Quantifying the Effects of Photoperiod, Temperature and Daily Irradiance on Flowering Time of Soybean Isolines

Abstract
:1. Introduction
2. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isoline | Maturity genotype † | Photoperiod coefficient (standard error) | ||||
|---|---|---|---|---|---|---|
| Additive (°C−1 d−1 h−1) | Multiplicative (h−1) | |||||
| Dep. | Ind. | Dep. | Ind. | |||
| OT 94-47 | H | e1e2e3e4e5e7 | 1.32 (0.12) | 1.53 (0.18) | 0.0206 (0.0057) | 0.0193 (0.0098) |
| OT 89-5 | H | e1e2e3e4e5E7 | 1.66 (0.13) | 1.94 (0.23) | 0.0416 (0.0068) | 0.0478 (0.0116) |
| OT 93-26 ‡ | H | E1e2e3e4e5E7 | 2.32 (0.09) | 2.52 (0.15) | 0.0806 (0.0048) | 0.0822 (0.0069) |
| OT 93-28 ‡ | H | E1e2e3e4e5E7 | 2.38 (0.09) | 2.57 (0.15) | 0.0839 (0.0045) | 0.0859 (0.0070) |
| OT 94-41 | H | e1e2E3e4e5E7 | 2.10 (0.09) | 2.28 (0.14) | 0.0675 (0.0050) | 0.0686 (0.0079) |
| L92-21 | C | e1e2E3e4e5E7 | 1.90 (0.12) | 2.17 (0.18) | 0.0543 (0.0063) | 0.0609 (0.0092) |
| L62-667 | H | e1e2e3E4e5E7 | 2.00 (0.12) | 2.25 (0.18) | 0.0624 (0.0067) | 0.0663 (0.0104) |
| L71-920 | C | e1e2e3E4e5E7 | 2.06 (0.10) | 2.23 (0.16) | 0.0657 (0.0054) | 0.0650 (0.0084) |
| L71-802 | H | E1e2e3E4e5E7 | 3.17 (0.05) | 3.28 (0.07) | 0.1304 (0.0029) | 0.1290 (0.0046) |
| L80-5914 | C | E1e2e3E4e5E7 | 3.13 (0.06) | 3.18 (0.07) | 0.1283 (0.0036) | 0.1234 (0.0040) |
| L84-307 | H | e1E2e3E4e5E7 | 2.71 (0.10) | 2.81 (0.11) | 0.1034 (0.0052) | 0.1011 (0.0064) |
| L63-2404 | C | e1E2e3E4e5E7 | 2.65 (0.07) | 2.76 (0.09) | 0.0996 (0.0038) | 0.0972 (0.0052) |
| Harosoy | H | e1e2E3E4e5E7 | 2.25 (0.12) | 2.44 (0.18) | 0.0772 (0.0068) | 0.0780 (0.0090) |
| L84-307 | H | e1e2e3E4E5E7 | 2.10 (0.12) | 2.28 (0.19) | 0.0671 (0.0067) | 0.0686 (0.0103) |
| L74-441 | H | E1E2e3E4e5E7 | 3.63 (0.05) | 3.73 (0.07) | 0.1567 (0.0027) | 0.1561 (0.0043) |
| L67-2324 | H | E1e2E3E4e5E7 | 3.79 (0.06) | 3.82 (0.04) | 0.1662 (0.0029) | 0.1615 (0.0031) |
