, and Production of Metabolites of Glucosinolates in Cabbage (Brassica oleracea Var. capitata) Cooked for Different Durations. J. Agric. Food Chem. 2006, 54, 7628–7634. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, F.; Tomás-Barberán, F.; García-Viguera, C. Glucosinolates and vitamin C content in edible parts of broccoli florets after domestic cooking. Eur. Food Res. Technol. 2002, 215, 310–316. [Google Scholar] [CrossRef]
- Miglio, C.; Chiavaro, E.; Visconti, A.; Fogliano, V.; Pellegrini, N. Effects of Different Cooking Methods on Nutritional and Physicochemical Characteristics of Selected Vegetables. J. Agric. Food Chem. 2008, 56, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Thornalley, P.J. Effect of storage, processing and cooking on glucosinolate content of Brassica vegetables. Food Chem. Toxicol. 2007, 45, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Mawlong, I.; Sujith Kumar, M.S.; Gurung, B.; Singh, K.H.; Singh, D. A simple spectrophotometric method for estimating total glucosinolates in mustard de-oiled cake. Int. J. Food Prop. 2017, 20, 3274–3281. [Google Scholar] [CrossRef]
- Peschel, W.; Dieckmann, W.; Sonnenschein, M.; Plescher, A. High antioxidant potential of pressing residues