
Phytate (Inositol Hexakisphosphate) in Soil and Phosphate Acquisition from Inositol Phosphates by Higher Plants. A Review

{kind=link}
Abstract
:1. Introduction
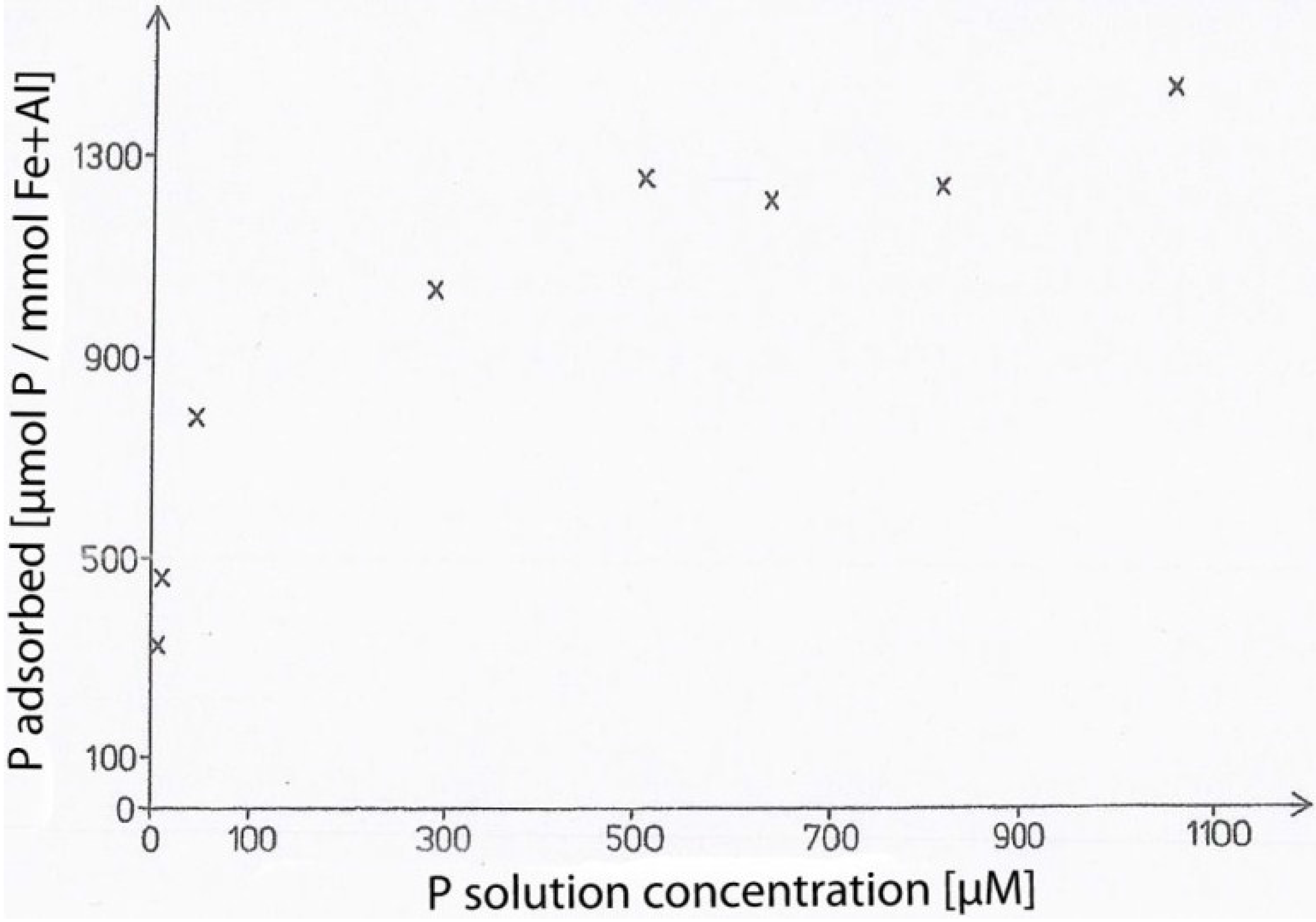
2. The Binding of Inositol Hexakisphosphate in Soil

3. The Role of Phosphatases (Phytases) in IHP-P Acquisition from Soil by Plants
4. Mobilization of IHP from the Soil Solid Phase: Mechanisms and Relevance
5. The Role of Soil Microorganisms in the P Acquisition from IHP-P
- the role of mycorrhiza;
- the effect of microorganisms on the mineralization on root released carboxylates; and/or
- the role of soil microorganisms in producing phosphatases (phytases).