| L66-432 | C | E1e2E3E4e5E7 | 3.66 (0.09) | 3.75 (0.09) | 0.1588 (0.0050) | 0.1569 (0.0054) |
| L64-4584 | H | e1E2E3E4e5E7 | 3.17 (0.06) | 3.19 (0.08) | 0.1311 (0.0034) | 0.1243 (0.0043) |
| Clark | C | e1E2E3E4e5E7 | 3.14 (0.07) | 3.13 (0.09) | 0.1294 (0.0041) | 0.1203 (0.0054) |
| L64-4830 | H | e1e2E3E4E5E7 | 3.22 (0.10) | 3.12 (0.12) | 0.1345 (0.0051) | 0.1194 (0.0068) |
| L94-1110 | C | e1e2E3E4E5E7 | 2.72 (0.15) | 2.70 (0.16) | 0.1042 (0.0082) | 0.0930 (0.0092) |
| L71L-3004 | H | E1E2E3E4e5E7 | 4.44 (0.05) | 4.47 (0.07) | 0.2034 (0.0030) | 0.2001 (0.0042) |
| L65-3366 | C | E1E2E3E4e5E7 | 4.23 (0.05) | 4.31 (0.09) | 0.1916 (0.0031) | 0.1910 (0.0050) |
| L71L-3015 | H | E1e2E3E4E5E7 | 4.21 (0.05) | 4.28 (0.09) | 0.1907 (0.0026) | 0.1889 (0.0050) |
| L74-66 | H | e1E2E3E4E5E7 | 3.12 (0.12) | 3.36 (0.16) | 0.1276 (0.0069) | 0.1361 (0.0087) |
| L92-1195 | C | e1E2E3E4E5E7 | 3.21 (0.12) | 2.99 (0.13) | 0.1310 (0.0069) | 0.1114 (0.0080) |

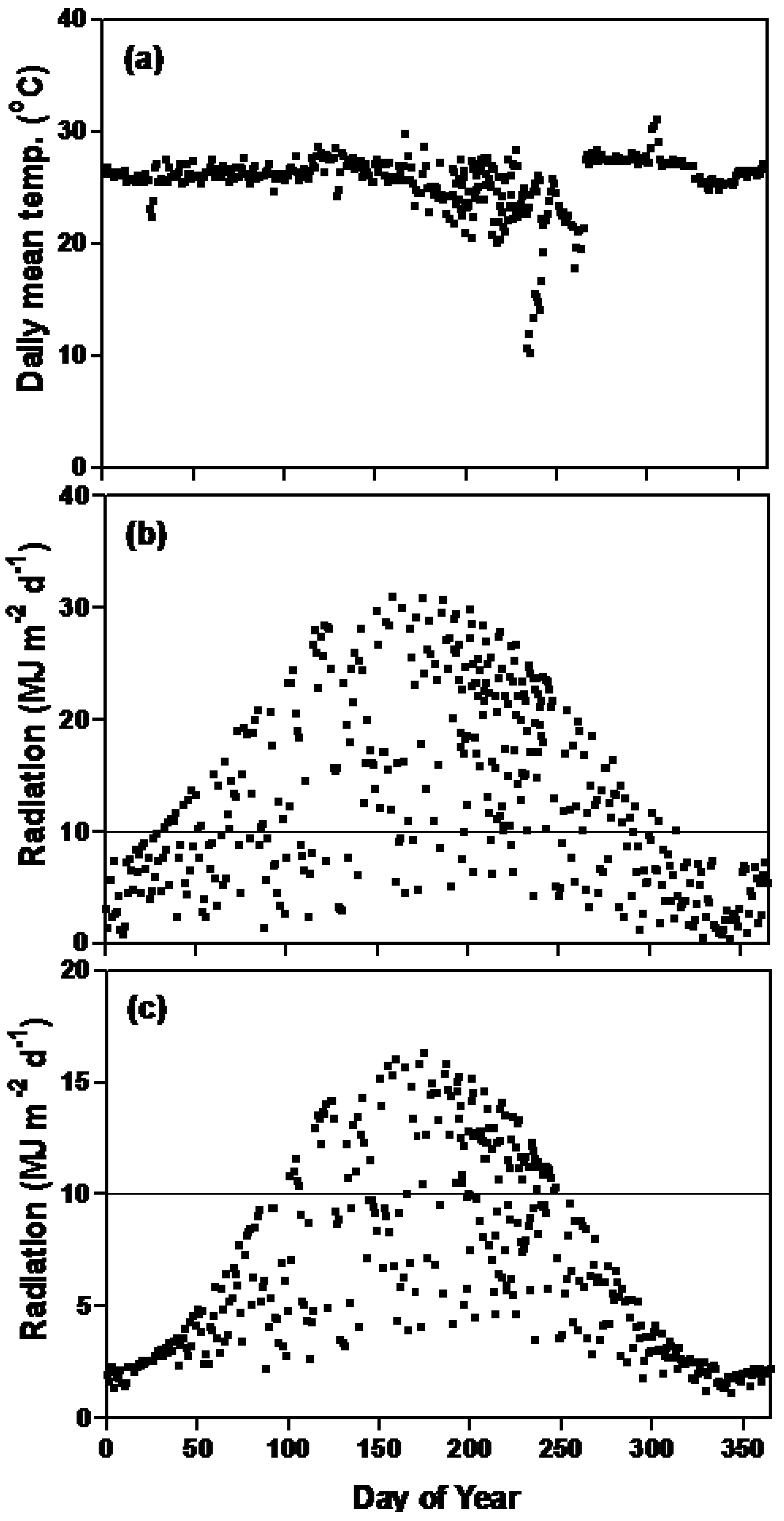
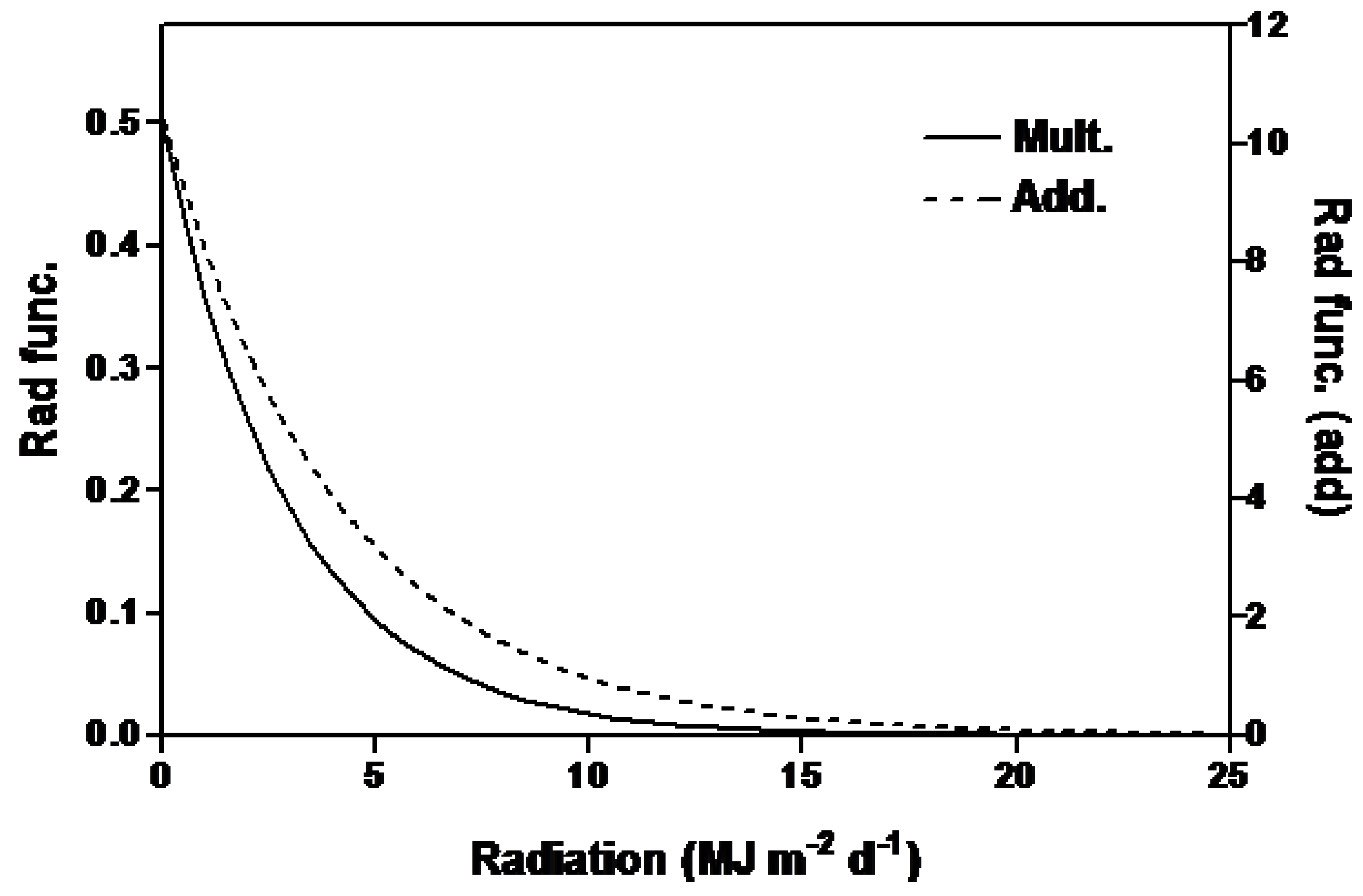
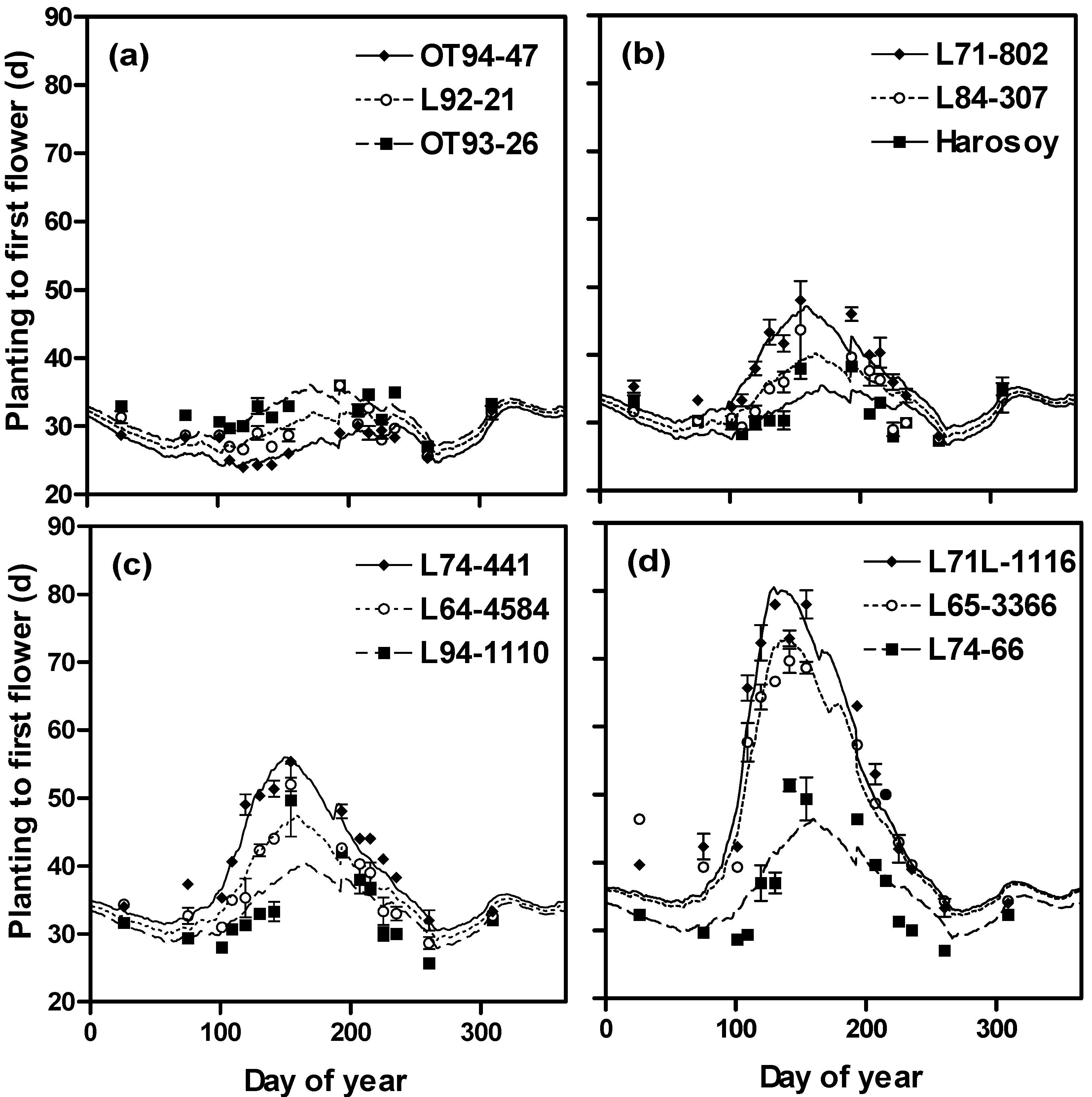
2.1. Irradiance



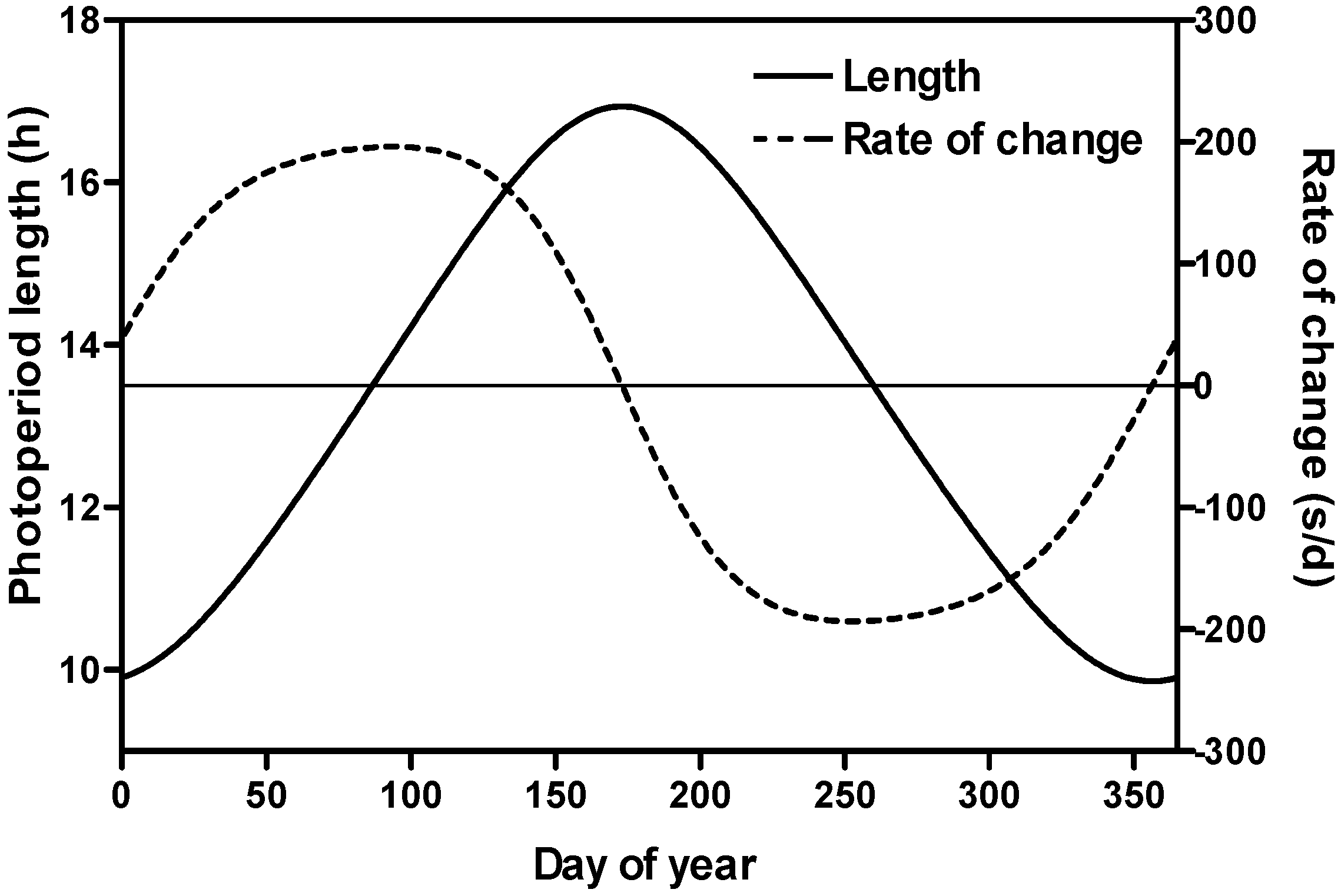
2.2. Rate of Photoperiod Change
| Model | Multiplicative | Additive | |||
|---|---|---|---|---|---|
| N | R2 | SEE (d) | R2 | SEE (d) | |
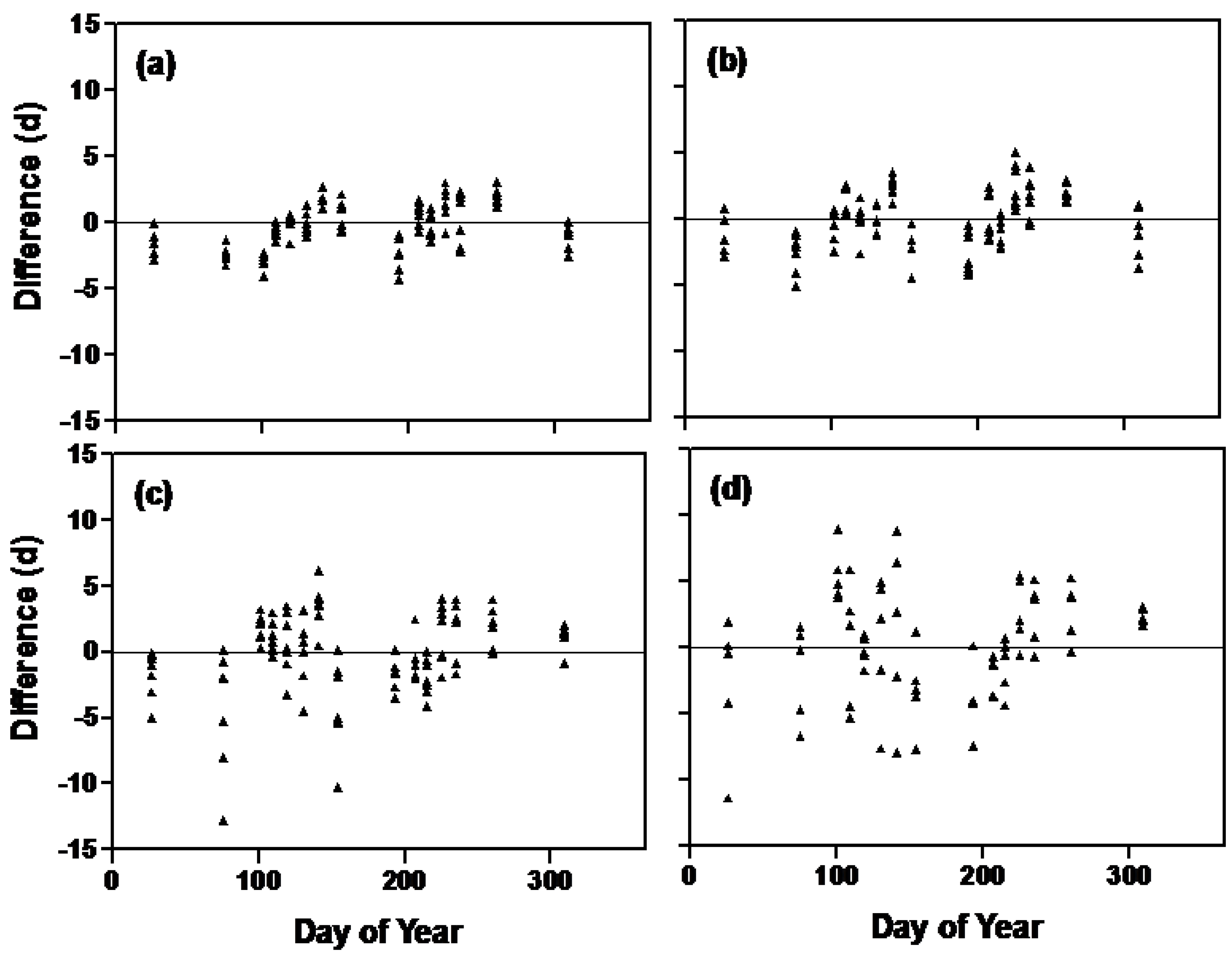
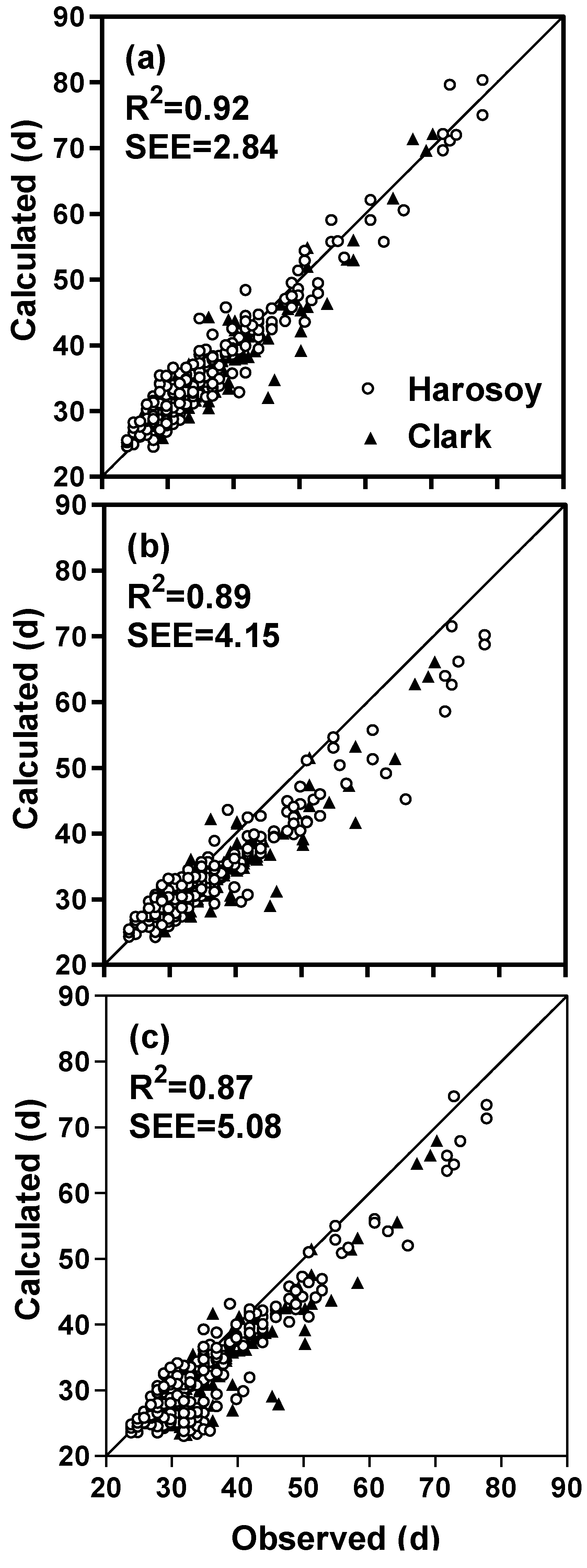
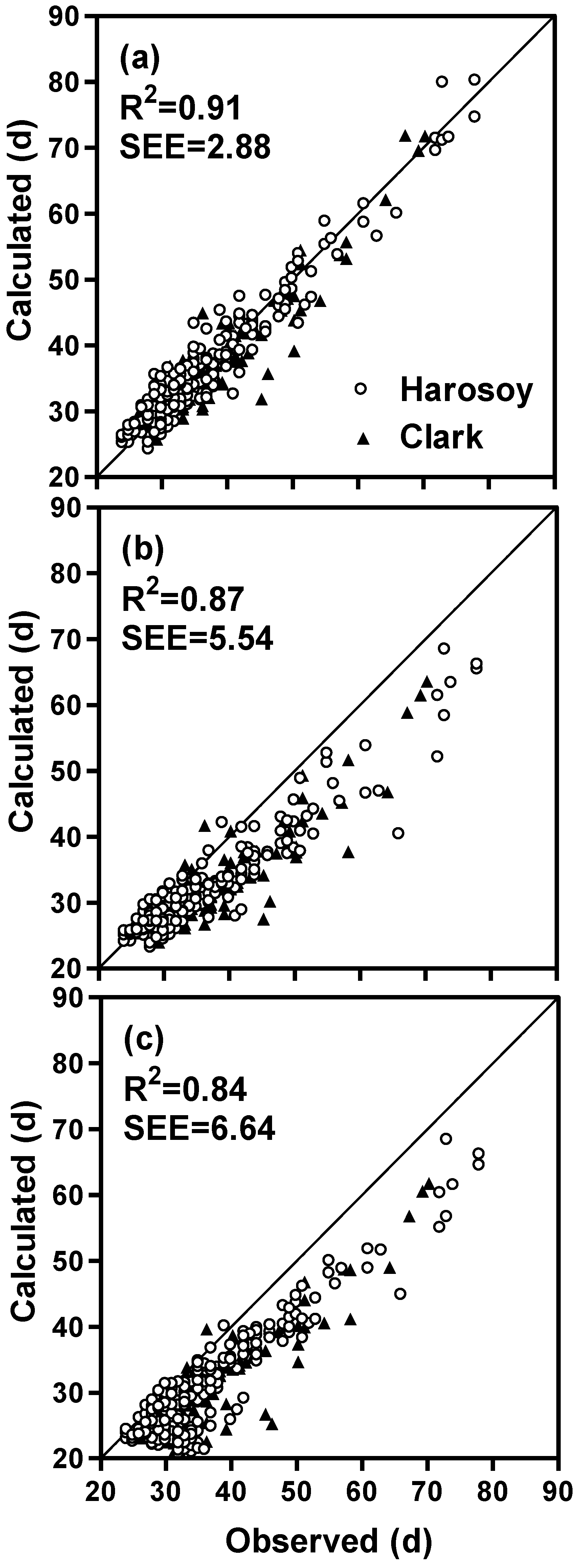
| Complete model † (full dataset) | 389 | 0.92 | 2.84 | 0.91 | 2.88 |
| Without non-linear photoperiod function | 389 | 0.90 | 3.18 | 0.89 | 3.34 |
| Without rate of photoperiod change function | 389 | 0.90 | 4.15 | 0.87 | 5.54 |
| Without irradiance function | 389 | 0.87 | 5.08 | 0.84 | 6.64 |
| Without all of the above three functions | 389 | 0.80 | 7.46 | 0.75 | 10.10 |
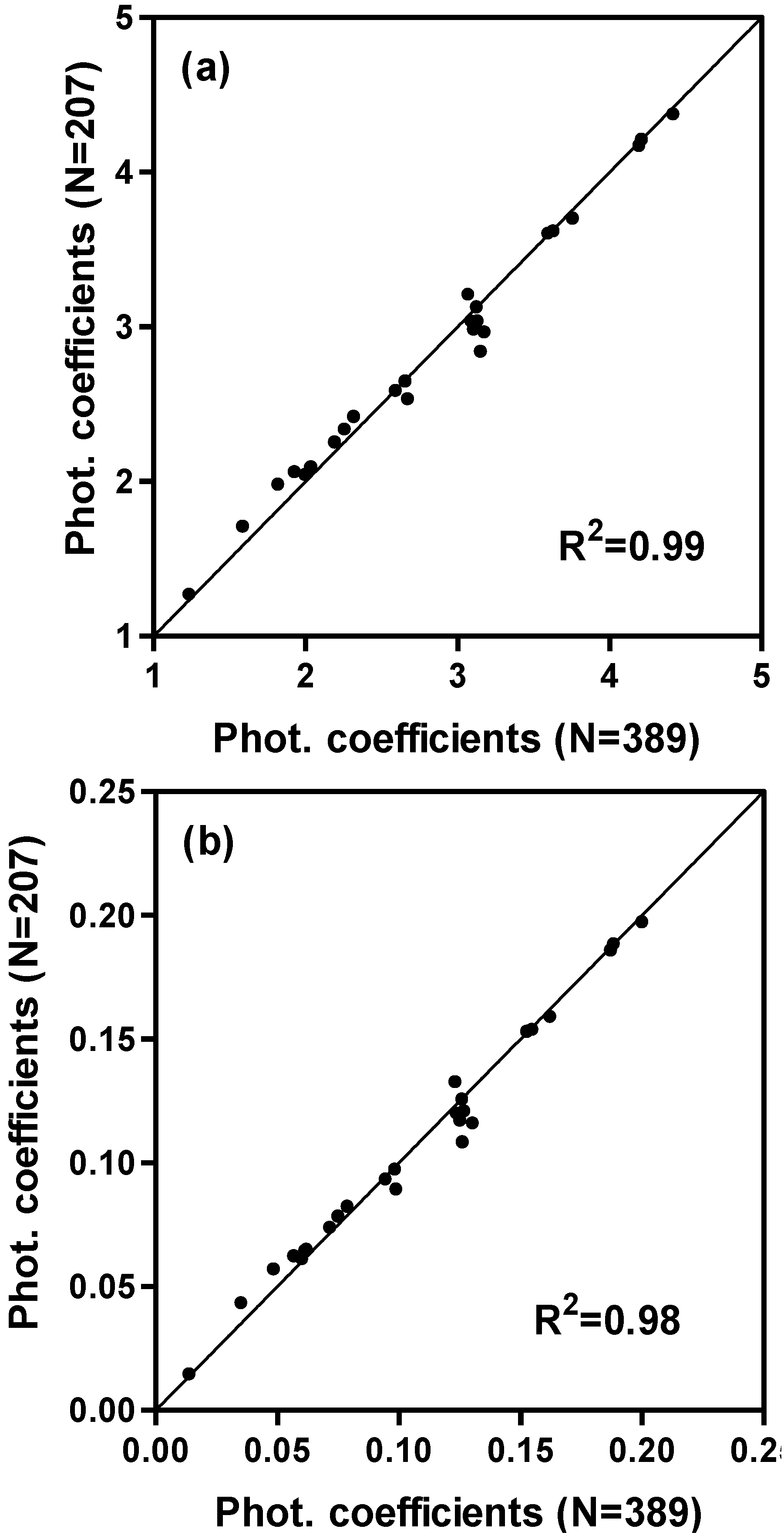
| Complete model (dependent data) | 207 | 0.92 | 2.53 | 0.93 | 2.89 |
