
Rootstock Breeding of Stone Fruits Under Modern Cultivation Regime: Current Status and Perspectives

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
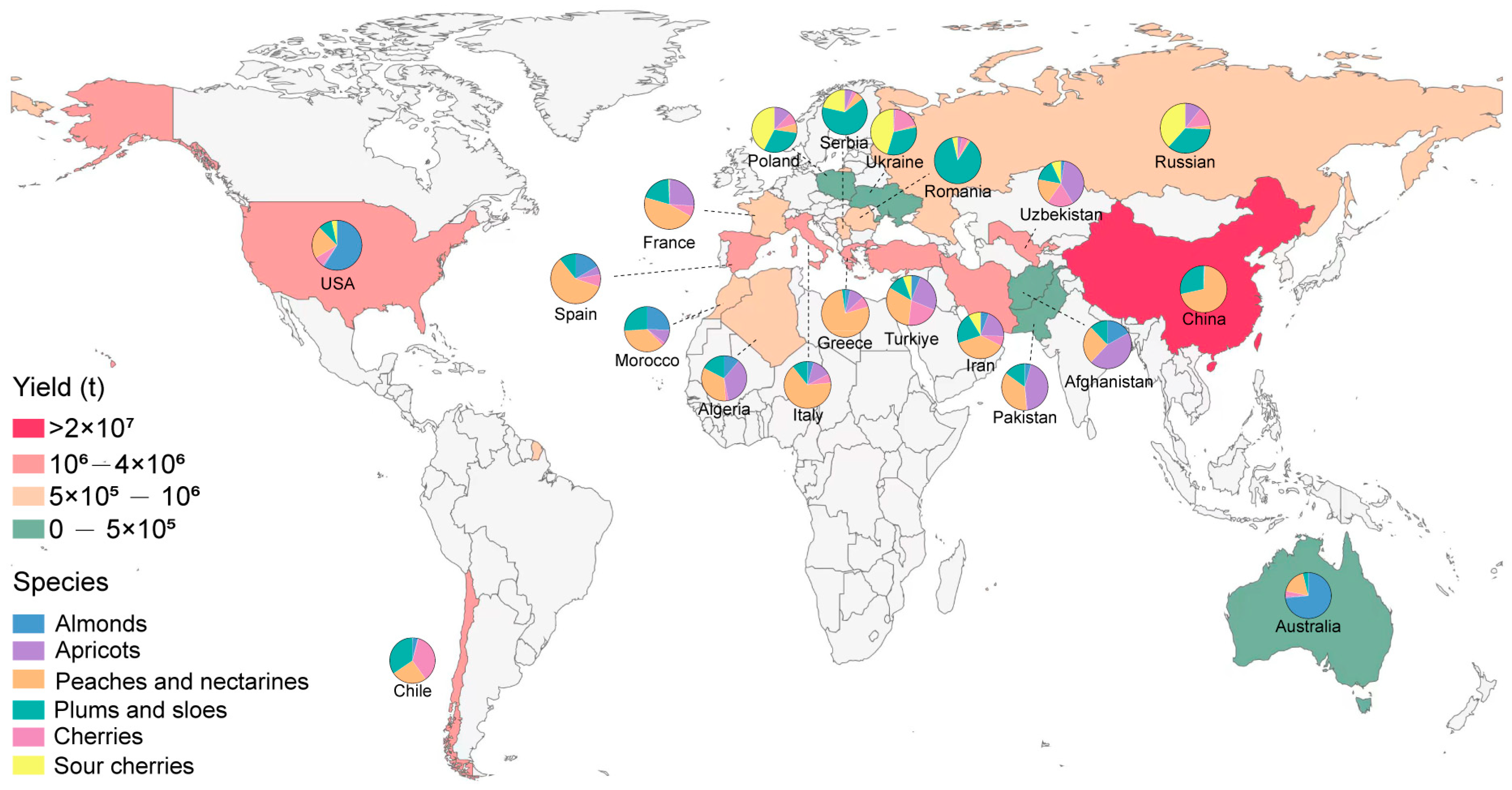
:1. Introduction
2. Modern Trends in Fruit Tree Cultivation Regime and the Demand for Advanced Rootstock Breeding
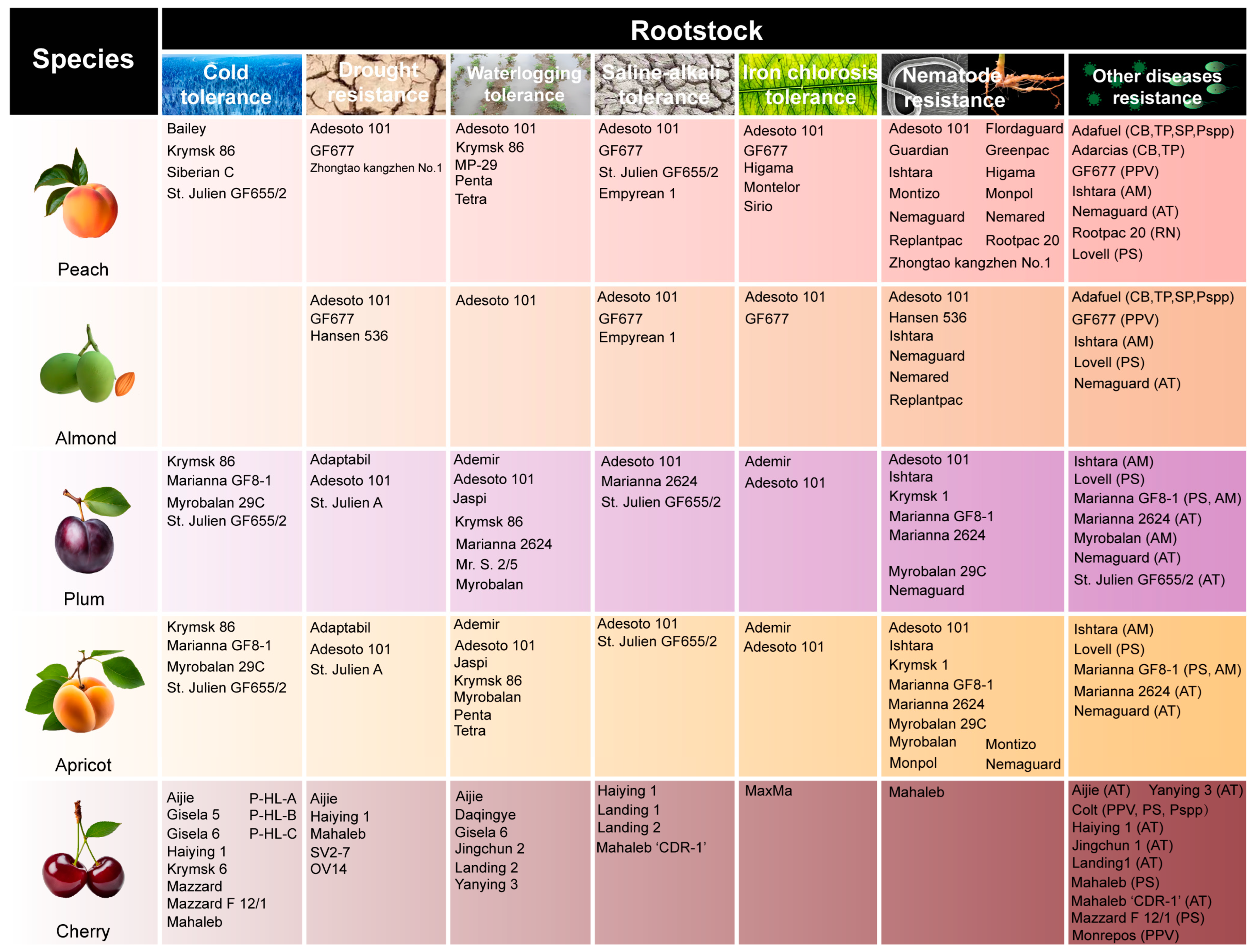
3. Breeding Objectives for Rootstocks of Stone Fruits
3.1. Efficient Clonal Propagation
3.2. Abiotic Stress Tolerance
3.2.1. Temperature Stress
3.2.2. Water Stress
3.2.3. Salinization
3.2.4. Nutritional Stress
3.3. Biotic Stress Tolerance
3.3.1. Viral Diseases
3.3.2. Bacterial Diseases
3.3.3. Fungal Diseases
3.3.4. Pest Damage
3.4. Graft Compatibility
3.4.1. Apricot
3.4.2. Plum
3.4.3. Peach
3.4.4. Almond
3.4.5. Cherry
3.5. Dwarfing
3.5.1. Cherry
3.5.2. Peach and Almond
3.5.3. Plum and Apricot
3.6. Others
4. Breeding Achievements in Rootstocks of Stone Fruit Trees
5. Molecular Breeding Techniques for Rootstocks of Stone Fruit Trees
5.1. Marker-Assisted Breeding
5.2. Genetic Engineering Breeding
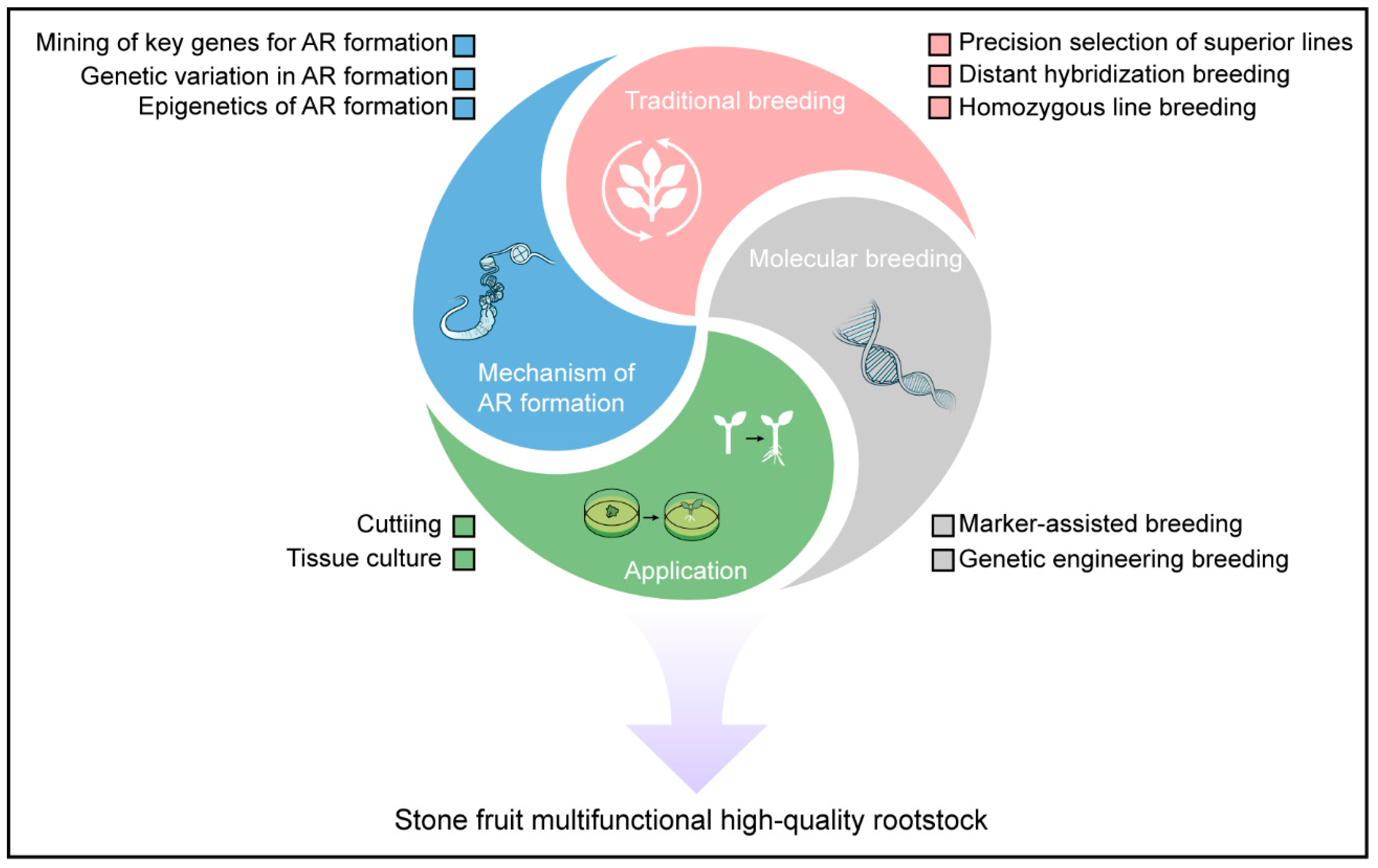
6. Future Perspectives in Stone Fruit Rootstock Research
6.1. Revealing the Molecular Mechanism of Adventitious Root Formation in Rootstocks
6.2. Developing of Efficient Breeding Strategies and New Techniques for Rootstocks
6.3. Optimizing Clonal Propagation Techniques for Rootstocks
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Riva, S.C.; Opara, U.O.; Fawole, O.A. Recent developments on postharvest application of edible coatings on stone fruit: A review. Sci. Hortic. 2020, 262, 109074. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. Food and Agriculture Organization of the United Nations Database (FAOSTAT); Food and Agriculture Organization: Rome, Italy, 2022. [Google Scholar]
- Chen, G.; Boddu, R.; Aadil, R.M. Study on Double-Layer Stereo Ecological Cultivation Technology of Greenhouse Gardening Fruit Trees. J. Food Qual. 2022, 2022, 2655438. [Google Scholar] [CrossRef]
- Ziogas, V. Fruit Growing: Cultivation Strategies for Sustainable Agriculture and Quality Produce. Agronomy 2024, 14, 207. [Google Scholar] [CrossRef]
- Torres-Sánchez, J.; de la Rosa, R.; León, L.; Jiménez-Brenes, F.M.; Kharrat, A.; López-Granados, F. Quantification of dwarfing effect of different rootstocks in ‘Picual’ olive cultivar using UAV-photogrammetry. Precis. Agric. 2021, 23, 178–193. [Google Scholar] [CrossRef]
- Li, W.; Chu, C.; Li, H.; Zhang, H.; Sun, H.; Wang, S.; Wang, Z.; Li, Y.; Foster, T.M.; López-Girona, E.; et al. Near-gapless and haplotype-resolved apple genomes provide insights into the genetic basis of rootstock-induced dwarfing. Nat. Genet. 2024, 56, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Scalisi, A.; O’Connell, M.G.; Stefanelli, D.; Zhou, S.; Pitt, T.; Graetz, D.; Dodds, K.; Han, L.; De Bei, R.; Stanley, J.; et al. Narrow orchard systems for pome and stone fruit—A review. Sci. Hortic. 2024, 338, 113815. [Google Scholar] [CrossRef]
- Liu, R.; Jia, J.; Wang, C.; Wu, Q.; Du, L.; Li, W.; Yang, W.; Ma, J.; Zhang, D.; Xing, L.; et al. Transcriptomic and primary metabolic profiles reveal the mechanism of development and maturation of fuji apple grafted onto different dwarfed intermediate rootstocks. Sci. Hortic. 2025, 343, 114060. [Google Scholar] [CrossRef]
- Korkmaz, K.; Bolat, I.; Uzun, A.; Sahin, M.; Kaya, O. Selection and Molecular Characterization of Promising Plum Rootstocks (Prunus cerasifera L.) among Seedling-Origin Trees. Life 2023, 13, 1476. [Google Scholar] [CrossRef]
- Mayer, N.A.; Ueno, B.; Rickes, T.B.; de Resende, M.V.L.A. Cloning of rootstock selections and Prunus spp. cultivars by softwood cuttings. Sci. Hortic. 2020, 273, 109609. [Google Scholar] [CrossRef]
- Justamante, M.S.; Mhimdi, M.; Molina-Pérez, M.; Albacete, A.; Moreno, M.Á.; Mataix, I.; Pérez-Pérez, J.M. Effects of Auxin (Indole-3-butyric Acid) on Adventitious Root Formation in Peach-Based Prunus Rootstocks. Plants 2022, 11, 913. [Google Scholar] [CrossRef]
- Tsafouros, A.; Frantzeskaki, A.; Assimakopoulou, A.; Roussos, P.A. Spatial and temporal changes of mineral nutrients and carbohydrates in cuttings of four stone fruit rootstocks and their contribution to rooting potential. Sci. Hortic. 2019, 253, 227–240. [Google Scholar] [CrossRef]
