Abstract
Solidago canadensis L. is an expansive perennial that forms persistent, species-poor plant communities. It often spreads in fallow areas, displacing native floristic ingredients. Its expansiveness is largely due to morphological features, but it can also be the effect of allelopathic interaction. The aim of the experiment was to investigate the effect of aqueous extracts (decoction, infusion, and maceration) from dry S. canadensis leaves on germination and early growth stages of Raphanus sativus L. var. radicula Pers., in three cultivars: ‘Rowa’, ‘Półdługa’, and ‘Krakowianka’. In comparison to the control, the percentage of germinated radish seeds of ‘Rowa’ cultivar was statistically lower on the infusion and macerate. Regardless of the cultivar, the smallest changes in germination were found in seeds watered with decoction, and the largest with macerate. Seedlings length was most inhibited on macerate substrates, and least with infusion. Regardless of the form of the extract, each of them negatively affected the initial growth of radish seedlings. A fresh mass of ‘Rowa’ seedlings was inhibited by all Canadian goldenrod extracts. In relation to the control, the ‘Krakowianka’ cultivar was the least sensitive to S. canadensis extracts. The total chlorophyll content was the lowest in the seedlings of the ‘Rowa’ and ‘Półdługa’ cultivars germinated on macerate, compared to the control and two others extracts. The percentage of electrolyte leakage depended on the type of extract used and the radish cultivar. The study showed that depending on the cultivar, the aqueous extracts from S. canadensis leaves decreasing of germination and early growth of R. sativus.
1. Introduction
The proliferation of invasive species, through competition, changes in resource availability, and disturbance to the ecosystem, is one of the greatest threats to biodiversity [1,2]. Due to the properties of easy adaptation to new habitat conditions, effective competition for space, water, nutrients and pollinating insects, they gradually displace autochthonous species [3,4,5,6,7,8]. A good example of this kind of competition is the Canadian goldenrod, a species easily populating new areas in various regions of the world [9].
Canadian goldenrod Solidago canadensis L. (Asteraceae) is a perennial herbaceous plant that originates in North America. Currently, it grows in Europe, Asia, Australia, and New Zealand. It is found in humid habitats, on meadows, roadsides, in ditches, on both shady and sunny areas [10]. It differs, from the similar S. gigantea Aiton, of hairiness of the stem up to the inflorescences and the same length of the tubular flowers in capitulas (pseudanthia) of the entire complex inflorescence, resembling a panicle (Figure 1). In Poland, S. canadensis was introduced for ornamental purposes and as a food source for bees. This species is currently found throughout Poland; only in the north-eastern part of its population is slightly smaller [9,11,12]. S. canadensis due to its high growth, ease of reproduction, shows strong competitive abilities and influences the distribution of native plant species [13,14,15].
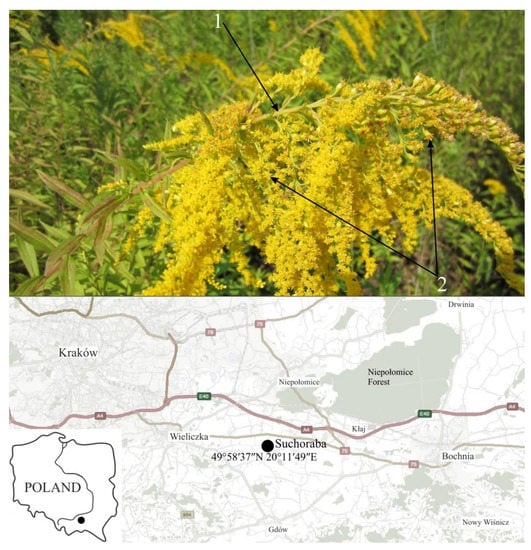
Figure 1.
General appearance of Solidago canadensis L.–arrows indicate diagnostic features: (1) hairy stem and (2) all the “lingule” flowers of equal length in composite inflorescences (Photo. J. Puła); place of collecting the plant material for experiment.
According to Huang and Guo [16], S. canadensis significantly reduces the overall number and diversity of species and causes severe losses in agriculture [17,18,19]. For example, it reduces germination and growth of a mulberry (Morus alba L.); morning glory (Pharbitis nil (L.) Roth), wheat (Triticum aestivum L.) and rape (Brassica campestris (L.) Koch [17]. Researchers have offered various hypotheses to explain the increased competitive ability of invasive plant species. These include phenotypic plasticity, release from natural enemies, evolution of increased competitive ability and the production of allelopathic compounds. The effects of S. canadensis invasion are less in communities with more numbers of native species [18,20].
Abhilasha et al. [10] proved that S. canadensis releases allelochemical compounds into the soil. This invasive alien weed causes severe shifts in the microorganism community in invaded areas, e.g., the enhancement (number of nitrite bacteria, aerobic Azotobacter, sulfate reducer, ammonifier and aerobic cellulose-decomposer), as well as reduction in the growth of denitrifier anaerobic cellulose-decomposer and desulphate reducer [21]. Sometimes the presence of specific allelochemicals derived from S. canadensis could be suggested in tomatoes cultivation due to their inhibitory effect on tomatoes’ soilborne pathogens (i.e., Pythium ultimum Trow. and Rhizoctonia solani J.G. Kühn) [22]. Although other authors pointed out that S. gigantea’s effect on soil microorganisms cannot last in subsequent vegetation periods and it depends on occurring plants (i.e., plant communities). During the three years mesocosm experiment changes in measured soil enzymes (i.e., β-glucosidase, β-N-acetylhexosaminidase and acid phosphatase, the enzymes mediating mineralization of C, N and P, respectively) were not clear. Effecta of increased β-glucosidase activity in rgw second year were no longer visible in the third vegetation period. S. gigantea did not affect β-N-acetylhexosaminidase activity, and acid phosphatase activity increased in Filipendulion plant community, but decreased in Molinion and Magnocaricion plant communities [23]. Moreover, authors, based on fungal biomass content in the invaded soil (accumulated during experiment), concluded that S. gigantea in their invasion strategy seem to follow toward pathogen-free areas as the plants fail to control their fungal pathogens in place.