| Complete model (independent data) | 187 | 0.89 | 3.61 | 0.90 | 3.55 |
| N | Multiplicative Model | Additive Model | ||||||
|---|---|---|---|---|---|---|---|---|
| Coefficient | Value | SE | Units | Coefficient | Value | SE | Units | |
| 389 | b * | 0.0021 | 3.4 × 10−5 | °C−1 d−1 | b | 0.0024 | 3.8 × 10−5 | °C−1 d−1 |
| 389 | g * (plus) | 9.464 | 1.080 | dh−1 | g (plus) | 11.40 | 0.983 | dh−1 |
| 389 | g * (neg) | −9.822 | 1.078 | dh−1 | g (neg) | −11.02 | 0.921 | dh−1 |
| 389 | h * | 0.499 | 0.060 | h−1 | h | 10.20 | 0.438 | OC h−1 |
| 389 | k * | 0.333 | 0.066 | d J−1 | k | 0.228 | 0.023 | d J−1 |
| 207 | b * | 0.0021 | 4.2 × 10−5 | °C−1 d−1 | b | 0.0025 | 3.7 × 10−5 | °C−1 d−1 |
| 207 | g * (plus) | 7.975 | 1.461 | dh−1 | g (plus) | 12.92 | 0.804 | dh−1 |
| 207 | g * (neg) | −11.62 | 1.353 | dh−1 | g (neg) | −14.64 | 0.795 | dh−1 |
| 207 | h * | 0.460 | 0.050 | h−1 | h | 11.94 | 0.484 | °C h−1 |
| 207 | k * | 0.297 | 0.068 | d J−1 | k | 0.302 | 0.018 | d J−1 |
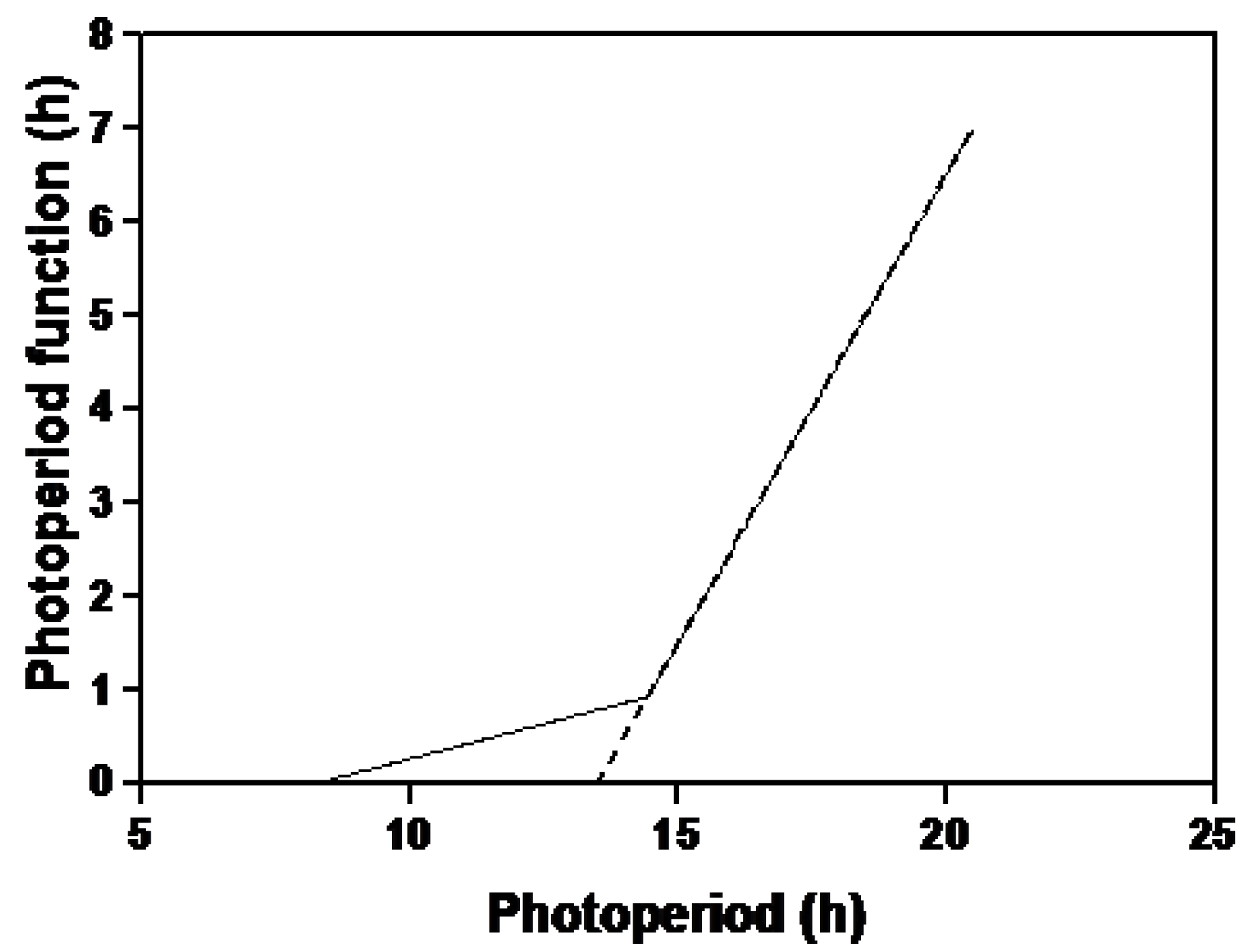
2.3. Photoperiod

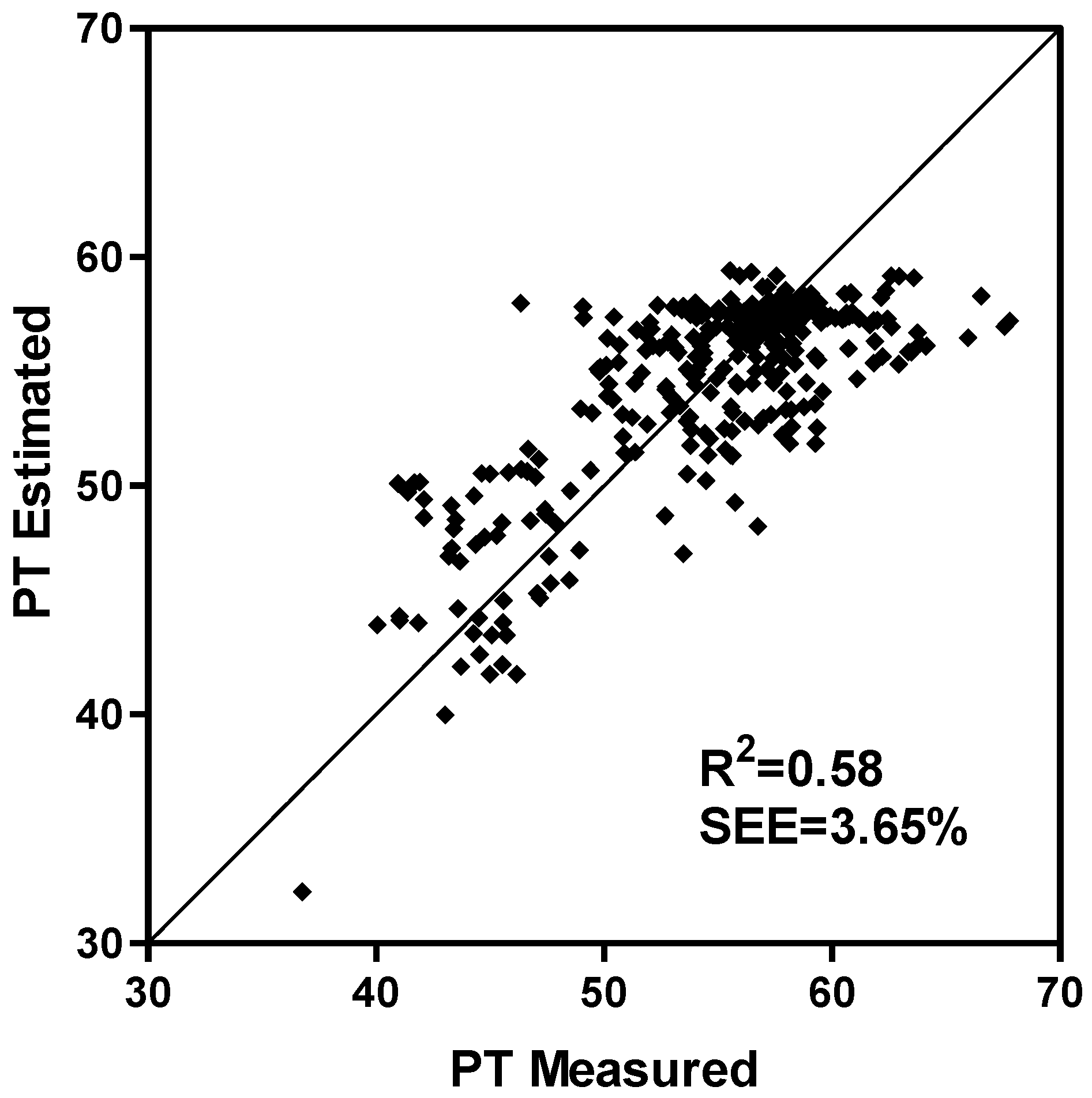
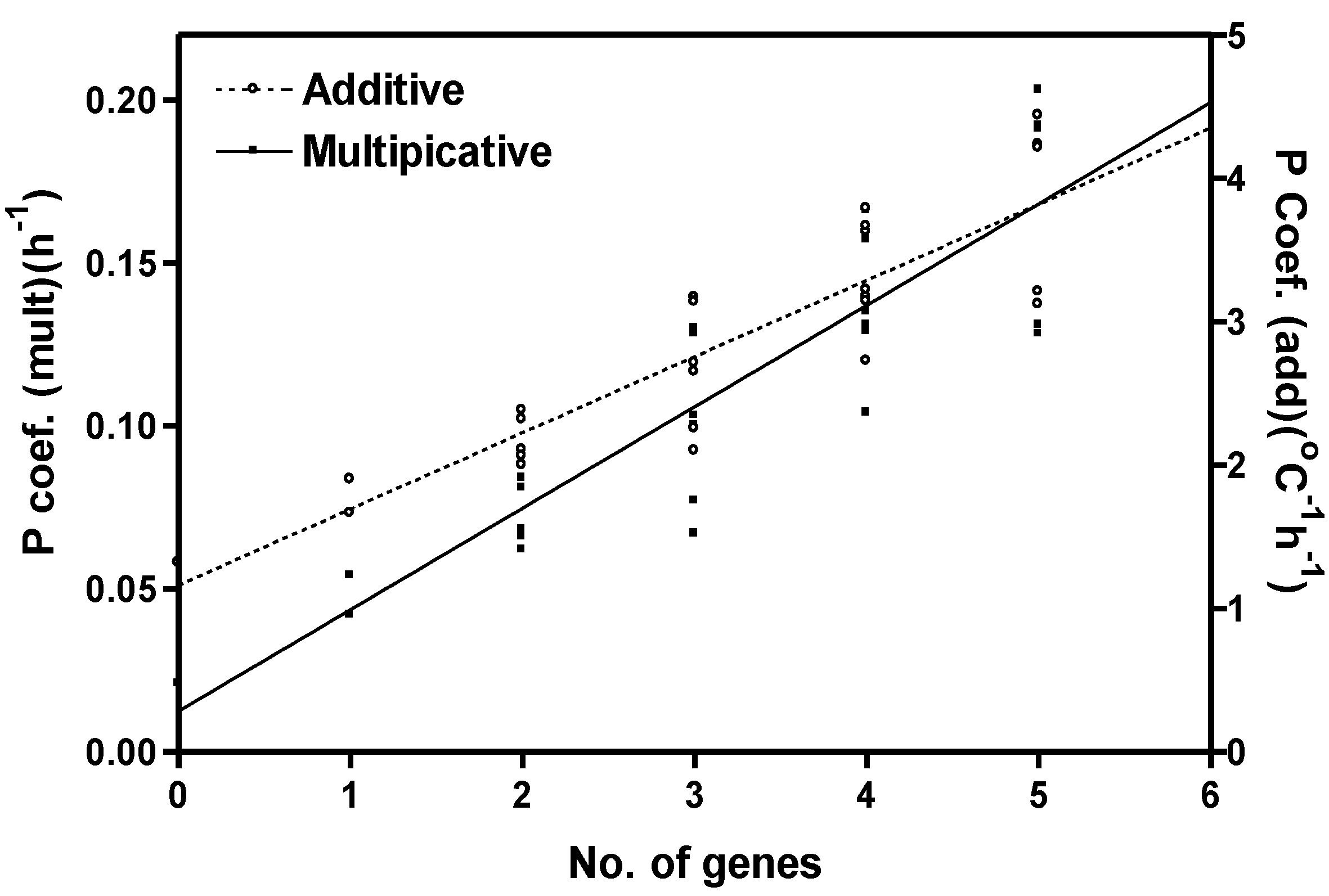
2.4. Model Development and Extension

| Coefficient | Value | SE | Units |
|---|---|---|---|
| a1 | 61.17 | 1.81 | % |
| a2 | −25.47 | 2.20 | % |
| a3 | −0.061 | 0.039 | % °C −1 |
| a4 | 0.439 | 0.050 | % °C −1 |


3. Experimental
3.1. Plant Experiments
3.2. Theoretical Considerations for Modeling
4. Conclusions



Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bernard, R.L. Two major genes for time of flowering and maturity in soybeans. Crop Sci. 1971, 11, 242–244. [Google Scholar] [CrossRef]
- Buzzell, R.I. Inheritance of a soybean flowering response to fluorescent-daylength conditions. Can. J. Genet. Cytol. 1971, 13, 703–707. [Google Scholar]
- Buzzell, R.I.; Voldeng, H.D. Inheritance of insensitivity to long daylength. Soybean Genet. Newsl. 1980, 7, 26–29. [Google Scholar]