- Tsafouros, A.; Roussos, P.A. The possible bottleneck effect of polyamines’ catabolic enzymes in efficient adventitious rooting of two stone fruit rootstocks. J. Plant Physiol. 2020, 244, 152999. [Google Scholar] [CrossRef]
- Blažková, J.; Hlušičková, I. Testing of wood hardiness to winter freezes in selections from progenies of Cerapadus × Prunus avium L. crosses. Hortic. Sci. 2002, 29, 133–142. [Google Scholar] [CrossRef]
- Turhan, E.; Ergin, S. Soluble Sugars and Sucrose-Metabolizing Enzymes Related to Cold Acclimation of Sweet Cherry Cultivars Grafted on Different Rootstocks. Sci. World J. 2012, 2012, 979682. [Google Scholar] [CrossRef]
- Dogan, M.; Bolat, I.; Turan, M.; Kaya, O. Elucidating stress responses in Prunus rootstocks through comprehensive evaluation under drought, heat shock and combined stress conditions. Sci. Hortic. 2025, 339, 113882. [Google Scholar] [CrossRef]
- Cao, F.; Wei, Y.; Wang, X.; Li, Y.; Peng, F. A Study of the Evaluation of the Pecan Drought Resistance of Grafted ‘Pawnee’ Trees From Different Seedling Rootstocks. HortScience 2019, 54, 2139–2145. [Google Scholar] [CrossRef]
- Layne, R.E.C.; Harrison, T.B. ‘Haggith’ Apricot: Rootstock Seed Source. HortScience 1975, 10, 428. [Google Scholar] [CrossRef]
- Ljubojević, M.; Zorić, L.; Maksimović, I.; Dulić, J.; Miodragović, M.; Barać, G.; Ognjanov, V. Anatomically assisted cherry rootstock selection. Sci. Hortic. 2017, 217, 197–208. [Google Scholar] [CrossRef]
- Jia, L.T.; Qin, X.; Lyu, D.G.; Qin, S.J.; Zhang, P. ROS production and scavenging in three cherry rootstocks under short-term waterlogging conditions. Sci. Hortic. 2019, 257, 108647. [Google Scholar] [CrossRef]
- Jalili, S.; Arzani, K.; Prudencio, A.S.; Salazar, J.A.; Martínez-García, P.J.; Bouzari, N.; Martínez-Gómez, P. Integrated Morphological, Physiological and Molecular Analysis of the Drought Response in Cultivated and Wild Prunus L. Subgenera Cerasus Species. Plant Mol. Biol. Report. 2023, 41, 440–453. [Google Scholar] [CrossRef]
- Gerbi, H.; Paudel, I.; Zisovich, A.; Sapir, G.; Ben-Dor, S.; Klein, T. Physiological drought resistance mechanisms in wild species vs. rootstocks of almond and plum. Trees 2021, 36, 669–683. [Google Scholar] [CrossRef]
- Martínez-García, P.J.; Hartung, J.; Pérez de los Cobos, F.; Martínez-García, P.; Jalili, S.; Sánchez-Roldán, J.M.; Rubio, M.; Dicenta, F.; Martínez-Gómez, P. Temporal Response to Drought Stress in Several Prunus Rootstocks and Wild Species. Agronomy 2020, 10, 1383. [Google Scholar] [CrossRef]
- Jurado-Mañogil, C.; Martínez-Melgarejo, P.A.; Martínez-García, P.; Rubio, M.; Hernández, J.A.; Barba-Espín, G.; Diaz-Vivancos, P.; Martínez-García, P.J. Comprehensive study of the hormonal, enzymatic and osmoregulatory response to drought in Prunus species. Sci. Hortic. 2024, 326, 112786. [Google Scholar] [CrossRef]
- McGee, T.; Schaffer, B.; Shahid, M.A.; Chaparro, J.X.; Sarkhosh, A. Carbon and nitrogen metabolism in peach trees on different Prunus rootstocks in response to flooding. Plant Soil 2022, 475, 427–441. [Google Scholar] [CrossRef]
- Ziegler, V.H.; Ploschuk, E.; Weibel, A.; Insausti, P. Short-term responses to flooding stress of three Prunus rootstocks. Sci. Hortic. 2017, 224, 135–141. [Google Scholar] [CrossRef]
- Klumb, E.K.; Braga, E.J.B.; Bianchi, V.J. Differential expression of genes involved in the response of Prunus spp. rootstocks under soil flooding. Sci. Hortic. 2020, 261, 109038. [Google Scholar] [CrossRef]
- Toro, G.; Pimentel, P.; Salvatierra, A. Effective Categorization of Tolerance to Salt Stress through Clustering Prunus Rootstocks According to Their Physiological Performances. Horticulturae 2021, 7, 542. [Google Scholar] [CrossRef]
- Sandhu, D.; Kaundal, A.; Acharya, B.R.; Forest, T.; Pudussery, M.V.; Liu, X.; Ferreira, J.F.S.; Suarez, D.L. Linking diverse salinity responses of 14 almond rootstocks with physiological, biochemical, and genetic determinants. Sci. Rep. 2020, 10, 21087. [Google Scholar] [CrossRef]