S. canadensis’ main allelochemicals are flavones, phenolics, saponins, polyacetylates, essential oils [24,25], and other allelochemicals compounds, unknown so far [18,26]. Bearing in mind reports of allelopathic abilities of Canadian goldenrod, an attempt was made to assess this type of impact on seeds of Raphanus sativus L. var. radicula Pers. in three cultivars: ‘Rowa’, ‘Półdługa’, and ‘Krakowianka’. The research used radish seeds as a model organism. Radish seeds germinate in a very short time compared to other species. They have low nutritional and environmental requirements. As part of the research hypothesis, it was assumed that aqueous extracts of S. canadensis (in various forms: infusion, decoction and macerate) exhibit biological activity in relation to the studied radish cultivars, affecting: (1) its seed germination indexes, (2) length, fresh and dry mass of seedlings, (3) total chlorophyll content in seedlings and (4) percentage electrolytes leakage from seedling cells.
2. Results
2.1. Germination Indexes
On the first and second days of germination, the significantly highest germination speed (GS) of ‘Rowa’ and ‘Półdługa’ radish seeds was recorded in the control. On the third day, the lowest values of the index were recorded in all three radish cultivars on the macerate. In the following days, the GS was similar to seeds germinated on a distilled water and the decoction, regardless of the cultivar. In the case of the ‘Krakowianka’ cultivar, the highest GS for seeds watered with infusion was observed. For all radish cultivars treated with the macerate extract, a statistically significance decrease in GS values was found. On day 7, for the ‘Rowa’ and ‘Półdługa’ cultivars, the significantly highest percentage of germinated seeds was found for seeds treated with distilled water (control) and decoction. The macerate caused the complete disappearance of ‘Rowa’ cultivar germination. No radish sprouts were observed, but seeds were swollen, their seed coat was cracked in some cases. Significantly the lowest GS values for ’Półdługa’ and ’Krakowianka’ were found for seeds watered with macerate, compared to the control and the other two extracts (Table 1).

Table 1.
Germination speed (GS) (%) of Raphanus sativus L. cv. Rowa, Półdługa, Krakowianka, on the aqueous extracts from Solidago canadensis L. leaves and distilled water (control).
Compared to the control, the germination index (GI) values changed depending on the extract used. The largest changes were noted for macerate, in all radish cultivars. The decoction extract had the smallest but still statistically negative impact on the GI for ‘Rowa’ and ‘Krakowianka’. For the ‘Rowa’ and ‘Krakowianka’, no significance coefficient of the rate of germination (CRG) differences, between the control and decoction and infusion were observed. The CRG index for ‘Półdługa‘ was the highest for seeds watered with distilled water. For all radish cultivars germinated on macerate, the lowest values of this parameter were revealed. In the case of the T50 index, for ‘Rowa’ cultivar significant inhibiting influence of all S. canadensis aqueous extracts on the value of this parameter was demonstrated. For ‘Półdługa’, the infusion and macerate extracts decreased the T50, compared to the control. For ‘Krakowianka‘ a statistically significant differences were observed between macerate, and others extracts and distilled water. The seedling vigor index (SVI) index values for radish seedlings germinated on Petri dishes with aqueous extracts of S. canadensis were significantly lower, compared to the control. The smallest values of this parameter were shown for seedlings watered with macerate. The ‘Półdługa’ cultivar was an exception, for which no differences between the control and the macerate were found (Table 2).
Table 2.
Germination parameters of Raphanus sativus L. cv. Rowa, Półdługa, Krakowianka seeds germinated on the aqueous extracts from Solidago canadensis L. leaves and distilled water (control).
2.2. Length of Seedlings
The growth of radish seedlings treated with aqueous extracts of Canadian goldenrod leaves was inhibited by all extracts. In comparison to the control, regardless the cultivar, the macerate had the most negative affected of seedlings length of radish (Table 3). The inhibition percentage (IP) values were the lowest for seedlings watered with infusion. Among the radish cultivars analyzed, the elongation was inhibited most in the ‘Rowa’ and in the least in ‘Krakowianka’ cultivars (Table 3).
Table 3.
Length of Raphanus sativus L. cv. Rowa, Półdługa, Krakowianka seedlings, treatment of aqueous extracts from Solidago canadensis L. leaves.
2.3. Fresh, Dry Mass and Water Content
The fresh mass of radish seedlings was significantly larger in the control conditions for ‘Rowa‘ and ‘Półdługa‘. For ‘Rowa‘ cultivar, each of the type extracts decreased the fresh mass values. On the macerate extracts, ‘Rowa‘ seeds was swollen and had a crack seed coat, which significantly influenced the lowering of this parameter. The fresh mass of ‘Półdługa‘ seedlings was inhibited by decoction and macerate, compared to the control. For the ‘Krakowianka‘ no statistical differences in the values of this parameter were observed. Generally, the largest inhibition of fresh mass values were revealed for radish seedlings on macerate (Table 4).