- Saindon, G.; Beversdorf, W.D.; Voldeng, H.D. Adjustment of the soybean phenology using the E4 locus. Crop Sci. 1989, 29, 1361–1365. [Google Scholar] [CrossRef]
- McBlain, B.A.; Bernard, R.L. A new gene affecting the time of flowering and maturity in soybeans. J. Hered. 1987, 78, 160–162. [Google Scholar]
- Bonato, E.R.; Vello, N.A. E6, a dominant gene conditioning early flowering and maturity in soybeans. Genet. Mol. Biol. 1999, 22, 229–232. [Google Scholar] [CrossRef]
- Cober, E.R.; Voldeng, H.D. A new soybean maturity and photoperiod-sensitivity locus linked to E1 and T. Crop Sci. 2001, 41, 698–701. [Google Scholar] [CrossRef]
- Cober, E.R.; Molnar, S.J.; Charette, M.; Voldeng, H.D. A new locus for early maturity in soybean. Crop Sci. 2010, 50, 524–527. [Google Scholar] [CrossRef]
- Watanabe, S.; Hideshima, R.; Xia, Z.; Tsubokura, Y.; Sato, S.; Nakamoto, Y.; Yamanaka, N.; Takahashi, R.; Ishimoto, M.; Anai, T.; et al. Map-based cloning of the gene associated with the soybean maturity locus E3. Genetics 2009, 182, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Kanazawa, A.; Matsumura, H.; Takahashi, R.; Harada, K.; Abe, J. Genetic redundancy in soybean photoresponses associated with duplication of the phytochrome A gene. Genetics 2008, 180, 995–1007. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Xia, Z.; Hideshima, R.; Tsubokura, Y.; Sato, S.; Yamanaka, N.; Takahashi, R.; Anai, T.; Tabata, S.; Kitamura, K.; et al. A map-based cloning strategy employing a residual heterozygous line reveals that the GIGANTEA gene is involved in soybean maturity and flowering. Genetics 2011, 188, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Watanabe, S.; Yamada, T.; Tsubokura, Y.; Nakashima, H.; Zhai, H.; Anai, T.; Sato, S.; Yamazaki, T.; Lü, S.; et al. Positional cloning and characterization reveal the molecular basis for soybean maturity locus E1 that regulates photoperiodic flowering. Proc. Natl. Acad. Sci. USA 2012, 109, E2155–E2164. [Google Scholar] [CrossRef] [PubMed]