from evening primrose in comparison to other oilseed cakes and plant antioxidants. Ind. Crops Prod. 2007, 25, 44–54. [Google Scholar] [CrossRef]

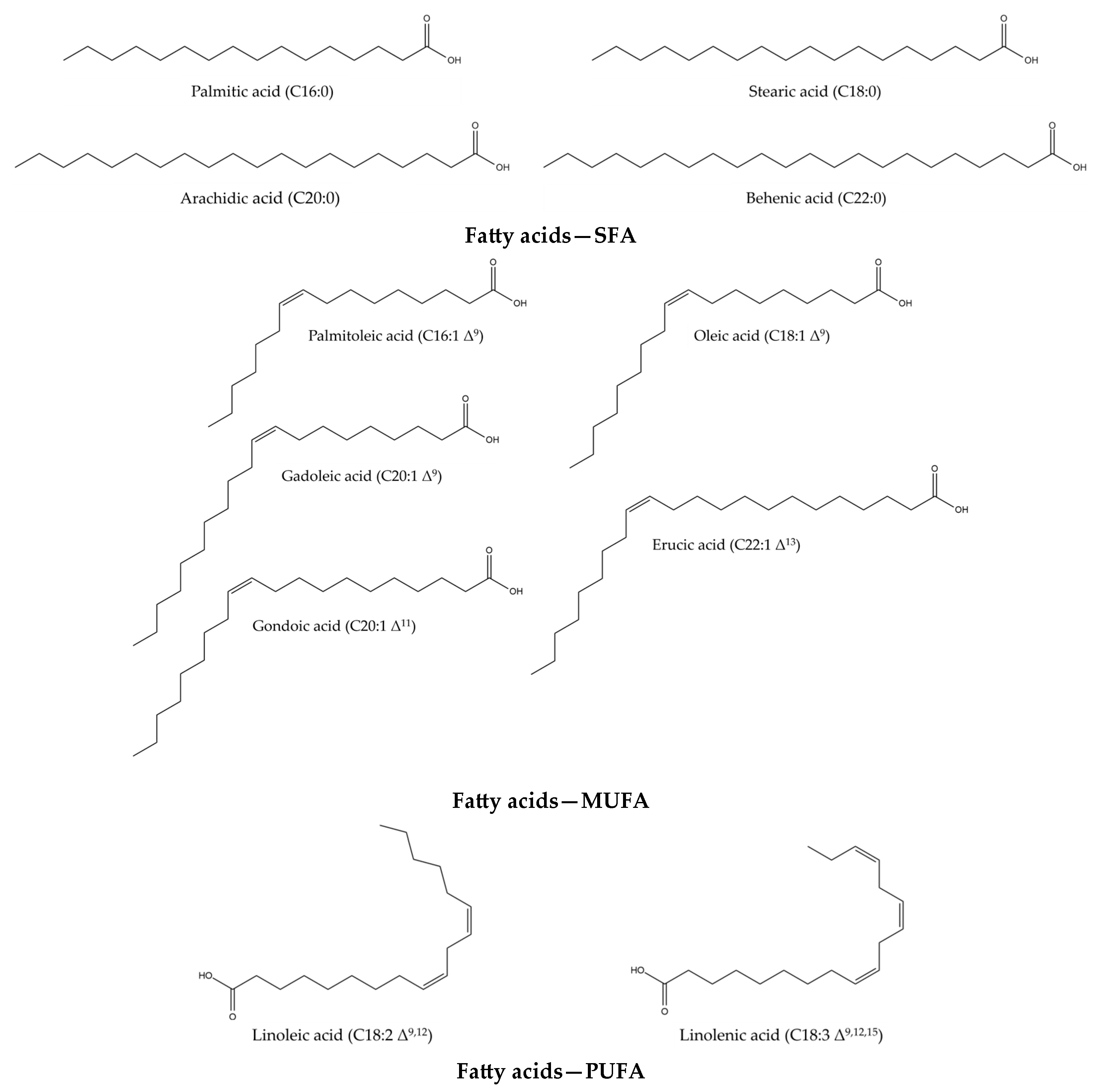
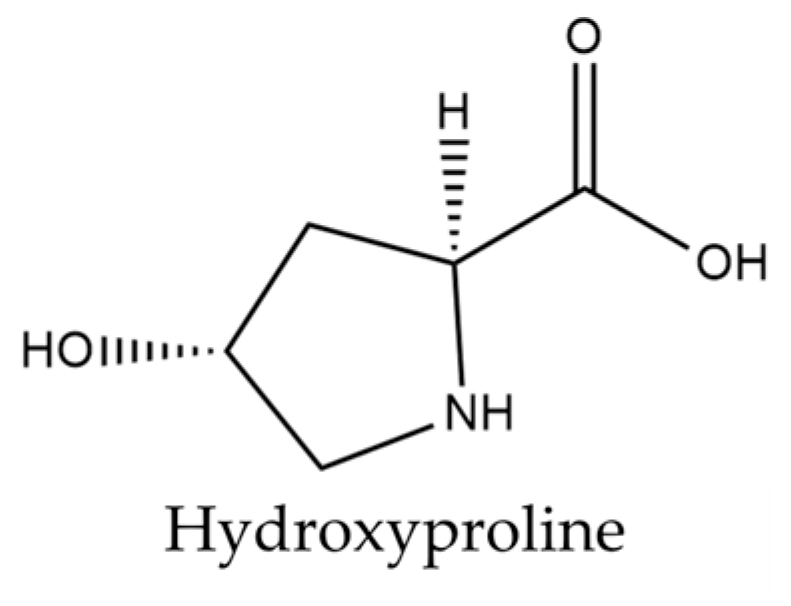
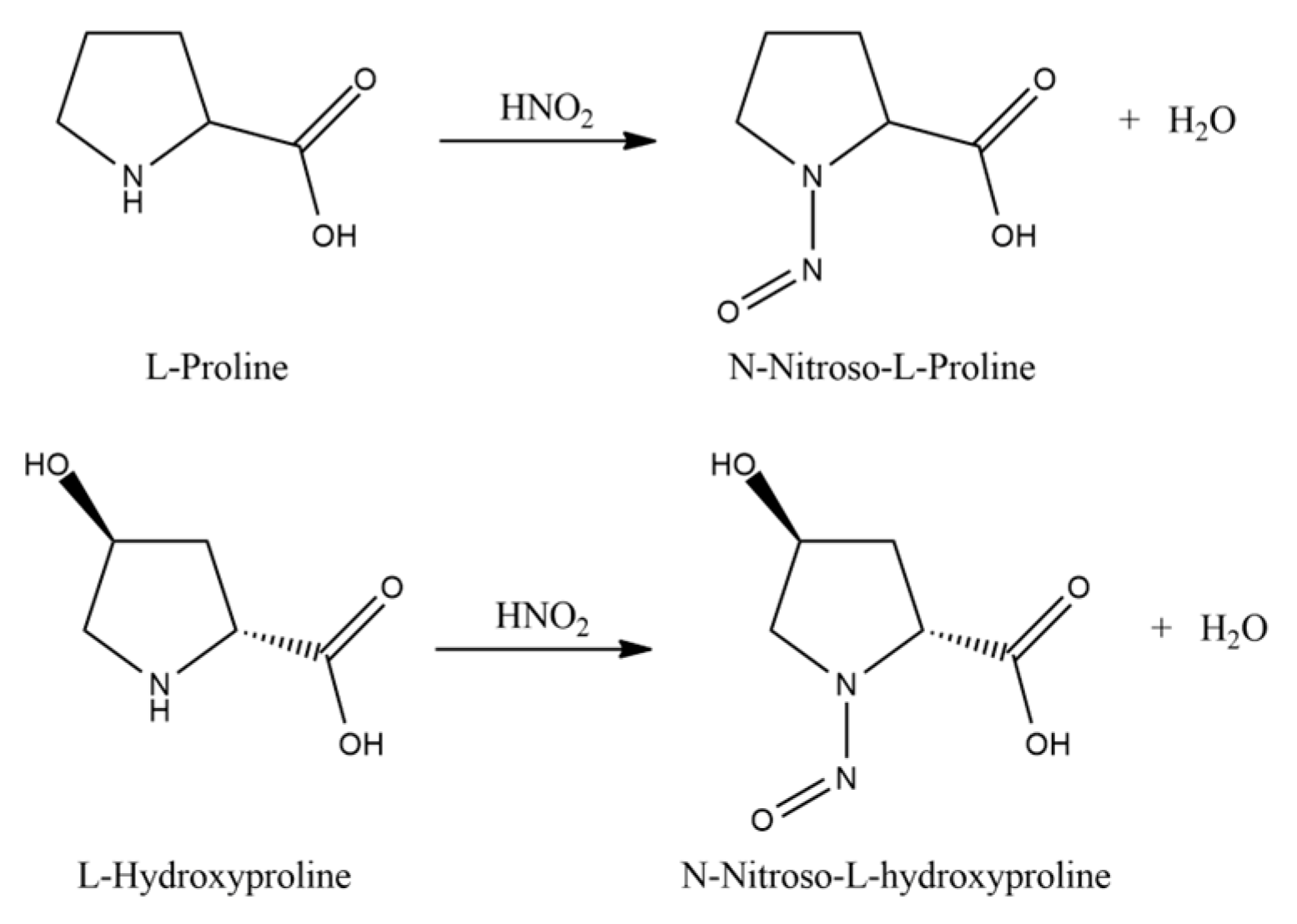
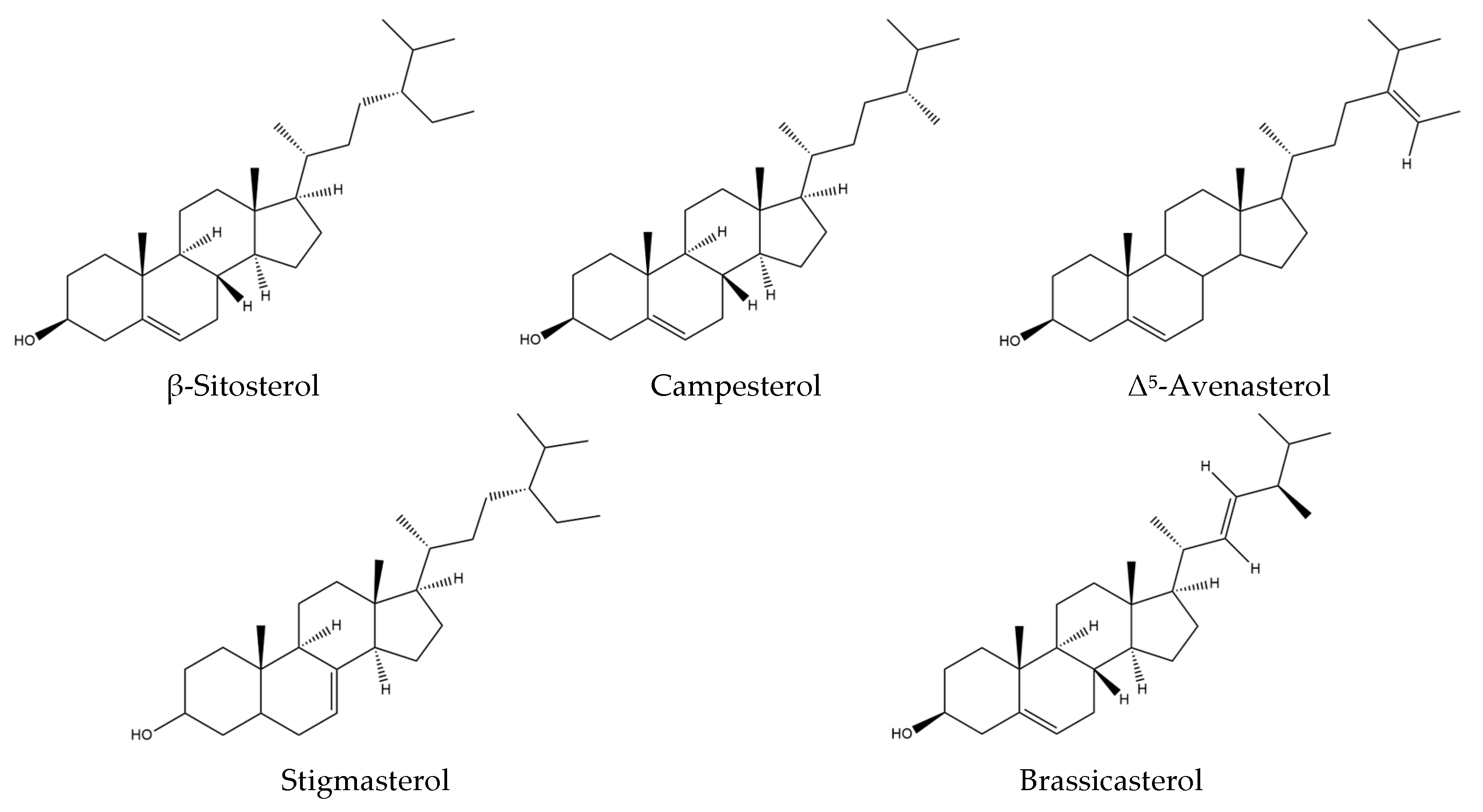
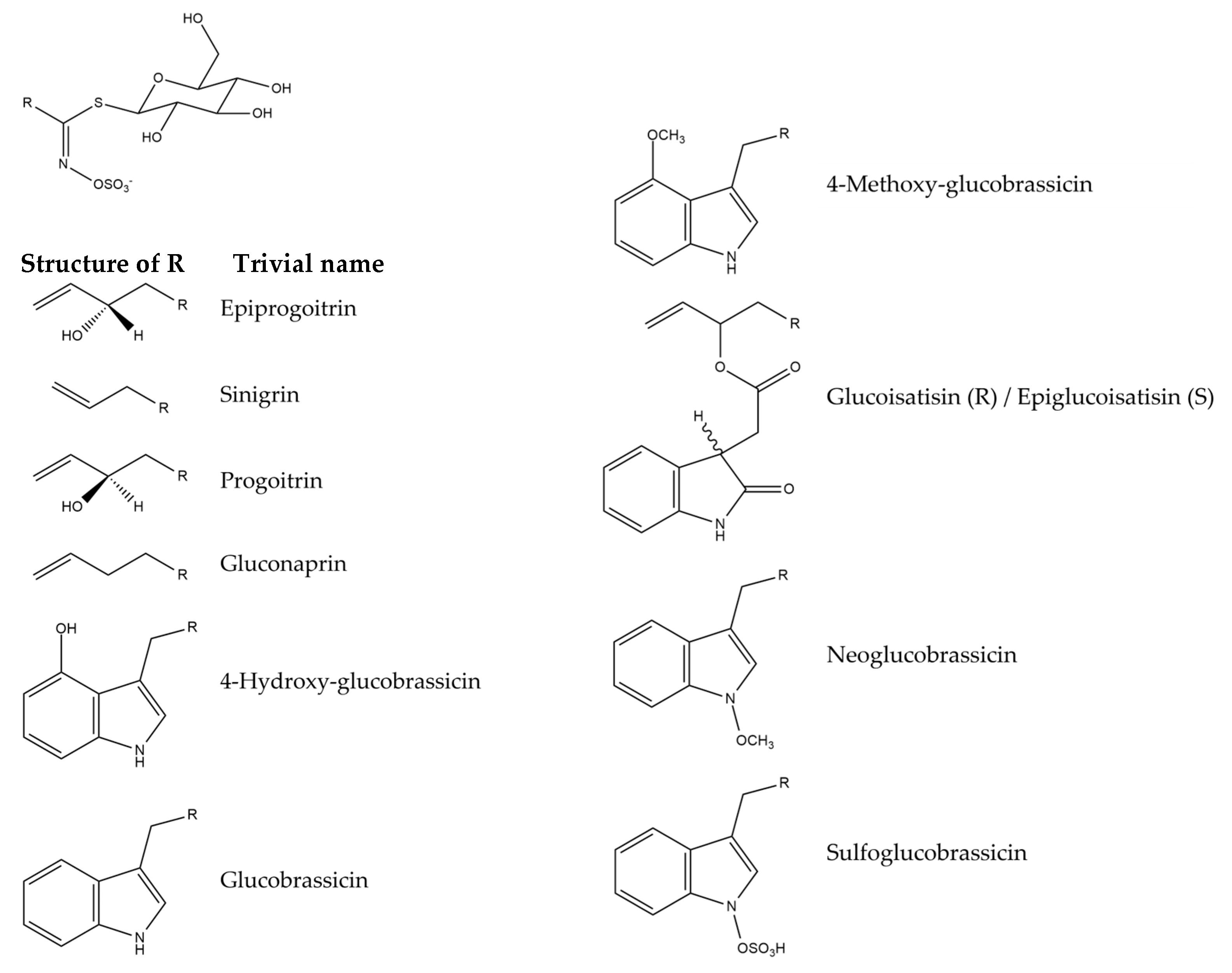
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).