Table 4.
Fresh and dry mass and water content of radish seeds Raphanus sativus L. cv. Rowa, Półdługa, Krakowianka, treatment of aqueous extracts from Solidago canadensis L. leaves.
The dry mass values of all radish cultivars germinated on S. canadensis extracts, did not differ significantly, compared to the control. The percentage of water content in ‘Rowa’ radish seedlings was significantly lower for seedlings watered with macerate. For the ‘Półdługa’ and ‘Krakowianka’ cultivars, no statistically significant changes were found in the values of this parameter (Table 4).
2.4. Chlorophyll Content and Electrolyte Leakage
The total chlorophyll content in seedlings of ‘Rowa‘ and ‘Krakowianka‘ was significantly lower in each treatment, compared to the control sample. The lowest concentration of chlorophyll was demonstrated for seedlings of the ‘Rowa’ and ‘Półdługa’ cultivars, watered with macerate. For the ‘Półdługa‘ cultivar, no differences in the chlorophyll content were found between the seedlings grown on the infusion and distilled water (Figure 2).
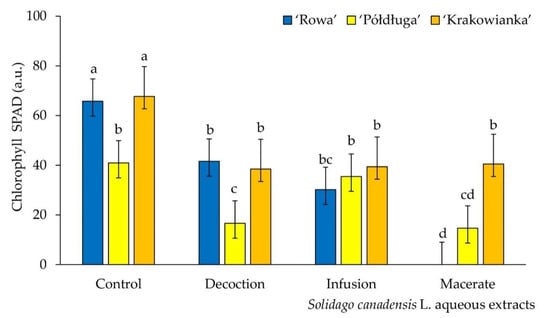
Figure 2.
Chlorophyll content of radish seeds (Raphanus sativus L. cv. Rowa, Półdługa, Krakowianka) treatment of aqueous extracts from Solidago canadensis L. leaves; mean values ± SD (n = 10) with different letters (a–d) differ statistically according to Duncan test at p < 0.05.
Percentage of electrolytes’ leakage in radish seedlings was the smallest in the control. Each of the aqueous extracts increased the degree of destabilization of the cell membranes of seedlings, regardless of the cultivar. The largest disturbances in water and ion management were found in seedlings of the ‘Rowa’ cultivar watered with macerate, in comparison to tested cultivars and extracts type. The ‘Rowa’ seeds on macerate, unlike the other varieties and extracts, were swollen, their seed coat was cracked in some case, hence most probably such a high% of electrolyte leakage. Compared to the control, for the ‘Rowa’ cultivar watered with decoction, no statistically significant differences were found. However, for the other two cultivars, significant differences were observed in the values of this parameter (Figure 3).
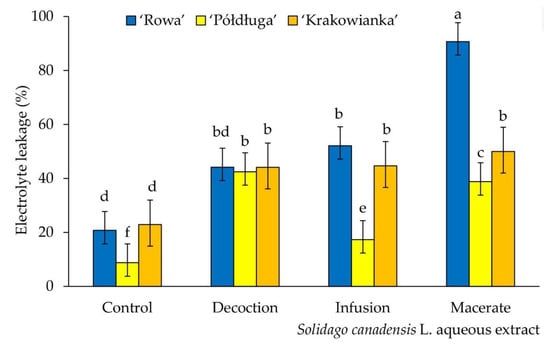
Figure 3.
Electrolyte leakage of three cultivars (‘Rowa‘, ‘Półdługa‘, ‘Krakowianka‘) of radish seeds (Raphanus sativus L.) treatment of aqueous extracts from Solidago canadensis L. leaves; mean values ± SD (n = 10) with different letter (a–f) differ statistically according to Duncan test at p < 0.05.
3. Discussion
In the conducted studies, aqueous extracts of Solidago canadensis L. leaves significantly slowed germination of seeds of the analyzed radish cultivars. Generally, germination indexes in each of the types of extracts were significantly lower, compared to the control (Table 1 and Table 2). The highest sensitivity to the effect of the extracts was demonstrated for the ‘Rowa’, and the lowest for ‘Krakowianka’ cultivar. Confirmation for the results obtained here, indicating a strong inhibitory effect of chemicals released from goldenrod leaves, are previous experimental studies [27,28,29,30]. Allelopathic potential of goldenrod extracts was found, among others in crops of carrot (Daucus sp.), barley (Hordeum sp.), maize (Zea mays L.) [31], lettuce (Lactuca sp.) [32] and coriander (Coriandrum sp.). The effect was observed even for crop’s weeds such as: velvetleaf (Abutilon theophrasti Med.) and redroot pigweed (Amaranthus retroflexus L.) [33].
The consequence of the negative impact of Canadian goldenrod on the germination of radish seeds was the inhibition of the elongation growth of seedlings and a decrease in the value of fresh mass. In the case of the dry mass, a statistically significant differences was not noted for tested radish cultivars, watered with three types of extracts (Table 3 and Table 4). Bingyao et al. [17] and Butcko and Jensen [34] observed the inhibitory and stimulating effect of S. canadensis extracts. The positive effect on mass increase may result from the stimulating effect of growth regulators responsible for cell growth, or from the activity of enzymes that improve metabolism and accelerate the flow of leaf assimilates to spare organs [35,36].