- Cober, E.R.; Tanner, J.W.; Voldeng, H.D. Genetic control of photoperiod response in early-maturing, near-isogenic soybean lines. Crop Sci. 1996, 36, 601–605. [Google Scholar] [CrossRef]
- Cober, E.R.; Stewart, D.W.; Voldeng, H.D. Photoperiod and temperature responses in early-maturing, near-isogenic soybean lines. Crop Sci. 2001, 41, 721–727. [Google Scholar] [CrossRef]
- Verma, V.D.; Sawaji, B.V. Screening of soybean germplasm for photoperiodic insensitivity under natural conditions in Vidarbha region of Maharashtra, India. Soybean Genet. Newsl. 1994, 21, 137–140. [Google Scholar]
- Bhatia, V.S.; Manglik, P.; Bhatnagar, P.S.; Guruprasad, K.N. Variation in sensitivity of soybean genotypes to varying photoperiods in India. Soybean Genet. Newsl. 1997, 24, 99–100. [Google Scholar]
- Mayers, J.; Lawn, R.; Byth, D. Agronomic studies on soybean (Glycine max (L.) Merrill) in the dry season of the tropics. III. Effect of artificial photoperiod extension on phenology, growth and seed yield. Aust. J. Agric. Res. 1991, 42, 1109–1121. [Google Scholar] [CrossRef]
- Hadley, P.; Roberts, E.H.; Summerfield, R.J.; Minchin, F.R. Effects of temperature and photoperiod on flowering in soya bean [Glycine max (L.) Merrill]: A quantitative model. Ann. Bot. 1984, 53, 669–681. [Google Scholar]
- Constable, G.A.; Rose, I.A. Variability of soybean phenology response to temperature, daylength and rate of change in daylength. Field Crops Res. 1988, 18, 57–69. [Google Scholar] [CrossRef]
- Summerfield, R.J.; Lawn, R.J.; QI, A.; Ellis, R.H.; Roberts, E.H.; Chay, P.M.; Brouwer, J.B.; Rose, J.L.; Shanmugasundaram, S.; Yeates, S.J.; et al. Towards the reliable prediction of time to flowering in six annual crops. II. Soyabean ( Glycine max). Exp. Agric. 1993, 29, 253–289. [Google Scholar] [CrossRef]