6. Summary and Outlook
- Research on the binding of IHP in soil with a main focus on the binding to the soil humic substances.
- Mobilization of IHP and other inositol phosphates from the soil solid phase by root exudates with the central focus on the role of di- and tricarboxylic acids and the acquisition of IHP-P by the roots of higher plants.
Conflicts of Interest
References
- Cosgrove, D.J. Inositol Phosphates: Their Chemistry, Biochemistry and Physiology; Elsevier: Amsterdam, The Netherlands, 1980. [Google Scholar]
- Annunziata, M.F. Origin and biochemical transformations of inositol stereoisomers and their phosphorylated derivatives in soil. In Inositol Phosphates: Linking Agriculture and the Environment; Turner, B.L., Richardson, A.E., Mullaney, E.J., Eds.; CABI: Wallingford, UK, 2007; pp. 41–60. [Google Scholar]
- Turner, B.L.; Chessman, A.W.; Godage, H.Y.; Riley, A.M.; Potter, B.V. Determination of neo- and D-chiro-inositol hexakisphosphate in soils by solution 31P NMR spectroscopy. Environ. Sci. Technol. 2012, 46, 4994–5002. [Google Scholar] [CrossRef] [PubMed]
- Mullaney, E.J.; Ullah, A.H. Phytases: Attributes, catalytic mechanisms and applications. In Inositol Phosphates: Linking Agriculture and the Environment; Turner, B.L., Richardson, A.E., Mullaney, E.J., Eds.; CABI: Wallingford, UK, 2007; pp. 97–110. [Google Scholar]
- Dao, T.H. Oranic ligand effects on the enzymatic dephosphorylation of myo-inositol hexakis dihydrogenphosphate in dairy wastewater. J. Environ. Qual. 2004, 33, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Toor, G.S.; Peak, J.D.; Sims, J.T. Phosphorus speciation in broiler litter and turkey manure produced from modified diets. J. Environ. Qual. 2005, 34, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.H. Ligand effects on inositol phosphate solubility and bioavailability in animal manures. In Inositol Phosphates: Linking Agriculture and the Environment; Turner, B.L., Richardson, A.E., Mullaney, E.J., Eds.; CABI: Wallingford, UK; Cambridge, MA, USA, 2007; pp. 169–185. [Google Scholar]
- Lott, J.N.A.; Ockenden, I.; Raboy, V.; Batten, G.D. Phytic acid and phosphorus in crop seeds and fruits: A global estimate. Seed Sci. Res. 2000, 10, 11–33. [Google Scholar]
- McKercher, R.; Anderson, G. Organic phosphate sorption by neutral and basic soils. Commun. Soil Sci. Plant Anal. 1989, 20, 723–732. [Google Scholar] [CrossRef]
- Celi, L.; Barberis, E. Abiotic stabilization of organic phosphorus in the environment. In Organic Phosphorus in the Environment; Turner, B.L., Frossard, E., Baldwin, D.S., Eds.; CABI: Oxfordshire, UK, 2005; pp. 113–132. [Google Scholar]
- Celi, L.; Barberis, E. Abiotic reaction of inositol phosphates in soil. In Inositol Phosphates: Linking Agriculture and the Environment; Turner, B.L., Richardson, A.E., Mullaney, E.J., Eds.; CABI: Wallingford, UK, 2007; pp. 207–220. [Google Scholar]
- Dalal, R.C. Soil organic phosphorus. Adv. Agron. 1977, 29, 85–117. [Google Scholar]
- Anderson, G. Other organic compounds. In Soil Components. Vol 1: Organic Components; Gieseking, J.E., Ed.; Springer: Berlin, Heidelberg, Germany, 1975; pp. 305–331. [Google Scholar]
- Anderson, G. Assessing organic phosphorus in soils. In The Role of Phosphorus in Agriculture; Khasawneh, F.E., Sample, E.C., Kamprath, E.J., Eds.; American Society of Agronomy: Madison, WI, USA, 1980; pp. 411–431. [Google Scholar]