- Shao, Y.H.; Cheng, Y.K.; Pang, H.G.; Chang, M.Q.; He, F.; Wang, M.M.; Davis, D.J.; Zhang, S.X.; Betz, O.; Fleck, C.; et al. Investigation of Salt Tolerance Mechanisms Across a Root Developmental Gradient in Almond Rootstocks. Front. Plant Sci. 2021, 11, 595055. [Google Scholar] [CrossRef]
- Paula, B.V.d.; Marques, A.C.R.; Rodrigues, L.A.T.; Souza, R.O.S.d.; Kulmann, M.S.d.S.; Kaminski, J.; Ceretta, C.A.; Melo, G.W.B.d.; Mayer, N.A.; Antunes, L.E.; et al. Morphological and kinetic parameters of the uptake of nitrogen forms in clonal peach rootstocks. Sci. Hortic. 2018, 239, 205–209. [Google Scholar] [CrossRef]
- Chen, Q.J.; Lian, M.; Guo, J.; Zhang, B.B.; Yang, S.K.; Huang, K.X.; Peng, F.T.; Xiao, Y.S. Comparative Transcriptome Analysis of Two Peach Rootstocks Uncovers the Effect of Gene Differential Expression on Nitrogen Use Efficiency. Int. J. Mol. Sci. 2022, 23, 11144. [Google Scholar] [CrossRef] [PubMed]
- Aras, S.; Keles, H.; Bozkurt, E. Physiological and histological responses of peach plants grafted onto different rootstocks under calcium deficiency conditions. Sci. Hortic. 2021, 281, 109967. [Google Scholar] [CrossRef]
- Sun, S.X.; Li, J.; Song, H.Y.; Chen, D.; Tu, M.Y.; Chen, Q.Y.; Jiang, G.L.; Zhou, Z.Q. Comparative transcriptome and physiological analyses reveal key factors in the tolerance of peach rootstocks to iron deficiency chlorosis. 3 Biotech 2022, 12, 38. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, S.; Pinochet, J.; Abadía, A.; Moreno, M.Á.; Gogorcena, Y. Tolerance Response to Iron Chlorosis of Prunus Selections as Rootstocks. HortScience 2008, 43, 304–309. [Google Scholar] [CrossRef]
- Somavilla, L.M.; Simão, D.G.; Tiecher, T.L.; Hammerschimitt, R.K.; de Oliveira, J.M.S.; Mayer, N.A.; Pavanello, E.P.; Trentin, E.; Belles, S.W.; Brunetto, G. Structural changes in roots of peach rootstock cultivars grown in soil with high zinc content. Sci. Hortic. 2018, 237, 1–10. [Google Scholar] [CrossRef]
- Usenik, V.; Marn, M.V. Sugars and organic acids in plum fruit affected by Plum pox virus. J. Sci. Food Agric. 2016, 97, 2154–2158. [Google Scholar] [CrossRef]
- Zhou, J.; Xing, F.; Wang, H.; Li, S. Occurrence, Distribution, and Genomic Characteristics of Plum Pox Virus Isolates from Common Apricot (Prunus armeniaca) and Japanese Apricot (Prunus mume) in China. Plant Dis. 2021, 105, 3474–3480. [Google Scholar] [CrossRef]
- Rubio, M.; Dicenta, F.; Masse, M.; Duval, H. Susceptibility of Prunus rootstocks against Marcus and Dideron isolates of Plum pox virus by graft-inoculation. Ann. Appl. Biol. 2013, 162, 214–220. [Google Scholar] [CrossRef]
- Rubio, M.; Martínez-Gómez, P.; Pinochet, J.; Dicenta, F. Evaluation of resistance to sharka (Plum pox virus) of several Prunus rootstocks. Plant Breed. 2005, 124, 67–70. [Google Scholar] [CrossRef]
- Tomić, J.; Glišić, I.; Milošević, N.; Štampar, F.; Mikulič-Petkovšek, M.; Jakopič, J. Determination of fruit chemical contents of two plum cultivars grafted on four rootstocks. J. Food Compos. Anal. 2022, 105, 103944. [Google Scholar] [CrossRef]
- Dehkordi, A.; Rubio, M.; Babaeian, N.; Albacete, A.; Martínez-Gómez, P. Phytohormone Signaling of the Resistance to Plum pox virus (PPV, Sharka Disease) Induced by Almond (Prunus dulcis (Miller) Webb) Grafting to Peach (P. persica L. Batsch). Viruses 2018, 10, 238. [Google Scholar] [CrossRef] [PubMed]
- Rubio, M.; Martínez-García, P.J.; Martínez-Gómez, P.; Dicenta, F. Plum pox virus (sharka) resistance in peach by grafting ‘Garrigues’ almond as interstock. Sci. Hortic. 2024, 338, 113749. [Google Scholar] [CrossRef]
- Corell-Sierra, J.; Corrêa, R.L.; Gómez, G.G.; Elena, S.F.; Oliveros, J.C.; Rodamilans, B.; Martínez-García, P.J.; Martínez-Gómez, P.; Rubio, M. Almond Grafting for Plum Pox Virus Resistance Triggers Significant Transcriptomic and Epigenetic Shifts in Peaches. Int. J. Mol. Sci. 2024, 26, 248. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Wan, T.; Wu, R.; Zhao, M.; Zhao, Y.; Cai, Y. Resistance analysis of cherry rootstock ‘CDR-1’ (Prunus mahaleb) to crown gall disease. BMC Plant Biol. 2020, 20, 516. [Google Scholar] [CrossRef]
- Thomidis, T.; Exadaktylou, E.; Tsipouridis, C. Susceptibility of five Prunus rootstocks to Agrobacterium tumefaciens. N. Z. J. Crop Hortic. Sci. 2005, 33, 343–345. [Google Scholar] [CrossRef]