- Summerfield, R.J.; Roberts, E.H.; Ellis, R.H.; Lawn, R.J. Towards the reliable prediction of time to flowering in six annual crops. I. The development of simple models for fluctuating field environments. Exp. Agric. 1991, 27, 11–31. [Google Scholar] [CrossRef]
- Upadhyay, A.P.; Ellis, R.H.; Summerfield, R.J.; Roberts, E.H.; Qi, A. Characterization of photothermal flowering responses in maturity isolines of soyabean [Glycine max (L.) Merrill] cv. Clark. Ann. Bot. 1994, 74, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Wallace, D.H. Simulation and prediction of plant phenology for five crops based on photoperiod × temperature interaction. Ann. Bot. 1998, 81, 705–716. [Google Scholar] [CrossRef]
- Major, D.J.; Johnson, D.R.; Tanner, J.W.; Anderson, I.C. Effects of daylength and temperature on soybean development1. Crop Sci. 1975, 15, 174–179. [Google Scholar] [CrossRef]
- Grimm, S.S.; Jones, J.W.; Boote, K.J.; Hesketh, J.D. Parameter estimation for predicting flowering date of soybean cultivars. Crop Sci. 1993, 33, 137–144. [Google Scholar] [CrossRef]
- Grimm, S.S.; Jones, J.W.; Boote, K.J.; Herzog, D.C. Modeling the occurrence of reproductive stages after flowering for four soybean cultivars. Agron. J. 1994, 86, 31–38. [Google Scholar] [CrossRef]
- Piper, E.L.; Boote, K.J.; Jones, J.W.; Grimm, S.S. Comparison of two phenology models for predicting flowering and maturity date of soybean. Crop Sci. 1996, 36, 1606–1614. [Google Scholar] [CrossRef]
- Piper, E.L.; Smit, M.A.; Boote, K.J.; Jones, J.W. The role of daily minimum temperature in modulating the development rate to flowering in soybean. Field Crops Res. 1996, 47, 211–220. [Google Scholar] [CrossRef]
- Stewart, D.W.; Cober, E.R.; Bernard, R.L. Modeling genetic effects on the photothermal response of soybean phenological development. Agron. J. 2003, 95, 65–70. [Google Scholar] [CrossRef]
- Stewart, D.W.; AAFC-ECORC, Ottawa, ON, Canada. Unpublished data. 2006.