Aqueous extracts from goldenrod also decreased the chlorophyll content (Figure 2). Chlorophyll is one of the chemical compounds contained in plants that affects their photosynthetic activity and, consequently, the production of biomass. In chloroplasts, it is associated with specific proteins, glycolipids and sulfolipids. Its low content may be caused by inhibition of the synthesis of magnesium and iron chelate and protoporphyrinogen IX [37,38]. Observations of changes in chlorophyll content carried out here were significantly related to the type of extract used and the radish cultivar. In these measurements, the ‘Rowa’ cultivar was the most sensitive to the macerate extract. It can be assumed that the observed negative effects of using goldenrod extracts may be an effect of allelochemical compounds. Plants subjected to allelopathy stress vary in color intensity, which shows that they most likely contain less chlorophyll [39]. The chlorophyll content in the leaves is reduced, e.g., by phenolics [40]. The effect of allelochemical inhibitors depends on their concentration in the extract. The reduction of chlorophyll content may be caused by its degradation or may be a consequence of the restriction of its synthesis by allelochemical substances [37,41].
Each of the types of extracts used also caused an increase in the electrolyte leakage, which disrupted the water-ion balance of radish seedlings (Figure 3). Cell membranes are complex and dynamic structures made of lipids and proteins [42]. They are a measure of structural integrity, function stability and an indicator of plant tolerance to stress. Membranes are one of the first structures affected by stress factors. Their mechanism of functioning and physiological role indicate that electrolyte leakage is associated with the loss of K+ ions, mediating the conductivity of cations through the membrane [43]. In the conducted studies, it was found that the extract prepared in the form of macerate, irrespective of the radish cultivar, caused the largest inhibitory effect for the leakage of electrolytes. In the wide spectrum of allelochemical compounds, such properties are attributed to phenolic compounds [44]. These compounds are among the most common synthesized by plants and show a negative effect even at low concentrations. Allelochemical substances can penetrate into the cell membranes and destroy their structural connections or create new modification. For example, phenolic acids can change channels structure in cell membranes and limit the functioning of enzyme proteins, ATP, effect on transport and accumulation of ions between membranes and the water management of the cell. Other compounds, e.g., terpenoids, quinones, p-coumaric acid and ferulic acid cause lipid peroxidation. The cell membranes are often damaged by free radicals, which are formed, among others, by during the oxidative transformation of phenolics [44,45,46,47].
Differences in germination and early growth of radish seedlings in the presence of aqueous extracts from S. canadensis showed specificity of seed responses. They depended on both the radish cultivar and the type of extract used. Essentially, macerates had the most negative effects on germination and seed growth as well as chlorophyll content and electrolyte leakage. Extracts in the form of decoction and infusion also limited these processes, but to a much lesser extent than macerate. This kind of plant structure response suggests that aqueous extracts from Canadian goldenrod dry leaves in all forms have allelopathic properties. The leaves, from mature and flowering individuals used in the experiment, had a clearly inhibitory effect on the physiology of radish seeds. According to earlier studies on allelopathy [48], this may also suggest that leaves from young plants have even greater allelopathic potential. Therefore, the hypothesis on the biological properties of Canadian goldenrod can be here clearly confirmed. S. canadensis owes its expansion, as an invasive species, not only to clonal growth and interspecies relations [20], but also to the production of allelochemical compounds that significantly affect soil microflora and other plants [4,10,17,49].
The emission of allelochemical substances to the soil by species of the genus Solidago may pose a significant threat in the restoration of wasteland for agricultural production [50]. Therefore, one of the tasks in combating invasive weeds, such as goldenrod, is to clarify their adaptation to the environment, both physiologically and ecologically [51]. This will help to introduce effective and synthetic schemes for their elimination, forecast potential distribution areas and create a system for estimating threats to other species [52]. In Poland, no activities have been carried out to control and eliminate goldenrod so far. The only treatments used in the practice of controlling Solidago are regular mowing, point destruction of plants by pulling or digging, as well as afforestation, which increases shading and worsens habitat conditions for goldenrod [6,9]. Therefore, further research is needed to find tolerant and susceptible crops to goldenrod to fully assess its allelopathic properties.
4. Materials and Methods
4.1. Plant Material
Solidago canadensis L. leaves were collected during the flowering phase at the turn of July and August 2019, in Suchoraba (49°58′37″ N 20°11′49″ E) in southern Poland (Figure 1). The species occurred there massively, in the form of a compact yield, in a fallow field. The harvested fresh Canadian goldenrod leaf material was sorted such that pure plant material was selected for drying. Then the leaves were placed in a single layer on sheets of filter paper and dried in the dark, in an dryer at a constant temperature of 25 °C (Wamed SUP-100, Warsaw, Poland) for 7 days. Properly dried leaves had a natural color and crumbled easily. The dried plant material was stored in a silica gel desiccator.
Radish seeds were purchased at “Polan”–Breeding and Seed Horticulture in Krakow (Poland) company.
4.2. Extract Preparation
The dried Canadian goldenrod leaves were ground mechanically in a mill (Braun, 4045 type, Germany), and then aqueous extracts in the form of decoction, infusion and macerate were prepared, according to Tyszyńska-Kownacka and Starek [53]. The infusion was prepared from 5 g of dry and crushed S. canadensis leaves, which were flooded over with 250 mL of boiling distilled water and left covered for 30 min. After cooling, the extract was filtered through Whatman filter papers. The decoction was prepared from 8.75 g of dry plant material poured with a 1 liter of cold distilled water. The solution was mixed and left for 24 h in the dark, at 20–25 °C temperature. Then, the extract was boiled for 15 min and filtered. A macerated extract was prepared by flooding 5 g of dry leaves of 100 mL cold distilled water and left in the dark, at room temperature for 24 h. After one day, the extract was filtered the same form as infusion and decoction. The extracts were stored during the experiment at 8 °C.