- Greco, D.; Aprile, A.; De Bellis, L.; Luvisi, A. Diseases Caused by Xylella fastidiosa in Prunus Genus: An Overview of the Research on an Increasingly Widespread Pathogen. Front. Plant Sci. 2021, 12, 712452. [Google Scholar] [CrossRef]
- Matsumoto, G.O.; Febres, V.J.; Harmon, P.F.; Chaparro, J.X. Survey of Xylella fastidiosa Infection in Prunus Germplasm in Gainesville, FL, USA. HortScience 2023, 58, 819–824. [Google Scholar] [CrossRef]
- Rapicavoli, J.; Ingel, B.; Blanco-Ulate, B.; Cantu, D.; Roper, C. Xylella fastidiosa: An examination of a re-emerging plant pathogen. Mol. Plant Pathol. 2017, 19, 786–800. [Google Scholar] [CrossRef]
- Moll, L.; Giralt, N.; Planas, M.; Feliu, L.; Montesinos, E.; Bonaterra, A.; Badosa, E. Prunus dulcis response to novel defense elicitor peptides and control of Xylella fastidiosa infections. Plant Cell Rep. 2024, 43, 190. [Google Scholar] [CrossRef]
- Beckman, T.G. ‘Sharpe’, a Clonal Plum Rootstock for Peach. HortScience 2008, 43, 2236–2237. [Google Scholar] [CrossRef]
- Beckman, T.G. ‘MP-29’, a Clonal Interspecific Hybrid Rootstock for Peach. HortScience 2012, 47, 128–131. [Google Scholar] [CrossRef]
- Baumgartner, K.; Fujiyoshi, P.; Ledbetter, C.; Duncan, R.; Kluepfel, D.A. Screening Almond Rootstocks for Sources of Resistance to Armillaria Root Disease. HortScience 2018, 53, 4–8. [Google Scholar] [CrossRef]
- Beluzán, F.; Armengol, J.; Abad-Campos, P. Pathogenicity of Oomycete Species to Different Prunus Hybrid Rootstocks. Plant Dis. 2023, 107, 1499–1509. [Google Scholar] [CrossRef] [PubMed]
- Türkölmez, Ş.; Derviş, S.; Çiftçi, O.; Ulubaş Serçe, Ç. First Report of Phytophthora chlamydospora Causing Root and Crown Rot on Almond (Prunus dulcis) Trees in Turkey. Plant Dis. 2016, 100, 1796. [Google Scholar] [CrossRef]
- Browne, G.T.; Ott, N.J.; Forbes, H.; Yaghmour, M.A.; Milliron, L.K. First Report of Phytophthora chlamydospora Causing Crown and Root Rot on Almond in California. Plant Dis. 2020, 104, 2033. [Google Scholar] [CrossRef]
- Browne, G.T. Resistance to Phytophthora Species among Rootstocks for Cultivated Prunus Species. HortScience 2017, 52, 1471–1476. [Google Scholar] [CrossRef]
- Lesmes-Vesga, R.A.; Cano, L.M.; Ritenour, M.A.; Sarkhosh, A.; Chaparro, J.X.; Rossi, L. Rootstocks for Commercial Peach Production in the Southeastern United States: Current Research, Challenges, and Opportunities. Horticulturae 2022, 8, 602. [Google Scholar] [CrossRef]
- Eliwa, G.I.; Hagag, E.S. Approach to New peach rootstocks resistant to root-knot nematodes (Meloidogyne species) selected from local Mit-Ghamer peach cultivar. Sci. Hortic. 2021, 284, 110118. [Google Scholar] [CrossRef]
- Rubio-Cabetas, M.J.; Lecouls, A.C.; Salesses, G.; Bonnet, A.; Esmenjaud, D. Evidence of a new gene for high resistance to Meloidogyne spp. in Myrobalan plum, Prunus cerasifera. Plant Breed. 1998, 117, 567–571. [Google Scholar] [CrossRef]
- Lecouls, A.C.; Rubio-Cabetas, M.J.; Minot, J.C.; Voisin, R.; Bonnet, A.; Salesses, G.; Dirlewanger, E.; Esmenjaud, D. RAPD and SCAR markers linked to the Ma1root-knot nematode resistance gene in Myrobalan plum (Prunus cerasifera Ehr.). Theor. Appl. Genet. 1999, 99, 328–335. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, J.; Li, H.; Luo, D.; Xie, J.; Li, H.; Liu, S.; Zhang, Y.; Chen, L.; Xie, X.; et al. A preliminary study on the root-knot nematode resistance of a cherry plum cultivar Mirabolano 29C. Czech J. Genet. Plant Breed. 2023, 59, 133–140. [Google Scholar] [CrossRef]
- Ji, P.; Liang, C.; Yang, Y.; Wang, R.; Wang, Y.; Yuan, M.; Qiu, Z.; Cheng, Y.; Liu, J.; Li, D. Comparisons of Anatomical Characteristics and Transcriptomic Differences between Heterografts and Homografts in Pyrus L. Plants 2022, 11, 580. [Google Scholar] [CrossRef] [PubMed]
- Reig, G.; Salazar, A.; Zarrouk, O.; Forcada, C.F.I.; Val, J.; Moreno, M.Á. Long-term graft compatibility study of peach-almond hybrid and plum based rootstocks budded with European and Japanese plums. Sci. Hortic. 2019, 243, 392–400. [Google Scholar] [CrossRef]