- Birch, C.J.; Vos, J.; Kiniry, J.; Bos, H.J.; Elings, A. Phyllochron responds to acclimation to temperature and irradiance in maize. Field Crops Res. 1998, 59, 187–200. [Google Scholar] [CrossRef]
- Tollenaar, M. Duration of the grain-filling period in maize is not affected by photoperiod and incident PPFD during the vegetative phase. Field Crops Res. 1999, 62, 15–21. [Google Scholar] [CrossRef]
- Bertero, H.D. Effects of photoperiod, temperature and radiation on the rate of leaf appearance in quinoa (Chenopodium quinoa Willd.) under field conditions. Ann. Bot. 2001, 87, 495–502. [Google Scholar] [CrossRef]
- Zhang, D.; Armitage, A.M.; Affolter, J.M.; Dirr, M.A. Environmental control of flowering and growth of Achillea millefolium L. “Summer Pastels”. HortScience 1996, 31, 364–365. [Google Scholar]
- Mattson, N.S.; Erwin, J.E. The impact of photoperiod and irradiance on flowering of several herbaceous ornamentals. Sci. Hortic. 2005, 104, 275–292. [Google Scholar] [CrossRef]
- Kirby, E.J.M.; Appleyard, M.; Fellowes, G. Effect of sowing date on the temperature response of leaf emergence and leaf size in barley. Plant Cell Environ. 1982, 5, 477–484. [Google Scholar] [CrossRef]
- Kernich, G.C.; Slafer, G.A.; Halloran, G.M. Barley development as affected by rate of change of photoperiod. J. Agric. Sci. 1995, 124, 379–388. [Google Scholar] [CrossRef]
- Baker, C.K.; Gallagher, J.N.; Monteith, J.L. Daylength change and leaf appearance in winter wheat. Plant Cell Environ. 1980, 3, 285–287. [Google Scholar]
- Slafer, G.A.; Connor, D.J.; Halloran, G.M. Rate of leaf appearance and final number of leaves in wheat: Effects of duration and rate of change of photoperiod. Ann. Bot. 1994, 74, 427–436. [Google Scholar] [CrossRef]
- Slafer, G.A.; Halloran, G.M.; Connor, D.J. Development rate in wheat as affected by duration and rate of change of photoperiod. Ann. Bot. 1994, 73, 671–677. [Google Scholar] [CrossRef]