- Tate, K. Soil phosphorus. In Soil Organic Matter and Biological Activity; Vaughan, D., Malcolm, R., Eds.; MartiniusNijhoff/Dr.W Jung Publishers: Dorderecht, The Netherlands, 1985; pp. 329–377. [Google Scholar]
- Harrison, A.F. Soil Organic Phosphorus: A Review of World Literature; CABI: Wallingford, UK, 1987. [Google Scholar]
- Stewart, J.W.; Tiessen, H. Dynamics of soil organic phosphorus. Biogeochemistry 1987, 4, 41–60. [Google Scholar] [CrossRef]
- Turner, B.L. Inositol phosphates in soil: Amounts, forms and significance of the phosphorylated inositol stereoisomers. In Inositol Phosphates: Linking Agriculture and the Environment; Turner, B.L., Richardson, A.E., Mullaney, E.J., Eds.; CABI: Wallingford, UK, 2007; pp. 186–206. [Google Scholar]
- Gerke, J. Humic (Organic matter)-Al(Fe)-phsophate complexes: An underestimated phosphate form in soils and source of plant-available phosphate. Soil Sci. 2010, 175, 417–425. [Google Scholar] [CrossRef]
- Thomas, R.L.; Bowman, B.T. The occurrence of high molecular weight phosphorus compounds in soil. Soil Sci. Soc. Am. Proc. 1966, 30, 799–801. [Google Scholar] [CrossRef]
- Moyer, J.; Thomas, R.L. Organic phosphorus and inositol phosphates in molecular size fractions of a soil organic matter extract. Soil Sci. Soc. Am. Proc. 1970, 34, 80–84. [Google Scholar] [CrossRef]
- Stewart, J.H.; Tate, M.E. Gel chromatography of soil organic phosphorus. J. Chromatogr. 1971, 60, 75–82. [Google Scholar] [CrossRef]
- Veinot, R.; Thomas, R.L. High molecular weight organic phosphorus complexes in soil organic matter: Inositol and metal content of various fractions. Soil Sci. Soc. Am. Proc. 1972, 36, 71–73. [Google Scholar] [CrossRef]
- Hong, J.K.; Yamane, I. Inositol phosphate and inositol in humic acid and fulvic acid fractions extracted by three methods. Soil Sci. Plant Nutr. 1980, 26, 491–496. [Google Scholar] [CrossRef]
- Hong, J.K.; Yamane, I. Distribution of inositol phosphate in the molecular size fractions of humic and fulvic acid fractions. Soil Sci. Plant Nutr. 1981, 27, 295–303. [Google Scholar] [CrossRef]
- Borie, F.; Zunino, H.; Martinez, L. Macromolecular associations and inositol phosphates in some Chilean volcanic soils of temperate regions. Commun. Soil Sci. Plant Anal. 1989, 20, 1881–1894. [Google Scholar] [CrossRef]
- Gerke, J. Chemische Prozesse der Nährstoffmobilisierung in der Rhizospäre und ihre Bedeutung für den Übergang vom Boden in die Pflanze; Cuvillier Verlag: Göttingen, Germany, 1995. [Google Scholar]
- Negrin, M.A.; Gonzalez-Carcedo, S.; Hernandez-Moreno, J.M. P fractionation in sodium bicarbonate extracts of andic soils. Soil Biol. Biochem. 1997, 27, 761–766. [Google Scholar] [CrossRef]
- Kirkby, E.A.; Johnston, A.E. Soil and fertilizer phosphorus in relation to crop nutrition. In The Ecophysiology of Plant-Phosphorus Interactions; Hammond, J.P., White, P.J., Eds.; Springer: New York, NY, USA, 2008; pp. 177–223. [Google Scholar]
- Römer, W. Ansätze für eine effizientere Nutzung des Phosphors auf der Basis experimenteller Befunde. Ber. Landwirtsch. 2009, 87, 5–30. [Google Scholar]
- Schwertmann, U. Differenzierung der Eisenoxide des Bodens durch Extraktion mit Ammoniumoxalatlösung. Z. Pflanzenernähr. Bodenk. 1964, 105, 169–174. [Google Scholar]
- Bowden, J.W.; Nagarajah, S.; Barrow, N.J.; Posner, A.M.; Quirk, J.P. Describing the adsorption of phosphate, citrate and selenite on a variable charge mineral surface. Aust. J. Soil Res. 1980, 18, 49–60. [Google Scholar] [CrossRef]