- Irisarri, P.; Errea, P.; Pina, A. Physiological and Molecular Characterization of New Apricot Cultivars Grafted on Different Prunus Rootstocks. Agronomy 2021, 11, 1464. [Google Scholar] [CrossRef]
- Reig, G.; Zarrouk, O.; Font i Forcada, C.; Moreno, M.Á. Anatomical graft compatibility study between apricot cultivars and different plum based rootstocks. Sci. Hortic. 2018, 237, 67–73. [Google Scholar] [CrossRef]
- Mendelné Pászti, E.; Bujdoso, G.; Ercisli, S.; Hrotkó, K.; Mendel, Á. Apricot Rootstocks with Potential in Hungary. Horticulturae 2023, 9, 720. [Google Scholar] [CrossRef]
- Bouhadida, M.; Casas, A.M.; Gonzalo, M.J.; Arús, P.; Moreno, M.Á.; Gogorcena, Y. Molecular characterization and genetic diversity of Prunus rootstocks. Sci. Hortic. 2009, 120, 237–245. [Google Scholar] [CrossRef]
- Ben Yahmed, J.; Ghrab, M.; Benmoussa, H.; Ben Mimoun, M. Physiological behavior and nutritional status of almond scion-rootstock combinations in a high-density planting system under warm Mediterranean conditions. Sci. Hortic. 2022, 303, 111209. [Google Scholar] [CrossRef]
- Mestre, L.; Reig, G.; Betrán, J.A.; Pinochet, J.; Moreno, M.Á. Influence of peach–almond hybrids and plum-based rootstocks on mineral nutrition and yield characteristics of ‘Big Top’ nectarine in replant and heavy-calcareous soil conditions. Sci. Hortic. 2015, 192, 475–481. [Google Scholar] [CrossRef]
- Reig, G.; Mestre, L.; Betrán, J.A.; Pinochet, J.; Moreno, M.Á. Agronomic and physicochemical fruit properties of ‘Big Top’ nectarine budded on peach and plum based rootstocks in Mediterranean conditions. Sci. Hortic. 2016, 210, 85–92. [Google Scholar] [CrossRef]
- Reig, G.; Garanto, X.; Mas, N.; Iglesias, I. Long-term agronomical performance and iron chlorosis susceptibility of several Prunus rootstocks grown under loamy and calcareous soil conditions. Sci. Hortic. 2020, 262, 109035. [Google Scholar] [CrossRef]
- Reig, G.; Iglesias, I.; Zazurca, L.; Torguet, L.; Martinez, G.; Miarnau, X. Physiological and Agronomical Responses of ‘Vairo’ Almond and ‘Big Top’ Nectarine Cultivars Grafted onto Different Prunus Rootstocks and Grown under Semiarid Mediterranean Conditions. Agronomy 2022, 12, 821. [Google Scholar] [CrossRef]
- Montesinos, Á.; Rubio-Cabetas, M.J.; Grimplet, J. Characterization of Almond Scion/Rootstock Communication in Cultivar and Rootstock Tissues through an RNA-Seq Approach. Plants 2023, 12, 4166. [Google Scholar] [CrossRef]
- Liu, Q.; Zhu, D.; Wang, J.; Zhang, L.; Hong, P.; Gong, Q. The evaluation of sweet cherry rootstocks and their application prospects in the world. Deciduous Fruits 2023, 55, 01–07. [Google Scholar]
- Skočajić, D.; Gašić, U.; Dabić Zagorac, D.; Nešić, M.; Tešić, Ž.; Meland, M.; Fotirić Akšić, M. Analysis of Phenolic Compounds for the Determination of Grafts (in) Compatibility Using In Vitro Callus Cultures of Sato-Zakura Cherries. Plants 2021, 10, 2822. [Google Scholar] [CrossRef]
- Jalali, A.; Moghaddam, E.G.; Marjani, A. Early detection of graft incompatibility in sweet cherry by internode association and callus fusion techniques. Plant Cell Tissue Organ Cult. (PCTOC) 2024, 156, 47. [Google Scholar] [CrossRef]
- Iglesias, I.; Botet, R. The selection of appropriate rootstock and training system towards sustainable production of stone fruits. Italus Hortus 2024, 31, 56–66. [Google Scholar] [CrossRef]
- Clark, J.R.; Finn, C.E. Register of New Fruit and Nut Cultivars List 43. HortScience 2006, 41, 1101–1133. [Google Scholar] [CrossRef]
- Lordan, J.; Zazurca, L.; Maldonado, M.; Torguet, L.; Alegre, S.; Miarnau, X. Horticultural performance of ‘Marinada’ and ‘Vairo’ almond cultivars grown on a genetically diverse set of rootstocks. Sci. Hortic. 2019, 256, 108558. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, M.; Tan, B.; Jiang, Y.; Zheng, X.; Ye, X.; Guo, Z.; Xiong, T.; Wang, W.; Li, J.; et al. A single nucleotide mutation in GID1c disrupts its interaction with DELLA1 and causes a GA-insensitive dwarf phenotype in peach. Plant Biotechnol. J. 2019, 17, 1723–1735. [Google Scholar] [CrossRef]
- Khadivi-Khub, A.; Anjam, K. Prunus scoparia, a suitable rootstock for almond (Prunus dulcis) under drought condition based on vegetative and fruit characteristics. Sci. Hortic. 2016, 210, 220–226. [Google Scholar] [CrossRef]