- Bonhomme, R.; Derieux, M.; Kiniry, J.R.; Edmeades, G.O.; Ozier-Lafontaine, H. Maize leaf number sensitivity in relation to photoperiod in multilocation field trials. Agron. J. 1991, 83, 153–157. [Google Scholar] [CrossRef]
- Clerget, B.; Dingkuhn, M.; Chantereau, J.; Hemberger, J.; Louarn, G.; Vaksmann, M. Does panicle initiation in tropical sorghum depend on day-to-day change in photoperiod? Field Crops Res. 2004, 88, 21–37. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Neumaier, N.; Farias, J.R.B.; Nepomuceno, A.L. Comparison of vegetative development in soybean cultivars for low-latitude environments. Field Crops Res. 2005, 92, 53–59. [Google Scholar] [CrossRef]
- List, R.J. Smithsonian meteorological tables. Smithson. Misc. Collect. 1949, 114, 1–527. [Google Scholar]
- Marquardt, D.W. An algorithm for least-squares estimation of nonlinear parameters. J. Soc. Ind. Appl. Math. 1963, 11, 431–441. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cober, E.R.; Curtis, D.F.; Stewart, D.W.; Morrison, M.J. Quantifying the Effects of Photoperiod, Temperature and Daily Irradiance on Flowering Time of Soybean Isolines. Plants 2014, 3, 476-497. https://doi.org/10.3390/plants3040476
Cober ER, Curtis DF, Stewart DW, Morrison MJ. Quantifying the Effects of Photoperiod, Temperature and Daily Irradiance on Flowering Time of Soybean Isolines. Plants. 2014; 3(4):476-497. https://doi.org/10.3390/plants3040476
Chicago/Turabian StyleCober, Elroy R., Daniel F. Curtis, Douglas W. Stewart, and Malcolm J. Morrison. 2014. "Quantifying the Effects of Photoperiod, Temperature and Daily Irradiance on Flowering Time of Soybean Isolines" Plants 3, no. 4: 476-497. https://doi.org/10.3390/plants3040476
APA StyleCober, E. R., Curtis, D. F., Stewart, D. W., & Morrison, M. J. (2014). Quantifying the Effects of Photoperiod, Temperature and Daily Irradiance on Flowering Time of Soybean Isolines. Plants, 3(4), 476-497. https://doi.org/10.3390/plants3040476