4.3. Germination Conditions
Seeds of radish cultivars (Raphanus sativus L. cv. Rowa, Półdługa and Krakowianka) were purified in 5% sodium hypochlorite by 5 min and then washed three times with distilled water. The radish seeds (25 seeds per dish) were placed on sterile Petri dishes (ø 9 cm) with three layers of Whatman filter paper (Grade 1: 11 μm–medium flow filter paper). At the beginning of the experiment, the seeds were watered with the aqueous extracts (6 mL) in different form: decoctions, infusions, and macerate from S. canadensis leaves. A control group was seeds watered with distilled water. Every day seeds were watered with 3 mL of extracts and distilled water (control group). The Petri dishes with seeds were placed in a growth chamber (Angelantoni Industrie, Massa Martana, Italy) at 12 h/12 h (day/night) photoperiod, at a light intensity of 200 μmol × m–2 × s–1; the temperature was 25 °C/20 °C, and the relative humidity oscillated around 70–90%.
4.4. Germination Indexes
The germination capacity of seeds were checked every day (by 7 days). After this time, the following germination indexes were determined: a germination rate (GR), and germination speed (GS) [54], seedling vigor index (SVI) [55], coefficient of the rate of germination (CRG) [56], time required for 50% germination (T50) [57].
where: GT is the number of germinated seeds daily in the treatment, GC is the number of germinated seeds daily in the control, and D is the number of corresponding days.
where: n1 = number of germinated seeds on time T1; n2 = number of germinated seeds on time T2; n3 = number of germinated seeds on timeT3.
where: N is the final number of germination and ni, nj cumulative numbers of seeds germinated by adjacent counts at times ti and tj when ni < N/2 < nj.
GR = (number of germinated seeds/total number of seeds) × 100,
GS = ((GT × D)/(GC × D)) × 100,
SVI = (seedling length (cm) × percentage of germinated seeds)/100,
CRG = ((n1 + n2 + nn)/((n1 × T1) + (n2 × T2) + (n3 × T3) + …)) × 100,
T50 = ti + ((N/2) − nj) × (ti − tj))/(ni − nj),
4.5. Seedlings Length
Effects of aqueous extracts from S. canadensis leaves on seedlings growth (length of under-and above ground parts) were measured by a caliper (TOPEX 31C615, Poland), with an accuracy of 0,1 cm, after 7 days of experiment. An inhibition percentage expressed as % index (IP) was measured according to formula used by Islam and Kato-Noguchi [58].
where LE—seedling length (cm) treated with the aqueous extract, LC—seedling length (cm) treated with the distilled water (control group).
IP = (1 − (LE/LC)) × 100,
4.6. Fresh, Dry Mass and Water Content
After 7 days of germination the values of fresh and dry masses of whole seedlings (together under–and aboveground parts) were measured. Seedlings were weighted (fresh mass–FM) (Radwag WPS120, Radom, Poland) and dried (dry mass–DM) at 105 °C for 48 h in dryer (Wamed SUP 100, Warsaw, Poland). The tissue water content (% H2O) was determined according to the formula % H2O = 100 − ((DM × 100)/FM).
4.7. Chlorophyll Content and Electrolyte Leakage
The total chlorophyll content was measured by Chl SPAD by inserting the cotyledon into the clip of the SPAD chlorophyllometer device. It is not invasive method. The electrolyte leakage in radish seedlings was measured according to method used by Redmann et al. [59]. After 7 days of germination, single seedlings were placed in polypropylene vials with 30 mL distilled water and shaken for 3 h on a shaker (Labnet, Rocker, New Jersey, NJ, USA) to determine the electrolytes leakage from live cells (E1). The electrolytes leakage was measured by conductivity meter (type CX-701) with electrode (K = 0.92) (Elmetron, Zabrze, Poland). Then the plant material in vials was frozen at −70 °C. After 24 h the shaking procedure was repeated to the total electrolyte leakage from dead cells (E2) was measured. The percentage of electrolyte leakage (EL) was calculated according to the formula: EL = (E1/E2) 100%. The obtained percent results of electrolyte leakages from the control group of seedlings were compared to the values of seedlings germinated on the dishes with aqueous extracts.
4.8. Statistic Analysis
The experiment was carried out in three repetitions in two independent series. Significance of differences between objects (means ± SD; n = 10) was tested using the ANOVA multifactor analysis test, Duncan post hoc at p < 0.05 in Statistica 13.0 for Windows.
5. Conclusions
In the studies carried out here, the impact of aqueous extracts was uneven and clearly depended on the type of acceptor (radish cultivar) and donor (extract types).
(1) After seven days, the germination speed of seeds was the highest for cultivars watered with decoction and infusion, compared to the control. The macerate caused the complete disappearance of ‘Rowa’ germination. On the Solidago canadensis decoction radish seeds germinated better than on macerate extracts (most similar to the natural source of allelochemical compounds from dead organic debris). The values of germination indexes differed depending on the cultivar. The macerate extract showed a significant reduction in the value of germination indexes for all radish cultivars, in relation to the control. (2) The longest seedlings among all radish cultivars, grown on distilled water, were observed. The most inhibiting properties had a macerate. Regardless of the form of leaves extracts, each of them inhibited the elongation of seedlings of three radish cultivars, in comparison to the control group. The fresh mass of ‘Rowa’ seedlings were significant reduced by each of the S. canadensis extracts. The dry mass for all radish cultivars by S. canadensis extracts was not differ significantly, compared to the control values. (3) The all aqueous extracts from S. canadensis leaves, for ‘Rowa’ and ‘Krakowianka‘ cultivars, reduced the chlorophyll content. (4) Compared to the control, with the ‘Rowa’ cultivar the highest electrolyte leakage on macerate from S. canadensis leaves was revealed. All extracts caused an increase the electrolyte leakage in three cultivars of radish seedlings. The exception was the ‘Rowa’ cultivar treated to decoction, which no statistically significant differences were found in the destabilization of cell membranes.