- Davies, J.A.; Leckie, J.O. Surface ionization and complexation at the oxide/water interface. 3. Adsorption of anions. J. Colloid Interface Sci. 1980, 74, 32–43. [Google Scholar] [CrossRef]
- Geelhoed, J.S.; van Riemsdijk, W.H.; Findenegg, G.R. Phosphate and sulfate adsorption on goethite. Single adsorption and competitive adsorption. Geochim. Cosmochim. Acta 1997, 61, 2389–2396. [Google Scholar] [CrossRef]
- Yan, Y; Li, W.; Zheng, A.; Liu, F.; Feng, X.; Sparks, D.L. Mechanisms of myo-inositol hexakisphsophate sorption on amorphous aluminum hydroxide: Spectroscopic evidences for rapid surface precipitation. Environ. Sci. Technol. 2014, 48, 6735–6742. [Google Scholar] [CrossRef] [PubMed]
- Cade-Menun, U. Using phosphorus-31 nuclear magnetic resonance spectroscopy to characterize organic phosphorus in environmental samples. In Organic Phosphorus in the Environment; Turner, B.L., Frossard, E., Baldwin, D.S., Eds.; CABI: Wallingford, UK, 2005; pp. 21–44. [Google Scholar]
- Richardson, A.E.; George, T.S.; Hens, M.; Simpson, R.J. Utilization of soil organic phosphorus by higher plants. In Organic Phosphorus in the Environment; Turner, B.L., Frossard, E., Baldwin, D.S., Eds.; CABI: Wallingford, UK, 2005; pp. 161–184. [Google Scholar]
- Turner, B.L.; Cade-Menun, B.J.; Westermann, D.T. Organic phosphorus composition and potential bioavailability in semi-arid arable soils of the western United States. Soil Sci. Soc. Am. J. 2003, 67, 1168–1179. [Google Scholar] [CrossRef]
- Turner, B.L. Soil organic phosphorus in tropical forests: An assessment of the NAOH-EDTA extraction procedure for quantitative analysis by solution 31P NMR spectroscopy. Eur. J. Soil Sci. 2008, 59, 453–466. [Google Scholar] [CrossRef]
- Urrutia, O.; Erro, J.; Guardada, I.; San Francisco, S.; Mandada, M.; Baigorri, R.; Yvin, J.C.; Garcia-Mina, J.M. Physico-chemical characterization of humic-metal-phosphate complexes and their potential application to the manufacture of new types of phosphate-based fertilizers. J. Plant Nutr. Soil Sci. 2014, 177, 128–136. [Google Scholar] [CrossRef]
- Gerke, J.; Herrmann, R. Adsorption of orthophosphate to humic-Fe complexes and to amorphous Fe-oxide. J. Plant Nutr. Soil Sci. 1992, 155, 233–236. [Google Scholar]
- Tinker, P.B.; Nye, P.H. Solute Movement in the Rhizosphere; Oxford University Press: Oxford, UK; New York, NY, USA, 2000. [Google Scholar]
- Jungk, A. Dynamics of nutrient movement at the soil-root interface. In Plant Roots, the Hidden Half; Waisel, Y, Eshel, A., Kafkafi, U., Eds.; Dekker: New York, NY, USA, 2002; pp. 587–616. [Google Scholar]
- Schachtmann, D.; Reid, R.J.; Ayling, S.M. Phosphorus uptake by plants: From soil to cell. Plant Physiol. 1998, 116, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Raghotama, K.G.; Kartikeyan, A.S. Phosphate acquisition. Plant Soil 2005, 274, 37–49. [Google Scholar] [CrossRef]
- Tarafdar, J.C.; Claassen, N. Organic phosphorus compounds as a phosphorus source for higher plants through the activity of phosphatases produced by plant roots and microorganisms. Biol. Fertil. Soils 1988, 5, 308–312. [Google Scholar] [CrossRef]
- Hayes, J.E.; Simpson, R.J.; Richardson, A.E. The growth and phosphorus utilization of plants in sterile media when supplied with inositol hexaphosphate, glucose-1-phosphate, or inorganic phosphate. Plant Soil 2000, 220, 165–174. [Google Scholar] [CrossRef]