- Reig, G.; Font i Forcada, C.; Mestre, L.; Betrán, J.A.; Moreno, M.Á. Potential of new Prunus cerasifera based rootstocks for adapting under heavy and calcareous soil conditions. Sci. Hortic. 2018, 234, 193–200. [Google Scholar] [CrossRef]
- Yaman, M.; Uğur, R.; Sümbül, A.; Keçe, Y.; Gönültaş, M.; Ünsal, H.T.; Güneş, A.; Yildiz, E.; Yilmaz, K.U. Determination of fruit characteristics, nutrients and biochemical contents of Transvalia (Prunus persica L.) peach cultivar grafted on different clonal rootstocks obtained by selection and hybridization. Sci. Hortic. 2024, 330, 113093. [Google Scholar] [CrossRef]
- Font i Forcada, C.; Gogorcena, Y.; Moreno, M.A. Fruit sugar profile and antioxidants of peach and nectarine cultivars on almond×peach hybrid rootstocks. Sci. Hortic. 2013, 164, 563–572. [Google Scholar] [CrossRef]
- Giorgi, M.; Capocasa, F.; Scalzo, J.; Murri, G.; Battino, M.; Mezzetti, B. The rootstock effects on plant adaptability, production, fruit quality, and nutrition in the peach (cv. ‘Suncrest’). Sci. Hortic. 2005, 107, 36–42. [Google Scholar] [CrossRef]
- Iglesias, I.; Giné-Bordonaba, J.; Garanto, X.; Reig, G. Rootstock affects quality and phytochemical composition of ‘Big Top’ nectarine fruits grown under hot climatic conditions. Sci. Hortic. 2019, 256, 108586. [Google Scholar] [CrossRef]
- López-Ortega, G.; García-Montiel, F.; Bayo-Canha, A.; Frutos-Ruiz, C.; Frutos-Tomás, D. Rootstock effects on the growth, yield and fruit quality of sweet cherry cv. ‘Newstar’ in the growing conditions of the Region of Murcia. Sci. Hortic. 2016, 198, 326–335. [Google Scholar] [CrossRef]
- Milošević, T.; Milošević, N.; Mladenović, J. Combining fruit quality and main antioxidant attributes in the sour cherry: The role of new clonal rootstock. Sci. Hortic. 2020, 265, 109236. [Google Scholar] [CrossRef]
- Bujdosó, G.; Magyar, L.; Hrotkó, K. Long term evaluation of growth and cropping of sweet cherry (Prunus avium L.) varieties on different rootstocks under Hungarian soil and climatic conditions. Sci. Hortic. 2019, 256, 108613. [Google Scholar] [CrossRef]
- Hernández, F.; Pinochet, J.; Moreno, M.A.; Martínez, J.J.; Legua, P. Performance of Prunus rootstocks for apricot in Mediterranean conditions. Sci. Hortic. 2010, 124, 354–359. [Google Scholar] [CrossRef]
- Kumar, A.; Rathore, J.; Waida, U.I.; Sharma, A.; Nagar, P.; Mir, M. Rootstocks of Stone Fruit Crops. In Production Technology of Stone Fruits; Springer: Singapore, 2021; pp. 131–169. [Google Scholar]
- Gruppe, W. An overview of the cherry rootstock breeding programme. Acta Hortic. 1985, 169, 189–198. [Google Scholar] [CrossRef]
- Zhu, D.; Wang, J.; Hong, P.; Tan, Y.; Liu, Q. A new tetraploid sweet cherry dwarf rootstock cultivar ‘Aijie’. Acta Hortic. Sin. 2020, 47, 2895–2896. [Google Scholar]
- Liu, L.; Zhang, J.; Dun, B.; Li, M.; Li, C. Breeding of a new sweet cherry rootstock cultivar ‘Haiying 1’. J. Fruit Sci. 2014, 31, 210–211. [Google Scholar]
- Zhang, X.; Yan, G.; Zhou, Y.; Wang, J.; Duan, X.; Wu, C.; Zhang, K. A new sweet cherry rootstock cultivar ‘Jingchun 2’. Acta Hortic. Sin. 2022, 49, 31–32. [Google Scholar]
- Zhang, X.; Yan, G.; Zhou, Y.; Wang, J.; Duan, X.; Zhang, K. A new sweet cherry rootstock cultivar ‘Jingchun 1’. Acta Hortic. Sin. 2021, 48, 2787–2788. [Google Scholar]
- Zhang, X.; Yan, G.; Zhou, Y.; Wang, J.; Duan, X.; Zhang, K. A new sweet cherry rootstock cultivar ‘Landing 3’. Acta Hortic. Sin. 2021, 48, 2789–2790. [Google Scholar]
- Zhang, X.; Zhang, F.X.; Sun, Q.T.; Li, Y.J.; Tian, C.P.; Li, F.D.; Wang, Y.X.; Li, S.P. Selection and cultivation of sweet cherry rootstock ‘Yanying No. 3’. Yantai Fruits 2018, 4, 25–27. [Google Scholar]
- Wang, L.-R.; Wang, X.-W.; Zhu, G.-R.; Fang, W.-C.; Chen, C.-W.; Cao, K.; Li, Y.; Wu, J.-L.; Wang, L.-L.; Niu, P. A replantation disease resistant new peach rootstock cultivar Zhong Tao Kang Zhen No.1. J. Fruit Sci. 2023, 40, 1766–1770. [Google Scholar]
- Bowman, K.D.; McCollum, G.; Albrecht, U. SuperSour: A New Strategy for Breeding Superior Citrus Rootstocks. Front. Plant Sci. 2021, 12, 741009. [Google Scholar] [CrossRef]