In the context of these studies and hypothesis, it can be assumed that S. canadensis contributes to the delayed germination process of radish seeds, already in the early stages of vegetation.
Author Contributions
Conceptualization, K.M., B.B.-K.; methodology, K.M., B.B.-K. and J.P.; software, P.Z.; validation, K.M., B.B.-K. and J.P.; formal analysis, A.K., P.Z.; investigation, A.K., P.Z.; resources, K.M., B.B.-K.; data curation, K.M., B.B.-K. and J.P.; writing—original draft preparation, K.M., B.B.-K.; writing—review and editing, K.M., B.B.-K. and J.P.; visualization, A.K., J.P.; supervision, K.M., B.B.-K. and J.P.; project administration, K.M., B.B.-K. and J.P.; funding acquisition, J.P., A.K. All authors have read and agreed to the published version of the manuscript.
Funding
The research was financed by the Ministry of Science and Higher Education of the Republic of Poland.
Conflicts of Interest
The authors declare that there is no conflict of interest related to this article.
References
- McGeoch, M.A.; Butchart, S.H.M.; Spear, D.; Marais, E.; Kleynhans, E.J.; Symes, A.; Chanson, J.; Hoffmann, M. Global indicators of biological invasion: Species numbers, biodiversity impact and policy responses. Divers. Distrib. 2010, 16, 95–108. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Essl, F. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Crooks, J.A. Characterizing ecosystem-level consequences of biological invasions: The role of ecosystem engineers. Oikos 2002, 97, 153–166. [Google Scholar] [CrossRef]
- Jianzhong, L.; Wei, Q.; Jiakuan, C.; Bo, L. Impact of invasive species on soil properties: Canadian goldenrod (Solidago canadensis) as a case study. Chin. Biodivers. 2005, 13, 347–356. [Google Scholar] [CrossRef]
- Hulme, P.E.; Pyšek, P.; Nentwig, W.; Vilà, M. Will threat of biological invasions unite the European Union? Science 2009, 324, 40–41. [Google Scholar] [CrossRef]
- Tokarska-Guzik, B.; Dajdok, Z.; Urbisz, A.; Zając, M.; Danielewicz, W. Identification and categorisation of plants of foreign origin as the basis for practical activities. Acta Bot. Sil. 2011, 6, 23–53. [Google Scholar]
- Tang, L.; Gao, Y.; Wang, C.; Zhao, B.; Li, B. A plant invader declines through its modification to habitats: A case study of a 16-year chronosequence of Spartina alterniflora invasion in a salt marsh. Ecol. Eng. 2012, 49, 181–185. [Google Scholar] [CrossRef]
- Fenesi, A.; Vágási, C.I.; Beldean, M.; Földesi, R.; Kolcsár, L.P.; Shapiro, J.T.; Török, E.; Kovács-Hostyánszki, A. Solidago canadensis impacts on native plant and pollinator communities in different-aged old fields. Basic App. Ecol. 2015, 16, 335–346. [Google Scholar] [CrossRef]
- Tokarska-Guzik, B.E. The Establishment and Spread of Alien Plant Species (Kenophytes) in the Flora of Poland; Scientific Works no 2372; Silesian University Press: Katowice, Poland, 2005; Available online: http://www.sbc.org.pl/Content/39618/the_establishment_and_spread.pdf (accessed on 28 March 2020).
- Abhilasha, D.; Quintana, N.; Vivanco, J.; Joshi, J. Do allelopathic compounds in invasive Solidago canadensis restrain the native European flora. J. Ecol. 2008, 96, 993–1001. [Google Scholar] [CrossRef]
- Rola, J.; Rola, H. Solidago spp. as a bio-indicator of the occurrence of fallow lands on agricultural land. Frag. Agron. 2010, 27, 122–131. [Google Scholar]
- Aframowicz-Cieślak, R.; Bernacki, L.; Celka, Z.; Chmiel, J.; Cwener, A.; Dajdok, Z.; Danielewicz, W.; Kurnicki, B.; Łysko, A.; Marciniuk, J.; et al. Distribution atlas of vascular plants in Poland: Appendix. In Distribution Atlas of Vascular Plants in Poland; Zając, A., Zając, M., Eds.; Jagiellonian University Press: Warsaw, Poland, 2019; pp. 1–319. [Google Scholar]
- Pavek, P.L.S. Plant Guide for Canada Goldenrod (Solidago canadensis); USDA Natural Resources Conservation Service: Pullman, WA, USA, 2011.