- George, T.S.; Richardson, A.E.; Hadobas, P.A.; Simpson, R.J. Characterization of transgenic Trifolium subterraneum L. which expresses phyA and releases extracellular phytase: Growth and phosphorus nutrition in laboratory media and soil. Plant Cell Environ. 2004, 27, 1351–1361. [Google Scholar] [CrossRef]
- Findenegg, G.R.; Nelemans, J.A. The effect of phytase on the availability of phosphorus from myo-inositol hexaphosphate (phytate) in maize roots. Plant Soil 1993, 154, 189–196. [Google Scholar] [CrossRef]
- Lung, S.C.; Lim, B.L. Assimilation of phytate-phosphorus by extracellular phytase activity of tobacco (Nicotiana tobaccum) is affected by the availability of soluble phytate. Plant Soil 2006, 279, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Beißner, L. Mobilisierung von Phosphor aus Organischen und Anorganischen Verbindungen Durch Zuckerrübenwurzeln. Ph.D. Thesis, Georg-August Universität, Göttingen, Germany, 1997. [Google Scholar]
- Espinoza, M.; Turner, B.L.; Haygart, P.M. Preconcentration and separation of trace phosphorus compounds in soil leachates. J. Environ. Qual. 1999, 28, 1497–1504. [Google Scholar] [CrossRef]
- Adams, M.; Pate, J.S. Availability of organic and inorganic forms of phosphorus to lupins (Lupinus spp.). Plant Soil 1992, 145, 107–113. [Google Scholar] [CrossRef]
- Gerke, J.; Beißner, L.; Römer, W. The quantitative effect of chemical phosphate mobilization by carboxyxlate anions on P uptake by a single root. I. The basic concept and determination of soil parameters. J. Plant Nutr. Soil Sci. 2000, 163, 207–212. [Google Scholar] [CrossRef]
- Nye, P.H. The diffusion of two interacting solutes in soil. J. Soil Sci. 1983, 34, 677–691. [Google Scholar] [CrossRef]
- Nye, P.H. On estimating the uptake of nutrients solubilized near roots or other surfaces. J. Soil Sci. 1984, 35, 439–446. [Google Scholar] [CrossRef]
- Gerke, J.; Römer, W.; Beißner, L. The quantitative effect of chemical phosphate mobilization by carboxylate anions on P uptake by a single root. II. The importance of soil and plant parameters for uptake of mobilized P. J. Plant Nutr. Soil Sci. 2000, 163, 213–219. [Google Scholar]
- Dinkelaker, B.; Römheld, V.; Marschner, H. Citric acid excretion and precipitation of calcium citrate in the rhizosphere of white lupin (Lupinus albus L.). Plant Cell Environ. 1989, 12, 185–192. [Google Scholar]
- Gerke, J.; Römer, W.; Jungk, A. The excretion of citric and malic acid by proteoid roots of Lupinus albus L.: Effects on solution concentration of phosphate, iron and aluminum in the proteoid rhizosphere in samples of an Oxisol and a Luvisol. J. Plant Nutr. Soil Sci. 1994, 157, 289–304. [Google Scholar]
- Römer, W.; Kang, D-K.; Egle, K.; Gerke, J.; Keller, H. The acquisition of cadmium by Lupinus albus L., Lupinus angustifolius L. and Lolium multiflorum Lam. J. Plant Nutr. Soil Sci. 2000, 163, 623–628. [Google Scholar] [CrossRef]
- Hocking, P.; Jeffery, S. Cluster root production and organic acid exudation in a group of old world lupins and a new world lupin. Plant Soil 2004, 258, 135–150. [Google Scholar] [CrossRef]
- Richardson, A.E.; George, T.S.; Jakobson, I.; Simpson, J.S. Plant utilization of inositol phosphates. In Inositol Phosphates: Linking Agriculture and the Environment; Turner, B.L., Richardson, A.E., Mullaney, E.J., Eds.; CABI: Wallingford, UK, 2007; pp. 242–260. [Google Scholar]
- Giaveno, C.; Celi, L.; Richardson, A.E.; Simpson, R.J.; Barberis, E. Interaction of phytases with minerals and availability of substrate affect the hydrolysis of inositol phosphates. Soil Biol. Biochem. 2010, 42, 491–498. [Google Scholar] [CrossRef]