- Vahdati, K.; Sarikhani, S.; Arab, M.M.; Leslie, C.A.; Dandekar, A.M.; Aletà, N.; Bielsa, B.; Gradziel, T.M.; Montesinos, Á.; Rubio-Cabetas, M.J.; et al. Advances in Rootstock Breeding of Nut Trees: Objectives and Strategies. Plants 2021, 10, 2234. [Google Scholar] [CrossRef]
- Lambert, P.; Campoy, J.A.; Pacheco, I.; Mauroux, J.-B.; Da Silva Linge, C.; Micheletti, D.; Bassi, D.; Rossini, L.; Dirlewanger, E.; Pascal, T.; et al. Identifying SNP markers tightly associated with six major genes in peach [Prunus persica (L.) Batsch] using a high-density SNP array with an objective of marker-assisted selection (MAS). Tree Genet. Genomes 2016, 12, 121. [Google Scholar] [CrossRef]
- Soriano, J.M.; Domingo, M.L.; Zuriaga, E.; Romero, C.; Zhebentyayeva, T.; Abbott, A.G.; Badenes, M.L. Identification of simple sequence repeat markers tightly linked to plum pox virus resistance in apricot. Mol. Breed. 2011, 30, 1017–1026. [Google Scholar] [CrossRef]
- Zuriaga, E.; Soriano, J.M.; Zhebentyayeva, T.; Romero, C.; Dardick, C.; Cañizares, J.; Badenes, M.L. Genomic analysis reveals MATH gene(s) as candidate(s) for Plum pox virus (PPV) resistance in apricot (Prunus armeniaca L.). Mol. Plant Pathol. 2013, 14, 663–677. [Google Scholar] [CrossRef] [PubMed]
- Rubio, M.; Ruiz, D.; Egea, J.; Martínez-Gómez, P.; Dicenta, F. Opportunities of marker-assisted selection for Plum pox virus resistance in apricot breeding programs. Tree Genet. Genomes 2014, 10, 513–525. [Google Scholar] [CrossRef]
- Zuriaga, E.; Romero, C.; Blanca, J.M.; Badenes, M.L. Resistance to Plum Pox Virus (PPV) in apricot (Prunus armeniaca L.) is associated with down-regulation of two MATHd genes. BMC Plant Biol. 2018, 18, 25. [Google Scholar] [CrossRef]
- Polo-Oltra, Á.; Romero, C.; López, I.; Badenes, M.; Zuriaga, E. Cost-Effective and Time-Efficient Molecular Assisted Selection for PPV Resistance in Apricot Based on ParPMC2 Allele-Specific PCR. Agronomy 2020, 10, 1292. [Google Scholar] [CrossRef]
- Gürcan, K.; Çetinsağ, N.; Pınar, H.; Macit, T. Molecular and biological assessment reveals sources of resistance to Plum pox virus—Turkey strain in Turkish apricot (Prunus armeniaca) germplasm. Sci. Hortic. 2019, 252, 348–353. [Google Scholar] [CrossRef]
- Rubio, M.; Martínez-Gómez, P.; Dicenta, F. Apricot breeding for multiple resistance to Plum pox virus and Apple chlorotic leaf spot virus. Sci. Hortic. 2023, 309, 111706. [Google Scholar] [CrossRef]
- Nicolás-Almansa, M.; Ruiz, D.; Salazar, J.A.; Guevara, A.; Cos, J.; Martínez-Gómez, P.; Rubio, M. Phenotypic and molecular characterization of new interspecific Japanese plum × apricot hybrids (plumcots). Sci. Hortic. 2023, 318, 112131. [Google Scholar] [CrossRef]
- Mayer, N.A.; Ueno, B.; Nava, G.; Bianchi, V.J.; Nicolao, G.; Roth, F.M.; Antunes, L.E.C. Performance of clonal rootstocks for peach and own-rooted ‘Maciel’ trees in area with a history of the PTSL syndrome. Sci. Hortic. 2024, 328, 112946. [Google Scholar] [CrossRef]
- Blenda, A.V.; Verde, I.; Georgi, L.L.; Reighard, G.L.; Forrest, S.D.; Muñoz-Torres, M.; Baird, W.V.; Abbott, A.G. Construction of a genetic linkage map and identification of molecular markers in peach rootstocks for response to peach tree short life syndrome. Tree Genet. Genomes 2007, 3, 341–350. [Google Scholar] [CrossRef]
- Meza, P.; Soto, B.; Rojas, L.; Esmenjaud, D. Identification of Meloidogyne Species from the Central Valley of Chile and Interaction with Stone Fruit Rootstocks. Plant Dis. 2016, 100, 1358–1363. [Google Scholar] [CrossRef] [PubMed]
- Maquilan, M.A.D.; Olmstead, M.A.; Olmstead, J.W.; Dickson, D.W.; Chaparro, J.X. Genetic analyses of resistance to the peach root-knot nematode (Meloidogyne floridensis) using microsatellite markers. Tree Genet. Genomes 2018, 14, 47. [Google Scholar] [CrossRef]
- Duval, H.; Heurtevin, L.; Dlalah, N.; Caravel, C.; Callot, C.; Van Ghelder, C. Identification and Expression of the peach TNL RMia genes for the Resistance to the Root-knot Nematode Meloidogyne incognita. bioRxiv 2024. [Google Scholar] [CrossRef]