- Chen, T.; Liu, W.L.; Zhang, C.B.; Wang, J. Effects of Solidago canadensis invasion on dynamics of native plant communities and their mechanisms. Chin. J. Plant Ecol. 2012, 36, 253–256. [Google Scholar] [CrossRef]
- Li, Y.Z.; Yin, X.; Wei, W.; Yang, E.Y.; Hang, Y.; Tian, Z.H.; Da, L.J. Inhibition of local plant Phragmites communis on the invasive plant Solidago canadensis. Acta Ecol. Sin. 2010, 30, 6881–6891. [Google Scholar]
- Huang, H.; Guo, S.L. Review on ecological studies on three invasive species of European genus Solidago. Guangxi Sci. 2004, 11, 69–74. [Google Scholar]
- Sun, B.Y.; Tan, J.Z.; Wan, Z.G.; Gu, F.G.; Zhu, M.D. Allelopathic effects of extracts from Solidago canadensis L. against seed germination and seedling growth of some plants. J. Environ. Sci. 2006, 18, 304–309. [Google Scholar]
- Yuan, Y.; Wang, B.; Zhang, S.; Tang, J.; Tu, C.; Hu, S.; Yong, J.W.H.; Xin, C. Enhanced allelopathy and competitive ability of invasive plant Solidago canadensis in its introduced range. Plant Ecol. 2013, 6, 253–263. [Google Scholar] [CrossRef]
- Ledger, K.J.; Pal, R.W.; Murphy, P.; Nagy, D.U.; Filep, R.; Callaway, R.M. Impact of an invader on species diversity is stronger in the non-native range than in the native range. Plant Ecol. 2015, 216, 1285–1295. [Google Scholar] [CrossRef]
- Powell, K.I.; Chase, J.M.; Knight, T.M. Invasive plants have scale-dependent effects on diversity by altering species-area relationships. Science 2013, 339, 316–318. [Google Scholar] [CrossRef]
- Zhang, F.; Wan, F. Canada goldenrod Solidago canadensis L. In Biological Invasions and Its Management in China; Wan, F., Jiang, M., Zhan, A., Eds.; Springer: Dordrecht, The Netherlands, 2017; Volume 13, pp. 143–151. [Google Scholar]
- Zhang, S.; Jin, Y.; Tang, J.; Chen, X. The invasive plant Solidago canadensis L. suppresses local soil pathogens through allelopathy. Appl. Soil Ecol. 2009, 41, 215–222. [Google Scholar] [CrossRef]
- Scharfy, D.; Güsewell, S.; Gessner, M.O.; Venterink, H.O. Invasion of Solidago gigantea in contrasting experimental plant communities: Effects on soil microbes, nutrients and plant–soil feedbacks. J. Ecol. 2010, 1379–1388. [Google Scholar] [CrossRef]
- Zhang, J.S.; Li, B.; Chen, J.K.; Zhou, T.S. Chemical constituents and antimicrobial activity of volatile oil from Solidago canadensis L. J. Found. Univ. Nat. Sci. 2006, 45, 412–415. [Google Scholar]
- Zhang, S.; Zhu, W.; Wang, B.; Tang, J.; Chen, X. Secondary metabolites from the invasive Solidago canadensis L. accumulation in soil and contribution to inhibition of soil pathogen Pythium ultimum. Appl. Soil Ecol. 2011, 48, 280–286. [Google Scholar] [CrossRef]
- Deng, Y.; Zhao, Y.; Padilla-Zakour, O.; Yang, G. Polyphenols, antioxidant and antimicrobial activities of leaf and bark extracts of Solidago canadensis L. Ind. Crop. Prod. 2015, 74, 803–809. [Google Scholar] [CrossRef]
- Fisher, R.F.; Woods, R.A.; Glavicic, M.R. Allelopathic effects of goldenrod and aster on young sugar maple. Can. J. For. Res. 1977, 8, 1–9. [Google Scholar] [CrossRef]
- Yang, R.Y.; Mei, L.X.; Tang, J.J.; Chen, X. Allelopathic effects of invasive Solidago canadensis L. on germination and growth of native Chinense plant species. Allelopath. J. 2007, 19, 241–248. [Google Scholar]
- Pisula, N.L.; Meiners, S.J. Allelopathic effects of goldenrod species on turnover in successional communities. Am. Midl. Nat. 2010, 163, 161–172. [Google Scholar] [CrossRef]
- Li, S.L.; Li, Z.H.; Wang, Y.F.; Ruan, X.; Pan, C.D.; Wang, Q. Preliminary study for the allelopathic effect of water extracts from Solidago canadensis leaves. Adv. Mat. Res. 2013, 699, 340–348. [Google Scholar] [CrossRef]
- Ye, X.Q.; Meng, J.L.; Wu, M. The effects of Solidago canadensis water extracts on maize seedling growth in association with the biomass allocation pattern. Peer J. 2019, 7, e6564. [Google Scholar] [CrossRef]
- Wang, C.; Jiang, K.; Wua, B.; Zhoua, J. The combined treatments of Canada goldenrod leaf extracts and cadmium pollution confer an inhibitory effect on seed germination and seedling development of lettuce. Aust. J. Bot. 2018, 66, 331–337. [Google Scholar] [CrossRef]
- Baličević, R.; Ravlić, M.; Živkovic, T. Allelopathic effect of invasive species giant goldenrod (Solidago gigantea Sit.) on craps and weeds. Herbologia 2015, 15, 19–29. [Google Scholar]
- Butcko, V.M.; Jensen, R.J. Evidence of tissue-specific allelopathic activity in Euthamia graminifolia and Solidago canadensis (Asteraceae). Am. Midl. Nat. 2002, 148, 253–262. [Google Scholar] [CrossRef]
- Petrášek, J.; Friml, J. Auxin transport routes in plant development. Development 2009, 136, 2675–2688. [Google Scholar] [CrossRef]