- Giles, C.D.; Hsu, P.-C.; Richardson, A.E.; Hurst, M.R.; Hill, J.E. Plant assimilation of phosphorus from an insoluble organic form is improved by the addition of organic anion producing Pseudomonas sp. Soil Biol. Biochem. 2014, 68, 263–269. [Google Scholar] [CrossRef]
- Bolan, N.S. A critical review on the role of mycorrhizal fungi in the uptake of phosphorus by plants. Plant Soil 1991, 134, 189–207. [Google Scholar] [CrossRef]
- Yao, Q.; Li, X.I.; Feng, G.; Christie, P. Mobilization of sparingly soluble inorganic phosphates by the external mycelium of an arbuscular mycorrhizal fungus. Plant Soil 2001, 230, 279–285. [Google Scholar] [CrossRef]
- Jungk, A. Root hairs and the acquisition of plant nutrients from soil. J. Plant Nutr. Soil Sci. 2001, 164, 121–129. [Google Scholar] [CrossRef]
- Gerke, J. The acquisition of phosphate by higher plants: Effect of carboxylate release by the roots. A critical review. J. Plant Nutr. Soil Sci. 2015. [Google Scholar] [CrossRef]
- Boudot, J.P. Relative efficiency of complexed aluminum, noncrystalline Al hydroxide, allophone and imogolite in retarding the biodegradation of citric acid. Geoderma 1992, 52, 29–39. [Google Scholar] [CrossRef]
- Jones, D.L.; Edwards, A.C. Influence of sorption on the biological utilization of two simple carbon substrates. Soil Biol. Biochem. 1998, 30, 1895–1902. [Google Scholar] [CrossRef]
- Bowen, G.D.; Rovira, A.D. The rizosphere and its management to improve plant growth. Adv. Agron. 1999, 66, 1–102. [Google Scholar]
- Oberson, A.; Joner, E.J. Microbial turnover of phosphorus in soil. In Organic Phosphorus in the Environment; Turner, B.L., Frossard, E., Baldwin, D.S., Eds.; CABI: Wallingford, UK, 2005; pp. 133–164. [Google Scholar]
- Quiquampoix, H.; Mousain, D. Enzymatic hydrolysis of organic phosphorus. In Organic Phosphorus in the Environment; Turner, B.L., Frossard, E., Baldwin, D.S., Eds.; CABI: Wallingford, UK; pp. 89–112.
- Tarafdar, J.C.; Jungk, A. Phosphatase activity in the rhizosphere and its relation to the depletion of soil organic phosphorus. Biol. Fertil. Soils 1987, 3, 199–204. [Google Scholar] [CrossRef]
- Chen, C.R.; Condron, L.M.; Davis, M.R.; Sherlock, R.R. Phosphorus dynamics in the rhizosphere of perennial ryegrass (Lolium perenne L.) and radiate pine (Pinus radiate D. Don). Soil Biol. Biochem. 2002, 34, 487–499. [Google Scholar] [CrossRef]
- Belinque, H.; Pucheu, N.; Kerber, N.; Rubio, G. Utilization of organic phosphorus sources by oilseed rape, sunflower and soybean. J. Plant Nutr. Soil Sci. 2015, 178, 339–344. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerke, J. Phytate (Inositol Hexakisphosphate) in Soil and Phosphate Acquisition from Inositol Phosphates by Higher Plants. A Review. Plants 2015, 4, 253-266. https://doi.org/10.3390/plants4020253
Gerke J. Phytate (Inositol Hexakisphosphate) in Soil and Phosphate Acquisition from Inositol Phosphates by Higher Plants. A Review. Plants. 2015; 4(2):253-266. https://doi.org/10.3390/plants4020253
Chicago/Turabian StyleGerke, Jörg. 2015. "Phytate (Inositol Hexakisphosphate) in Soil and Phosphate Acquisition from Inositol Phosphates by Higher Plants. A Review" Plants 4, no. 2: 253-266. https://doi.org/10.3390/plants4020253
APA StyleGerke, J. (2015). Phytate (Inositol Hexakisphosphate) in Soil and Phosphate Acquisition from Inositol Phosphates by Higher Plants. A Review. Plants, 4(2), 253-266. https://doi.org/10.3390/plants4020253