- Duval, H.; Van Ghelder, C.; Portier, U.; Confolent, C.; Meza, P.; Esmenjaud, D. New Data Completing the Spectrum of theMa, RMia, andRMjaGenes for Resistance to Root-Knot Nematodes (Meloidogyne spp.) in Prunus. Phytopathology 2019, 109, 615–622. [Google Scholar] [CrossRef]
- Lu, Z.; Niu, L.; Chagné, D.; Cui, G.; Pan, L.; Foster, T.; Zhang, R.; Zeng, W.; Wang, Z. Fine mapping of the temperature-sensitive semi-dwarf (Tssd) locus regulating the internode length in peach (Prunus persica). Mol. Breed. 2016, 36, 20. [Google Scholar] [CrossRef]
- Cantín, C.M.; Arús, P.; Eduardo, I. Identification of a new allele of the Dw gene causing brachytic dwarfing in peach. BMC Res. Notes 2018, 11, 386. [Google Scholar] [CrossRef]
- García-Almodóvar, R.C.; Clemente-Moreno, M.J.; Díaz-Vivancos, P.; Petri, C.; Rubio, M.; Padilla, I.M.G.; Ilardi, V.; Burgos, L. Greenhouse evaluation confirms in vitro sharka resistance of genetically engineered h-UTR/P1 plum plants. Plant Cell Tissue Organ Cult. (PCTOC) 2014, 120, 791–796. [Google Scholar] [CrossRef]
- Sidorova, T.; Pushin, A.; Miroshnichenko, D.; Dolgov, S. Generation of Transgenic Rootstock Plum ((Prunus pumila L. × P. salicina Lindl.) × (P. cerasifera Ehrh.)) Using Hairpin-RNA Construct for Resistance to the Plum pox virus. Agronomy 2017, 8, 2. [Google Scholar] [CrossRef]
- Sidorova, T.; Mikhailov, R.; Pushin, A.; Miroshnichenko, D.; Dolgov, S. Agrobacterium-Mediated Transformation of Russian Commercial Plum cv. “Startovaya” (Prunus domestica L.) With Virus-Derived Hairpin RNA Construct Confers Durable Resistance to PPV Infection in Mature Plants. Front. Plant Sci. 2019, 10, 286. [Google Scholar] [CrossRef]
- Alburquerque, N.; Pérez-Caselles, C.; Faize, L.; Ilardi, V.; Burgos, L. Trans-grafting plum pox virus resistance from transgenic plum rootstocks to apricot scions. Front. Plant Sci. 2023, 14, 1216217. [Google Scholar] [CrossRef] [PubMed]
- Mourenets, L.; Pushin, A.; Timerbaev, V.; Khmelnitskaya, T.; Gribkov, E.; Andreev, N.; Dolgov, S. Effect of Gene Silencing of Translation Initiation Factors eIF(iso)4G and eIF(iso)4E on Sour Cherry Rootstock Resistance to Sharka Disease. Int. J. Mol. Sci. 2022, 24, 360. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.J.; Xu, L.; Tan, Y.; Wei, H.R. Development of genetically modified sweet cherry rootstock ‘Gisela 6’ with overexpression of PcMPK3-HA gene by Agrobacterium-mediated genetic transformation. Plant Cell Tissue Organ Cult. 2022, 151, 375–384. [Google Scholar] [CrossRef]
- Jedličková, V.; Štefková, M.; Sánchez López, J.F.; Grimplet, J.; Rubio Cabetas, M.J.; Robert, H.S. Genome editing in almond using hairy root transformation system. Plant Cell Tissue Organ Cult. (PCTOC) 2024, 159, 74. [Google Scholar] [CrossRef]
- Wasmer, M. Roads Forward for European GMO Policy—Uncertainties in Wake of ECJ Judgment Have to be Mitigated by Regulatory Reform. Front. Bioeng. Biotechnol. 2019, 7, 132. [Google Scholar] [CrossRef]
- Escajedo San-Epifanio, L.; Filibi, I.; Lasa López, A.; Puigdomènech, P.; Uncetabarrenechea Larrabe, J. Possible EU futures for CRISPR-edited plants: Little margin for optimism? Front. Plant Sci. 2023, 14, 1141455. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ling, J.; Yu, W.; Yang, L.; Zhang, J.; Jiang, F.; Zhang, M.; Wang, Y.; Sun, H. Rootstock Breeding of Stone Fruits Under Modern Cultivation Regime: Current Status and Perspectives. Plants 2025, 14, 1320. https://doi.org/10.3390/plants14091320
Ling J, Yu W, Yang L, Zhang J, Jiang F, Zhang M, Wang Y, Sun H. Rootstock Breeding of Stone Fruits Under Modern Cultivation Regime: Current Status and Perspectives. Plants. 2025; 14(9):1320. https://doi.org/10.3390/plants14091320
Chicago/Turabian StyleLing, Juanjuan, Wenjian Yu, Li Yang, Junhuan Zhang, Fengchao Jiang, Meiling Zhang, Yuzhu Wang, and Haoyuan Sun. 2025. "Rootstock Breeding of Stone Fruits Under Modern Cultivation Regime: Current Status and Perspectives" Plants 14, no. 9: 1320. https://doi.org/10.3390/plants14091320
APA StyleLing, J., Yu, W., Yang, L., Zhang, J., Jiang, F., Zhang, M., Wang, Y., & Sun, H. (2025). Rootstock Breeding of Stone Fruits Under Modern Cultivation Regime: Current Status and Perspectives. Plants, 14(9), 1320. https://doi.org/10.3390/plants14091320