- Anžlovar, A.L.; Anžlovar, S. Allelopathic effect of aqueous extracts of Canadian goldenrod on germination and growth of radish. Acta Biol. Slov. 2019, 62, 27–34. [Google Scholar]
- Dżugan, M. Factors affecting the stability of green plant pigments. Soil Sci. Ann. 2006, 7, 27–33. [Google Scholar]
- Aarti, P.; Tanaka, R.; Tanaka, A. Effects of oxidative stress on chlorophyll biosynthesis in cucumber (Cucumis sativus) cotyledons. Physiol. Plant. 2006, 128, 186–197. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984; p. 422. [Google Scholar]
- Pandey, I.D.K. Inhibiton of najas (Najas graminea Del.) by parthenium (Parthenium hysterophorus L.). Allelopath. J. 1997, 4, 121–126. [Google Scholar]
- Baziramakenga, R.; Simard, R.R.; Leroux, G.D. Effects of benzoic and cinnamic acids on growth, mineral composition, and chlorophyll content of soybean. J. Chem. Ecol. 1994, 20, 2821–2833. [Google Scholar] [CrossRef] [PubMed]
- Yordanov, I.; Velikova, V.; Tsonev, T. Plant responses to drought, acclimation, and stress tolerance. Photosynthetica 2000, 38, 171–186. [Google Scholar] [CrossRef]
- Demidchik, V.; Straltsova, D.; Medvedev, S.S.; Pozhvanov, G.A.; Sokolik, A.; Yurin, V. Stress-induced electrolyte leakage: The role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp. Bot. 2014, 65, 1259–1270. [Google Scholar] [CrossRef]
- Einhelling, F.A. Mechanism of action of allelochemicals in allelopathy. In Allelopathy, Organisms, Processes and Applications; Inderjit, K.M.M., Dakshini, F.A., Einhelling, Eds.; ASC Symp., Ser. 582; American Chemical Society: Washington, DC, USA, 1995; pp. 96–116. [Google Scholar]
- Balke, N.E. Effects of allelochemicals on mineral uptake and associated physiological processes. In The Chemistry of Allelopathy; Thompson, A.C., Ed.; American Chemical Society: Washington, DC, USA, 1985; pp. 161–178. [Google Scholar] [CrossRef]
- Vaughan, D.; Ord, B.G. Extraction of potential allelochemicals and their effects on root morphology and nutrient contents. In Plant Growth and Ecological Perspective; Atkinson, A., Ed.; Blackwell Scientific: Oxford, UK, 1991; pp. 399–421. [Google Scholar]
- Walter, M.H. Regulation of lignification in defense. In Plant Gene Res. Genes Involved in Plant Defense; Boller, T., Meins, F., Eds.; Springer-Verlag: Wien, Austria, 1992; pp. 27–352. [Google Scholar] [CrossRef]
- Wójcik-Wojtkowiak, D.; Politycka, B.; Weyman-Kaczmarkowa, W. Allelopathy; University of Life Sciences in Poznań Press: Poznań, Poland, 1998; pp. 1–92. [Google Scholar]
- Dong, L.J.; Yu, H.W.; He, W.M. What determines positive, neutral, and negative impacts of Solidago canadensis invasion on native plant species richness? Sci. Rep. 2015, 5, 16804. [Google Scholar] [CrossRef]
- Đikić, M. Allelopathic effect of aromatic and medicinal plants on the seed germination of Galinsoga parviflora. Echinochloa crusgalli and Galium molugo. Herbologia 2005, 6, 51–57. [Google Scholar]
- Baležentienė, L. Allelopathic activity of invasive species Solidago canadensis L. In Proceedings of the 7th international Scientific Conference Rural Development 2015: Towards the Transfer of Knowledge, Innovations and Social Progress, Aleksandras Stulginskis University, Akademija, Lithuania, 19–20 November 2015; Raupelienė, A., Ed.; Aleksandras Stulginskis University Press: Kaunas, Lithuania, 2015. [Google Scholar]
- Guo, S.L.; Fang, F. Physiological adaptation of the invasive plant Solidago canadensis to environments. Acta Phytoecol. Sin. 2003, 27, 47–52. [Google Scholar] [CrossRef]
- Tyszyńska-Kownacka, D.; Starek, T. Herbs in Polish House; Wydawnictwo Warta: Warszawa, Poland, 1989; p. 223. [Google Scholar]
- AOSA (Association of Official Seed Analysis). Seed vigour testing. In Handbook on Seed Testing; Anmol Publications Pvt. Ltd.: New Delhi, India, 1983. [Google Scholar]
- Islam, A.K.M.A.; Anuar, N.; Yaakob, Z. Effect of genotypes and pre-sowing treatments on seed germination behavior of Jatropha. Asian J. Plant Sci. 2009, 8, 433–439. [Google Scholar] [CrossRef]
- Chiapusio, G.; Sanchez, A.M.; Reigosa, M.J.; Gonzalez, L.; Pellissier, F. Do germination indices adequately reflect allelochemical effects on the germination process? J. Chem. Ecol. 1997, 23, 2445–2453. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Afzal, I.; Khaliq, A. Optimization of hydropriming techniques for rice seed invigoration. Seed Sci. Technol. 2006, 34, 507–512. [Google Scholar] [CrossRef]
- Mominul Islam, A.K.M.; Kato-Noguchi, H. Allelopathic potentiality of medicinal plant Leucas aspera. Int. J. Agric. Sustain. 2012, 4, 1–7. [Google Scholar]
- Redmann, R.E.; Haraldson, J.; Gusta, L.V. Leakage of UV-absorbing substances as a measure of salt injury in leaf tissue of woody species. Acta Physiol. Plant. 1986, 67, 87–91. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).